
The Yin and Yang of Antibodies in Viral Infectious Diseases



Abstract
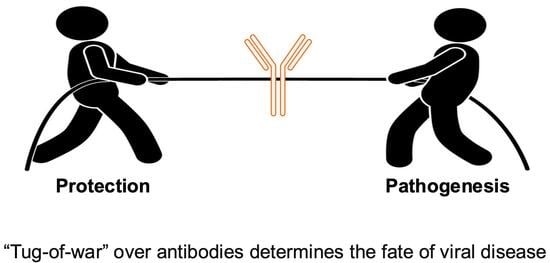
1. Introduction
2. Materials and Methods
3. Role of Antibodies in Viral Infections
3.1. Protective Role of Antibodies in Viral Infections
3.1.1. Neutralization
3.1.2. Tailoring the Response: Class-Switch Recombination and Fc-Mediated Effector Functions
3.1.3. Activation of the Complement System
3.1.4. Antibody-Mediated Phagocytosis
3.1.5. Antibody-Dependent Cellular Cytotoxicity (ADCC)
3.2. Detrimental and Pathological Roles of Antibodies in Viral Infections
3.2.1. Autoimmunity and Autoantibodies

{kind=link}
{kind=link}
| Autoimmune Disease/Condition | Associated Virus(es) | Autoantibodies/Autoantigen Targets | Proposed Immunopathogenic Mechanism(s) |
|---|---|---|---|
| Systemic Lupus Erythematosus (SLE) | Epstein–Barr Virus (EBV) [62] | Anti-nuclear antibodies (ANA), anti-dsDNA, anti-Sm, anti-CL/beta2-GPI complex, anti-RNP, anti-Ro/SSA, anti-La/SSB, antiphospholipid antibodies (aPL) [63] | FcγR dysregulation [64] |
| Rheumatoid Arthritis (RA) | Epstein–Barr Virus (EBV) [65], Cytomegalovirus (CMV) [66] | Rheumatoid Factor (RF) [42], anti-citrullinated protein antibodies (ACPAs) [67] | Bystander activation [68] |
| ANCA-Associated Vasculitis | Epstein–Barr Virus (EBV) [69], Hepatitis B Virus (HBV) [70] | Anti-neutrophil cytoplasmic antibodies (ANCAs) [43] | Neutrophil activation [71] |
| Multiple Sclerosis (MS) | Epstein–Barr Virus (EBV) [72] | Oligoclonal IgG bands (CSF); no single disease-defining serum autoantibody [73] | Molecular mimicry; Epitope spreading [74] |
| Guillain-Barré Syndrome (GBS) | Epstein–Barr Virus (EBV) [75], Cytomegalovirus (CMV) [76], Influenza [77], Zika Virus (ZIKV) [52] | Anti-ganglioside antibodies [52] | Molecular mimicry [51] |
| Multisystem Inflammatory Syndrome in Children (MIS-C) | SARS-CoV-2 [50] | Antibodies targeting viral nucleocapsid cross-reactive with self-protein SNX-8 [50] | Molecular mimicry [50] |
| Thrombocytopenia/Plasma Leakage | Dengue Virus (DENV) [37] | NS1 mimicry, endothelial and platelet autoimmunity [78] | Molecular mimicry [78] |
| Sjögren’s Syndrome | Epstein–Barr Virus (EBV) [79], Hepatitis C Virus (HCV) [56] | Anti-Ro/SSA (Ro52, Ro60), Anti-La/SSB, RF, ANA [80] | Epitope spreading [81] |
3.2.2. Therapeutic Strategies to Mitigate Antibody-Mediated Pathologies
3.2.3. Antibody-Dependent Enhancement (ADE)
4. Non-Neutralizing Antibodies (NNAbs)
5. Factors Influencing Antibody-Mediated Immunity
5.1. Viral-Based Factors
5.2. Host-Based Factors
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ADCC | Antibody-Dependent Cellular Cytotoxicity |
| ADCP | Antibody-Dependent Cellular Phagocytosis |
| ADE | Antibody-Dependent Enhancement |
| ANO2 | Anoctamin-2 |
| aPTT | Activated Partial Thromboplastin Time |
| BGLF5 | EBV Ribonuclease Protein |
| CCR6 | C-C chemokine receptor type 6 |
| CD107 | Lysosomal-Associated Membrane Protein 1 |
| CD16a | FcγRIIIa |
| CD16b | FcγRIIIb |
| CD32a | FcγRIIa |
| CD32b | FcγRIIb |
| CD32c | FcγRIIc |
| CD64 | FcγRI |
| CMV | Cytomegalovirus |
| CRP | C-Reactive Protein |
| CRYAB | Alpha-B Crystallin |
| CSR | Class Switching Recombination |
| CXCR3 | C-X-C Chemokine Receptor Type 3 |
| DENV | Dengue Virus |
| dsDNA | Double-Stranded DNA |
| EBNA1 | Epstein–Barr Nuclear Antigen 1 |
| EBV | Epstein–Barr Virus |
| FcγR | Fc Gamma Receptor |
| FcγRI | Fc Gamma Receptor I |
| FcγRIIa | Fc Gamma Receptor IIa |
| FcγRIIb | Fc Gamma Receptor IIb |
| FcγRIIc | Fc Gamma Receptor IIc |
| FcγRIIIa | Fc Gamma Receptor IIIa |
| FcγRIIIb | Fc Gamma Receptor IIIb |
| FcRs | Fc Receptors |
| GBS | Guillain-Barré Syndrome |
| GlialCAM | Glial Cell Adhesion Molecule |
| gp120 | Glycoprotein 120 |
| HA | Hemagglutinin |
| HA2 | Hemagglutinin Subunit 2 |
| HBV | Hepatitis B Virus |
| HCV | Hepatitis C Virus |
| HIV | Human Immunodeficiency Virus |
| HLA | Human Leukocyte Antigen |
| HSV | Herpes Simplex Virus |
| IFN-γ | Interferon-Gamma |
| IgA | Immunoglobulin A |
| IgD | Immunoglobulin D |
| IgE | Immunoglobulin E |
