
Preventive Effects of Cocoa and Cocoa Antioxidants in Colon Cancer



Abstract
:1. Introduction
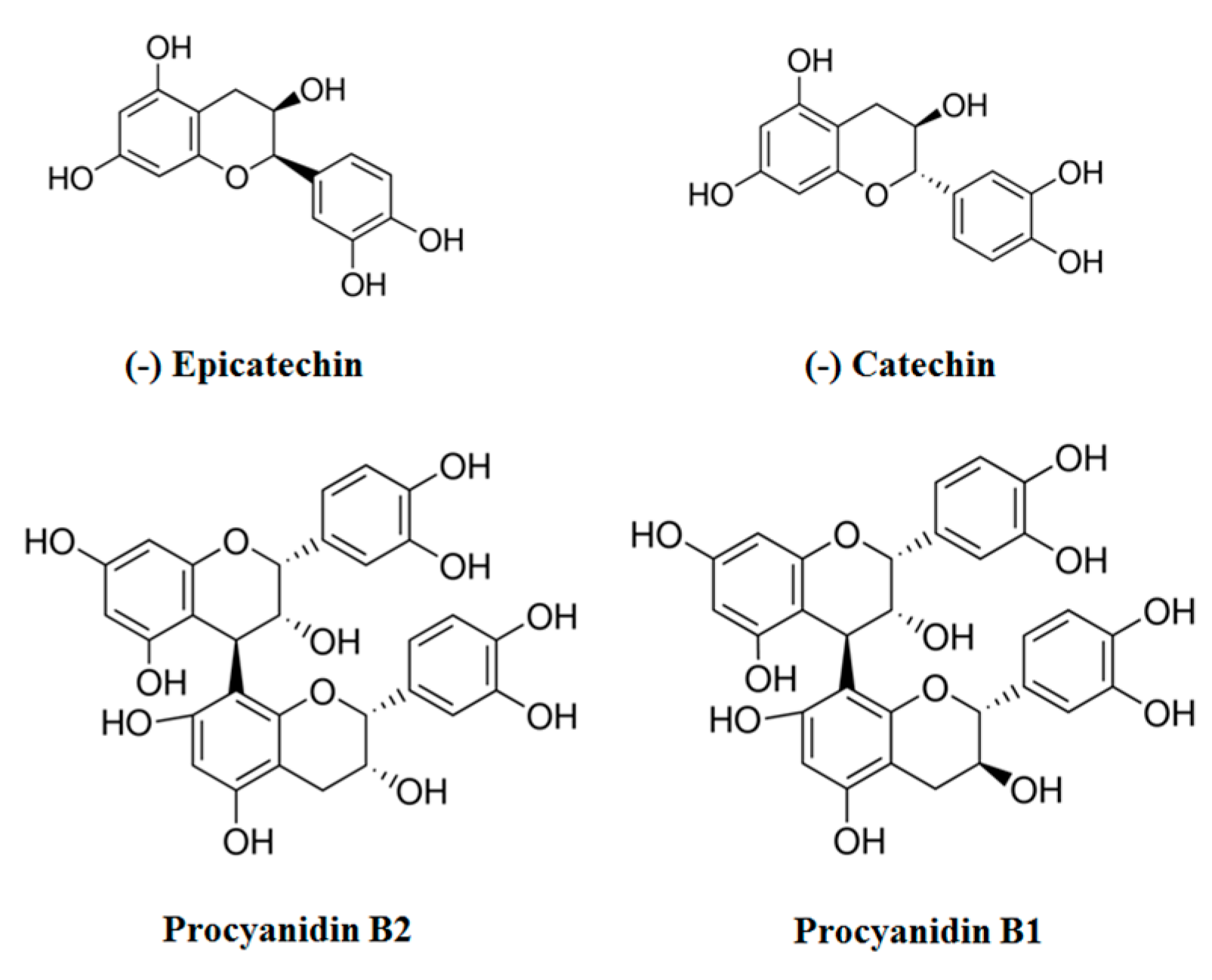
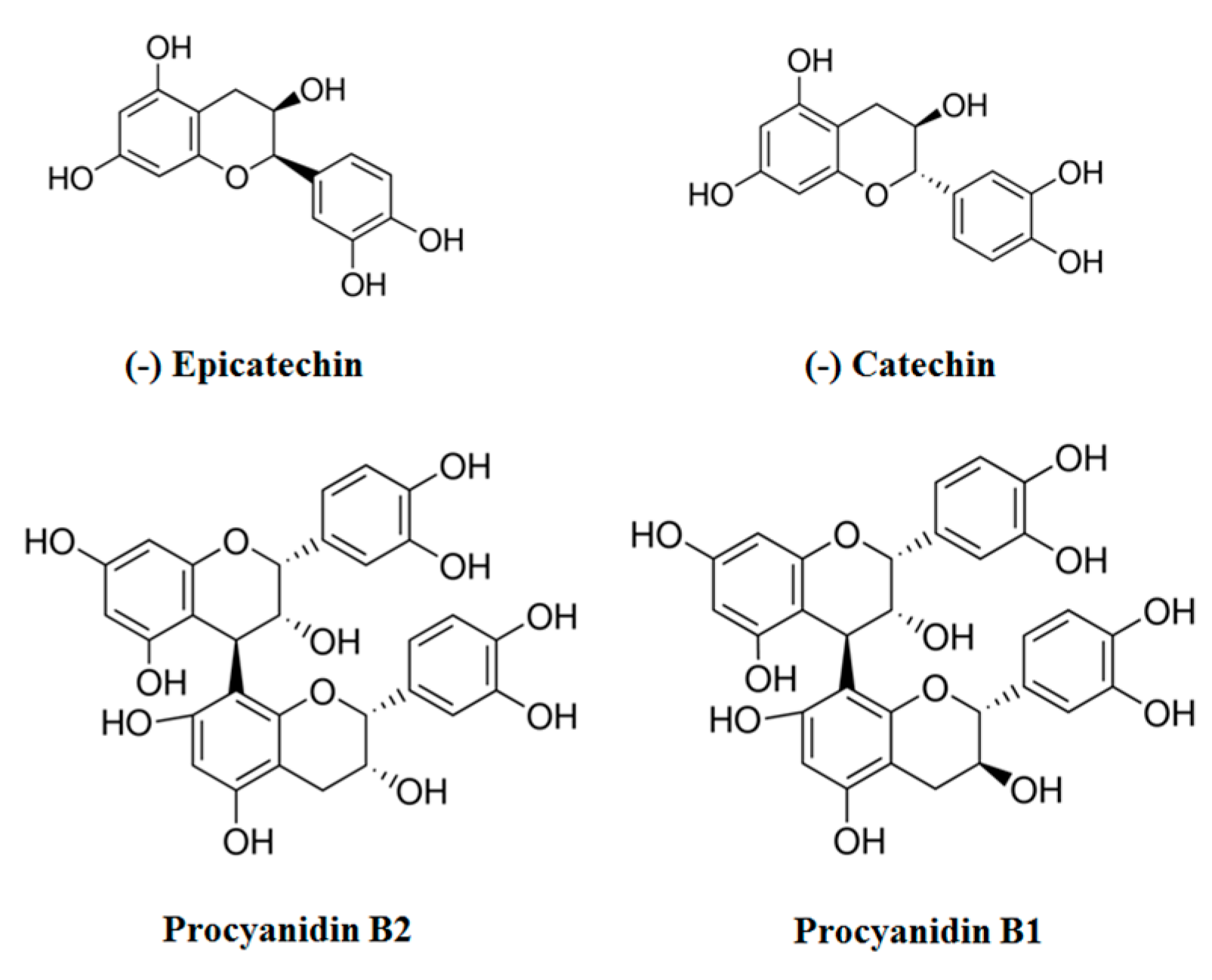
2. Chemopreventive Effects in Cultured Cells

{kind=link}
{kind=link}
| Biological Activity | Flavonoid | Cell | Concentration | Output | References |
|---|---|---|---|---|---|
| Antioxidant | Cocoa | Caco-2 | 10 µg/mL | acrylamide-incubated cells: ↓ GSH depletion, ↓ ROS generation, ↑ γ-GCS, ↑ GST | [20] |
| Antioxidant | Procyanidin B2 | Caco-2 | 10 µM (5.79 µg/mL) | acrylamide-incubated cells: ↓GSH depletion, ↓ ROS generation, ↑ γ-GCS, ↑ GST | [20] |
| Antioxidant | Epicatechin | Caco-2 | 10 µM (2.9 µg/mL) | acrylamide-incubated cells: ↓ GSH depletion, ↓ ROS generation | [20] |
| Apoptosis and proliferation/survival | Cocoa | Caco-2 | 10 µg/mL | ↓ acrylamide-induced caspase-3 and p-JNK | [20] |
| Apoptosis and proliferation/survival | Procyanidin B2 | Caco-2 | 10 µM (5.79 µg/mL) | ↑ ERK, ↑ p38, ↓ acrylamide-induced caspase-3 and p-JNK | [20] |
| Antioxidant | Catechin | Int-407 | 100 µM (29 µg/mL) | ↓ lipid peroxidation, ↓ ROS formation, ↑ GPx, ↑ GR, ↑ Nrf2, ↑ HO-1 | [21] |
| Antioxidant | Epicatechin | Caco-2 | 1–10 µM (0.29–2.9 µg/mL) | t-BOOH-treated cells: ↓ ROS generation, ↓ LDH, = GPx, = GST, = GR | [22] |
| Apoptosis and proliferation/survival | Procyanidin B2 | Caco-2 | 10 µM (5.79 µg/mL) | ↓ t-BOOH-induced caspase-3 | [22] |
| Apoptosis and proliferation/survival | Epicatechin | Caco-2 | 10 µM (2.9 µg/mL) | ↓ t-BOOH-induced caspase-3 | [22] |
| Antioxidant | Procyanidin B2 | Caco-2 | 1–10 µM (0.6–5.79 µg/mL) | ↑ GPx, ↑ GST, ↑ GR, ↑ Nrf2 translocation | [22,23] |
| Apoptosis and proliferation/survival | Hexamer procyanidins | Caco-2 | 2.5–20 µM (0.73–5.81 µg/mL) | ↓ DOC-induced caspase-3, ↓ PPAR cleavage | [24] |
| Apoptosis and proliferation/survival | Hexamer procyanidins | Caco-2 | 10 µM (2.91 µg/mL) | ↓ DOC-induced AKT, ERK, p38 and AP-1 | [24] |
| Antioxidant | Hexamer procyanidins | Caco-2 | 2.5–20 µM (0.73–5.81 µg/mL) | t-BOOH-treated cells: ↓ ROS generation, ↓ LDH ↓ DOC-induced cytotoxicity, ↓ oxidant generation, ↓ NADPH oxidase, ↓ Ca2+ | [24,25] |
| Anti-inflammatory | Hexamer procyanidins | Caco-2 | 2.5–60 µM (0.73–17.4 µg/mL) | TNF-treated cells: ↓ NF-κB activation (↓ p-IĸB, ↑ IĸB, ↓ p50 and p65 translocation, ↓ NF-κB-DNA binding), ↓ iNOS | [25] |
| Anti-inflammatory | Cocoa | Caco-2 | 50 µM (gallic acid equivalents, 14.5 µg/mL) | ↓ PGE2, ↑ COX-1, IL-stimulated cells: ↓ PGE2, = IL-8, = NF-κB | [26] |
| Anti-inflammatory | Cocoa | Caco-2 | 10 µg/mL | TNF-treated cells: ↓ IL-8, ↓ COX-2, ↓ iNOS, ↓ NFκB activation | [27] |
| Cell cycle | Polymer procyanidins | Caco-2 | 5–100 µg/mL | G2/M arrest, ↓ ornithine decarboxylase, ↓ S-adenosylmethionine decarboxylase | [28] |
| Cell cycle | Epicatechin | LoVo | 5–1000 µM (1.45–290 µg/mL) | S arrest | [29] |
| Apoptosis and proliferation/survival | Procyanidin B2 | SW480 | 10–50 µM (5.79–28.93 µg/mL) | ↑ proliferation, ↑ p-AKT, ↑ p-ERK | [30] |
| Apoptosis and proliferation/survival | Procyanidin B2 | Caco-2 | 10–50 µM (5.79–28.93 µg/mL) | = proliferation, = p-AKT, = p-ERK | [30] |
| Apoptosis and proliferation/survival | Epicatechin | SW480 and Caco-2 | 10–50 µM (2.9–14.5 µg/mL) | = proliferation, = p-AKT, = p-ERK | [30] |
2.1. Antioxidative Effects
2.2. Anti-Inflammatory Effects
2.3. Antiproliferative and Apoptotic Effects
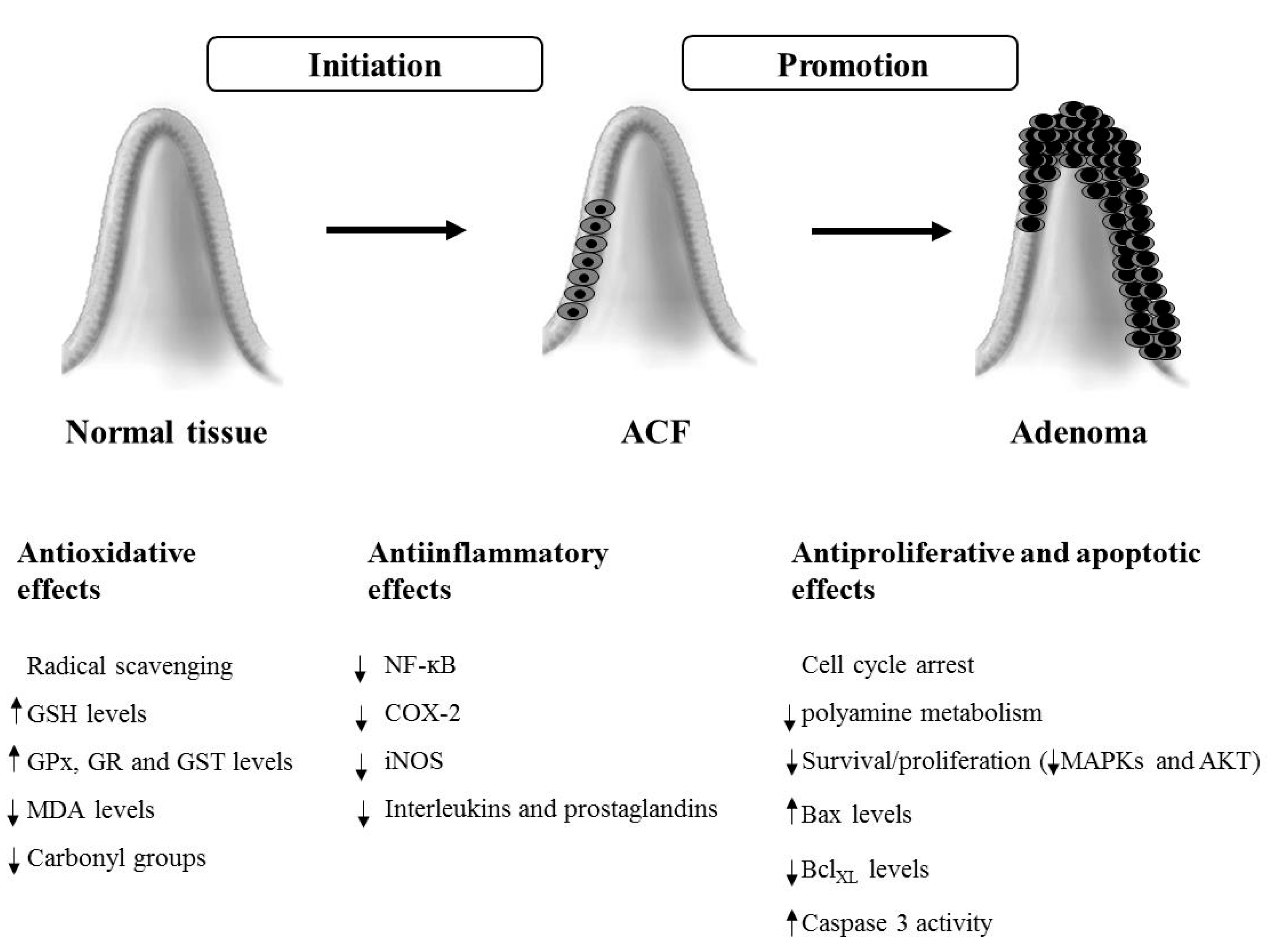
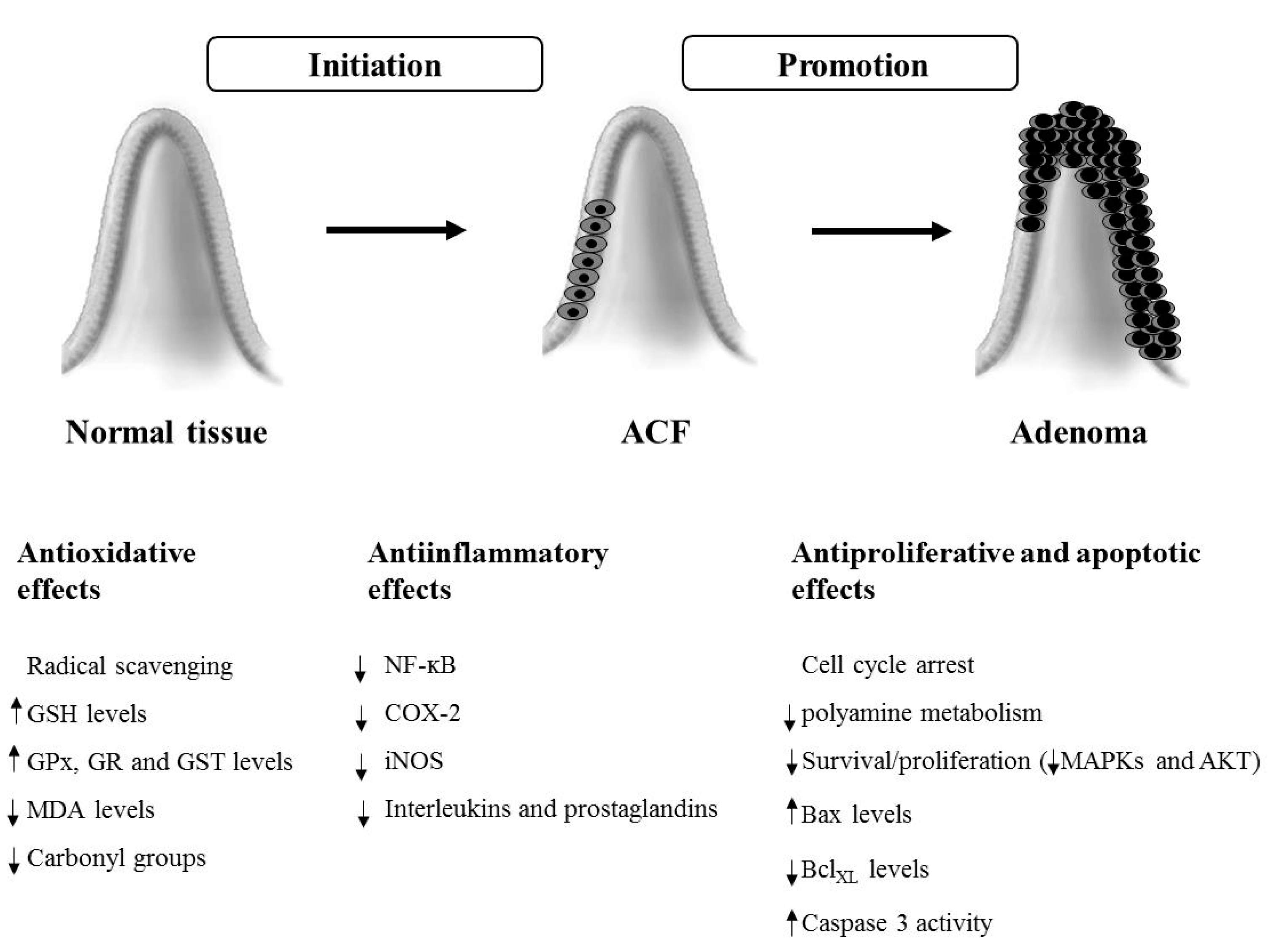
3. Chemopreventive Effects in Animal Models
4. Evidence of Chemopreventive Effects in Humans
4.1. Epidemiologic Studies
4.2. Intervention Studies
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Pritchard, C.C.; Grady, W.M. Colorectal cancer molecular biology moves into clinical practice. Gut 2011, 60, 116–129. [Google Scholar] [CrossRef] [PubMed]
- Reuter, S.; Gupta, S.C.; Chaturvedi, M.M.; Aggarwal, B.B. Oxidative stress, inflammation, and cancer: How are they linked? Free Radic. Biol. Med. 2010, 49, 1603–1616. [Google Scholar] [CrossRef] [PubMed]
- Aoi, W.; Naito, Y.; Takagi, T.; Kokura, S.; Mizushima, K.; Takanami, Y. Regular exercise reduces colon tumorigenesis associated with suppression of iNOS. Biochem. Biophys. Res. Commun. 2010, 399, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Chiou, Y.-S.; Tsai, M.-L.; Nagabhushanam, K.; Wang, Y.-J.; Wu, C.H.; Ho, C.T.; Pan, M.H. Pterostilbene is more potent than resveratrol in preventing azoxymethane (AOM)-induced colon tumorigenesis via activation of the NF-E2-related factor 2 (Nrf2)-mediated antioxidant signaling pathway. J. Agric. Food Chem. 2011, 59, 2725–2733. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Ng, J.; Arozulllah, A.; Ewing, R.; Llor, X.; Carroll, R.E.; Benya, R.V. Aberrant crypt focus size predicts distal polyp histopathology. Cancer Epidemiol. Biomark. Prev. 2008, 17, 1155–1162. [Google Scholar] [CrossRef] [PubMed]
- Pan, M.-H.; Lai, C.-S.; Wu, J.-C.; Ho, C.-T. Molecular mechanisms for chemoprevention of colorectal cancer by natural dietary compounds. Mol. Nutr. Food Res. 2011, 55, 32–45. [Google Scholar] [CrossRef] [PubMed]
- Ramos, S. Cancer chemoprevention and chemotherapy: Dietary polyphenols and signalling pathways. Mol. Nutr. Food Res. 2008, 52, 507–526. [Google Scholar] [CrossRef] [PubMed]
- Ramos, S. Effects of dietary flavonoids on apoptotic pathways related to cancer chemoprevention. J. Nutr. Biochem. 2007, 18, 427–442. [Google Scholar] [CrossRef] [PubMed]
- Martín, M.A.; Goya, L.; Ramos, R. Potential for preventive effects of cocoa and cocoa polyphenols in cancer. Food Chem. Toxicol. 2013, 56, 336–351. [Google Scholar] [CrossRef] [PubMed]
- Ramiro-Puig, E.; Castell, M. Cocoa: Antioxidant and immunomodulator. Br. J. Nutr. 2009, 101, 931–940. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.W.; Kim, Y.J.; Lee, H.J.; Lee, C.Y. Cocoa has more phenolic phytochemicals and a higher antioxidant capacity than teas and red wine. J. Agric. Food Chem. 2003, 51, 7292–7295. [Google Scholar] [CrossRef] [PubMed]
- Rusconi, M.; Conti, A. Theobroma cacao L., the Food of the Gods: A scientific approach beyond myths and claims. Pharmacol. Res. 2010, 61, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Vinson, J.A.; Proch, J.; Zubik, L. Phenol antioxidant quantity and quality in foods: Cocoa, dark chocolate, and milk chocolate. J. Agric. Food Chem. 2009, 47, 4821–4824. [Google Scholar] [CrossRef]
- Cooper, K.A.; Donovan, J.L.; Waterhouse, A.L.; Williamson, G. Cocoa and health: A decade of research. Br. J. Nutr. 2008, 99, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Manach, C.; Williamson, G.; Morand, C.; Scalbert, A.; Rémésy, C. Bioavailability and bioefficacy of polyphenols in humans. I. Review of 97 bioavailability studies. Am. J. Clin. Nutr. 2005, 81, 230S–242S. [Google Scholar] [PubMed]
- Sánchez-Rabaneda, F.; Jáuregui, O.; Casals, I.; Andrés-Lacueva, C.; Izquierdo-Pulido, M.; Lamuela-Raventós, R.M. Liquid chromatographic/electrospray ionization tandem mass spectrometric study of the phenolic composition of cocoa (Theobroma cacao). J. Mass Spectrom. 2003, 38, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Urpí-Sardá, M.; Monagas, M.; Khan, N.; Lamuela-Raventós, R.M.; Santos-Buelga, C.; Sacanella, E.; Castell, M.; Permanyer, J.; Andrés-Lacueva, C. Epicatechin, procyanidins, and phenolic microbial metabolites after cocoa intake in humans and rats. Anal. Bioanal. Chem. 2009, 394, 1545–1556. [Google Scholar] [CrossRef] [PubMed]
- Urpí-Sardá, M.; Ramiro-Puig, E.; Khan, N.; Ramos-Romero, S.; Llorach, R.; Castell, M.; González-Manzano, S.; Santos-Buelga, C.; Andrés-Lacueva, C. Distribution of epicatechin metabolites in lymphoid tissues and testes of young rats with a cocoa-enriched diet. Br. J. Nutr. 2010, 103, 1393–1397. [Google Scholar] [CrossRef] [PubMed]
- Monagas, M.; Urpí-Sardá, M.; Sánchez-Patán, F.; Llorach, R.; Garrido, I.; Gómez-Cordovés, C.; Andrés-Lacueva, C.; Bartolomé, B. Insights into the metabolism and microbial biotransformation of dietary flavan-3-ols and the bioactivity of their metabolites. Food Funct. 2010, 1, 233–253. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Ramiro, I.; Ramos, S.; Bravo, L.; Goya, L.; Martín, M.A. Procyanidin B2 and a cocoa polyphenolic extract inhibit acrylamide-induced apoptosis in human Caco-2 cells by preventing oxidative stress and activation of JNK pathway. J. Nutr. Biochem. 2011, 22, 1186–1194. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.T.; Wu, C.H.; Ho, C.Y.; Yen, G.C. Catechin protects against ketoprofen-induced oxidative damage of the gastric mucosa by up-regulating Nrf2 in vitro and in vivo. J. Nutr. Biochem. 2013, 24, 475–483. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Ramiro, I.; Martín, M.A.; Ramos, S.; Bravo, L.; Goya, L. Comparative effects of dietary flavanols on antioxidant defences and their response to oxidant-induced stress on Caco2 cells. Eur. J. Nutr. 2011, 50, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Ramiro, I.; Ramos, S.; Bravo, L.; Goya, L.; Martín, M.A. Procyanidin B2 induces Nrf2 translocation and glutathione S-transferase P1 expression via ERKs and p38-MAPK pathways and protect human colonic cells against oxidative stress. Eur. J. Nutr. 2012, 51, 881–892. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, M.; Jaggers, G.K.; Verstraeten, S.V.; Erlejman, A.G.; Fraga, C.G.; Oteiza, P.I. Large procyanidins prevent bile-acid-induced oxidant production and membrane-initiated ERK1/2, p38, and Akt activation in Caco-2 cells. Free Radic. Biol. Med. 2012, 52, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Erlejman, A.G.; Jaggers, G.; Fraga, C.G.; Oteiza, P.I. TNFα-induced NF-κB activation and cell oxidant production are modulated by hexameric procyanidins in Caco-2 cells. Arch. Biochem. Biophys. 2008, 476, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Romier-Crouzet, B.; van de Walle, J.; During, A.; Joly, A.; Rousseau, C.; Henry, O.; Larondelle, Y.; Schneider, Y.J. Inhibition of inflammatory mediators by polyphenolic plant extracts in human intestinal Caco-2 cells. Food Chem. Toxicol. 2009, 47, 1221–1230. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Ramiro, I.; Ramos, S.; López-Oliva, E.; Agís-Torres, A.; Bravo, L.; Goya, L.; Martín, M.A. Cocoa polyphenols prevent inflammation in the colon of 2 azoxymethane-treated rats and in TNF-a-stimulated Caco-2 cells. Br. J. Nutr. 2013, 110, 206–215. [Google Scholar] [CrossRef] [PubMed]
- Carnésecchi, S.; Schneider, Y.; Lazarus, S.A.; Coehlo, D.; Gossé, F.; Raul, F. Flavanols and procyanidins of cocoa and chocolate inhibit growth and polyamine biosynthesis of human colonic cancer cells. Cancer Lett. 2002, 175, 147–155. [Google Scholar] [CrossRef]
- Tan, X.; Hu, D.; Li, S.; Han, Y.; Zhang, Y.; Zhou, D. Differences of four catechins in cell cycle arrest and induction of apoptosis in LoVo cells. Cancer Lett. 2000, 158, 1–6. [Google Scholar] [CrossRef]
- Ramos, S.; Rodríguez-Ramiro, I.; Martín, M.A.; Goya, L.; Bravo, L. Dietary flavanols exert different effects on antioxidant defenses and apoptosis/proliferation in Caco-2 and SW480 colon cancer cells. Toxicol. Vitro 2011, 25, 1771–1781. [Google Scholar] [CrossRef] [PubMed]
- Finkel, T.; Holbrook, N. Oxidants, oxidative stress and the biology of ageing. Nature 2000, 408, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Verstraeten, S.V.; Hammerstone, J.F.; Keen, C.L.; Fraga, C.G.; Oteiza, P.I. Antioxidant and membrane effects of procyanidin dimers and trimers isolated from peanut and cocoa. J. Agric. Food Chem. 2005, 53, 5041–5048. [Google Scholar] [CrossRef] [PubMed]
- Pan, M.H.; Lai, C.S.; Ho, C.T. Anti-inflammatory activity of natural dietary flavonoids. Food Funct. 2010, 1, 15–31. [Google Scholar] [CrossRef] [PubMed]
- Bitzer, Z.T.; Glisan, S.L.; Dorenkott, M.R.; Goodrich, K.M.; Ye, L.; O’Keefe, S.F.; Lambert, J.D.; Neilson, A.P. Cocoa procyanidins with different degrees of polymerization possess distinct activities in models of colonic inflammation. J. Nutr. Biochem. 2015, 26, 827–831. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, D.W.; Giardina, C.; Tanaka, T. Mouse models for the study of colon carcinogenesis. Carcinogenesis 2009, 30, 183–196. [Google Scholar] [CrossRef] [PubMed]
- Reddy, B.S. Studies with the azoxymethane-rat preclinical model for assessing colon tumor development and chemoprevention. Environ. Mol. Mutagen. 2004, 44, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Raju, J. Azoxymethane-induced rat aberrant crypt foci: Relevance in studying chemoprevention of colon cancer. World J. Gastroenterol. 2008, 14, 6632–6635. [Google Scholar] [CrossRef] [PubMed]
- Weyant, M.J.; Carothers, A.M.; Dannenberg, A.J.; Bertagnoll, M.M. (+)-Catechin inhibits intestinal tumor formation and suppresses focal adhesion kinase activation in the Min/1 mouse. Cancer Res. 2001, 61, 118–125. [Google Scholar] [PubMed]
- Mahmoud, N.N.; Kucherlapati, R.; Bilinski, R.T.; Churchill, M.R.; Chadburn, A.; Bertagnolli, M.M. Genotype-phenotype correlation in murine Apc mutation: Differences in enterocyte migration and response to sulindac. Cancer Res. 1999, 59, 353–359. [Google Scholar] [PubMed]
- Reagan-Shaw, S.; Nihal, M.; Ahmad, N. Dose translation from animal to human studies revisited. FASEB J. 2007, 22, 659–661. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Ramiro, I.; Ramos, S.; López-Oliva, E.; Agís-Torres, A.; Gómez-Juaristi, M.; Mateos, R.; Bravo, L.; Goya, L.; Martín, M.A. Cocoa-rich diet prevents azoxymethane-induced colonic preneoplastic lesions in rats by restraining oxidative stress and cell proliferation and inducing apoptosis. Mol. Nutr. Food Res. 2011, 55, 1895–1899. [Google Scholar] [CrossRef] [PubMed]
- Hong, M.Y.; Nulton, E.; Shelechi, M.; Hernández, L.M.; Nemoseck, T. Effects of dark chocolate on azoxymethane-induced colonic aberrant crypt foci. Nutr. Cancer 2013, 65, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Pandurangan, A.K.; Saadatdoust, Z.; Esa, N.M.; Hamzah, H.; Ismail, A. Dietary cocoa protects against colitis-associated cancer by activating the Nrf2/Keap1 pathway. BioFactors 2015, 41, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Saadatdousta, Z.; Pandurangana, A.K.; Sadagopanc, S.K.A.; Esa, N.M.; Ismail, A.; Mustafa, M.R. Dietary cocoa inhibits colitis associated cancer: A crucial involvement of the IL-6/STAT3 pathway. J. Nutr. Biochem. 2015. [Google Scholar] [CrossRef] [PubMed]
- Bayard, V.; Chamorro, F.; Motta, J.; Hollenberg, N.K. Does flavanol intake influence mortality from nitric oxide-dependent processes? Ischemic heart disease, stroke, diabetes mellitus, and cancer in Panama. Int. J. Med. Sci. 2007, 4, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Arts, I.; Jacobs, D., Jr.; Gross, M.; Harnack, L.; Folsom, A. Dietary catechins and cancer incidence among postmenopausal women: The Iowa Women’s Health Study (United States). Cancer Causes Control 2002, 13, 373–382. [Google Scholar] [CrossRef] [PubMed]
- Rossi, M.; Negri, E.; Parpinel, M.; Lagiou, P.; Bosetti, C.; Talamini, R.; Montella, M.; Giacosa, A.; Franceschi, S.; la Vecchia, C. Proanthocyanidins and the risk of colorectal cancer in Italy. Cancer Causes Control 2010, 21, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Rouillier, P.; Senesse, P.; Cottet, V.; Valleau, A.; Faivre, J.; Boutron-Ruault, M.C. Dietary patterns and the adenoma-carcinoma sequence of colorectal cancer. Eur. J. Nutr. 2005, 44, 311–318. [Google Scholar] [CrossRef] [PubMed]
- McKelvey, W.; Greenland, S.; Sandler, R.S. A second look at the relation between colorectal adenomas and consumption of foods containing partially hydrogenated oils. Epidemiology 2000, 11, 469–473. [Google Scholar] [CrossRef] [PubMed]
- Boutron-Ruault, M.C.; Senesse, P.; Faivre, J.; Chatelain, N.; Belghiti, C.; Méance, S. Foods as risk factors for colorectal cancer: A case-control study in Burgundy (France). Eur. J. Cancer Prev. 1999, 8, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Maskarinec, G. Cancer protective properties of cocoa: A review of the epidemiologic evidence. Nutr. Cancer 2009, 61, 573–579. [Google Scholar] [CrossRef] [PubMed]
- Spadafranca, A.; Martinez Conesa, C.; Sirini, S.; Testolin, G. Effect of dark chocolate on plasma epicatechin levels, DNA resistance to oxidative stress and total antioxidant activity in healthy subjects. Br. J. Nutr. 2010, 103, 1008–1114. [Google Scholar] [CrossRef] [PubMed]
- Selmi, C.; Cocchi, C.A.; Lanfredini, M.; Keen, C.L.; Gershwin, M.E. Chocolate at heart: The anti-inflammatory impact of cocoa flavanols. Mol. Nutr. Food Res. 2008, 52, 1340–1348. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Agell, M.; Urpí-Sardá, M.; Sacanella, E.; Camino-López, S.; Chiva-Blanch, G.; Llorente-Cortés, V.; Tobias, E.; Roura, E.; Andrés-Lacueva, C.; Lamuela-Raventós, R.M.; et al. Cocoa consumption reduces NF-κB activation in peripheral blood mononuclear cells in humans. Nutr. Metab. Cardiovasc. Dis. 2013, 23, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Monagas, M.; Khan, N.; Andrés-Lacueva, C.; Casas, R.; Urpí-Sardá, M.; Llorach, R.; Lamuela-Raventós, R.M.; Estruch, R. Effect of cocoa powder on the modulation of inflammatory biomarkers in patients at high risk of cardiovascular disease. Am. J. Clin. Nutr. 2009, 90, 1144–1150. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martín, M.A.; Goya, L.; Ramos, S. Preventive Effects of Cocoa and Cocoa Antioxidants in Colon Cancer. Diseases 2016, 4, 6. https://doi.org/10.3390/diseases4010006
Martín MA, Goya L, Ramos S. Preventive Effects of Cocoa and Cocoa Antioxidants in Colon Cancer. Diseases. 2016; 4(1):6. https://doi.org/10.3390/diseases4010006
Chicago/Turabian StyleMartín, María Angeles, Luis Goya, and Sonia Ramos. 2016. "Preventive Effects of Cocoa and Cocoa Antioxidants in Colon Cancer" Diseases 4, no. 1: 6. https://doi.org/10.3390/diseases4010006
APA StyleMartín, M. A., Goya, L., & Ramos, S. (2016). Preventive Effects of Cocoa and Cocoa Antioxidants in Colon Cancer. Diseases, 4(1), 6. https://doi.org/10.3390/diseases4010006