| IgG | Immunoglobulin G |
| IgM | Immunoglobulin M |
| IL-6 | Interleukin-6 |
| ITAM | Immunoreceptor Tyrosine-Based Activation Motif |
| ITIM | Immunoreceptor Tyrosine-Based Inhibitory Motif |
| JEV | Japanese Encephalitis Virus |
| LMP-2 | Latent membrane protein 2 |
| MBP | Myelin Basic Protein |
| MBCs | Memory B Cells |
| MIS-C | Multisystem Inflammatory Syndrome in Children |
| MS | Multiple Sclerosis |
| NNAbs | Non-Neutralizing Antibodies |
| NK | Natural Killer |
| NS1 | Non-structural protein 1 |
| PD-1 | Programmed Cell Death Protein 1 |
| PT | Prothrombin Time |
| RA | Rheumatoid Arthritis |
| RSV | Respiratory Syncytial Virus |
| SARS | Severe Acute Respiratory Syndrome |
| SARS-CoV-2 | Severe Acute Respiratory Syndrome Coronavirus 2 |
| SHM | Somatic Hypermutation |
| SLE | Systemic Lupus Erythematosus |
| SNX8 | Sorting Nexin 8 |
| STAT1 | Signal Transducer and Activator of Transcription 1 |
| Tfh | T follicular helper cells |
| TGF-β | Transforming growth factor-beta |
| TIGIT | T Cell Immunoreceptor with Ig and ITIM Domains |
| TNF-α | Tumor necrosis factor-alpha |
| WNV | West Nile Virus |
| ZIKV | Zika Virus |
References
- Bonilla, F.A.; Oettgen, H.C. Adaptive immunity. J. Allergy Clin. Immunol. 2010, 125 (Suppl. S2), S33–S40. [Google Scholar] [CrossRef]
- Sharma, S.K.; Cells, B. Basics of Hematopoietic Stem Cell Transplant; Springer: Berlin/Heidelberg, Germany, 2023; pp. 87–120. [Google Scholar]
- Stavnezer, J.; Schrader, C.E. IgH chain class switch recombination: Mechanism and regulation. J. Immunol. 2014, 193, 5370–5378. [Google Scholar] [CrossRef]
- Nothelfer, K.; Sansonetti, P.J.; Phalipon, A. Pathogen manipulation of B cells: The best defence is a good offence. Nat. Rev. Microbiol. 2015, 13, 173–184. [Google Scholar] [CrossRef]
- Doria-Rose, N.A.; Joyce, M.G. Strategies to guide the antibody affinity maturation process. Curr. Opin. Virol. 2015, 11, 137–147. [Google Scholar] [CrossRef]
- Pollard, A.J.; Bijker, E.M. A guide to vaccinology: From basic principles to new developments. Nat. Rev. Immunol. 2021, 21, 83–100. [Google Scholar] [CrossRef]
- Dempsey, L.A. Pathogenic antibodies. Nat. Immunol. 2019, 20, 1414. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.M.; Hwang, Y.C.; Liu, I.J.; Lee, C.C.; Tsai, H.Z.; Li, H.J.; Wu, H.C. Development of therapeutic antibodies for the treatment of diseases. J. Biomed. Sci. 2020, 27, 1. [Google Scholar] [CrossRef]
- Slifka, M.K.; Amanna, I.J. Role of Multivalency and Antigenic Threshold in Generating Protective Antibody Responses. Front. Immunol. 2019, 10, 956. [Google Scholar] [CrossRef] [PubMed]
- Klasse, P.J.; Sattentau, Q.J. Occupancy and mechanism in antibody-mediated neutralization of animal viruses. J. Gen. Virol. 2002, 83, 2091–2108. [Google Scholar] [CrossRef]
- Pantaleo, G.; Correia, B.; Fenwick, C.; Joo, V.S.; Perez, L. Antibodies to combat viral infections: Development strategies and progress. Nat. Rev. Drug. Discov. 2022, 21, 676–696. [Google Scholar] [CrossRef] [PubMed]
- Burton, D.R. Antiviral neutralizing antibodies: From In Vitro to In Vivo activity. Nat. Rev. Immunol. 2023, 23, 720–734. [Google Scholar] [CrossRef]
- Klasse, P.J. Neutralization of Virus Infectivity by Antibodies: Old Problems in New Perspectives. Advances. Biol. 2014, 2014, 157895. [Google Scholar] [CrossRef]
- Boero, E.; Gorham, R.D., Jr.; Francis, E.A.; Brand, J.; Teng, L.H.; Doorduijn, D.J.; Ruyken, M.; Muts, R.M.; Lehmann, C.; Verschoor, A.; et al. Purified complement C3b triggers phagocytosis and activation of human neutrophils via complement receptor 1. Sci. Rep. 2023, 13, 274. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Liu, L.; Qiang, H.; Sun, H.; Jiang, Y.; Ren, L.; Jiang, Z.; Lei, S.; Chen, L.; Wang, Y.; et al. A potent broad-spectrum neutralizing antibody targeting a conserved region of the prefusion RSV F protein. Nat. Commun. 2024, 15, 10085. [Google Scholar] [CrossRef] [PubMed]
- Okagawa, T.; Konnai, S.; Nishimori, A.; Maekawa, N.; Ikebuchi, R.; Goto, S.; Nakajima, C.; Kohara, J.; Ogasawara, S.; Kato, Y.; et al. Anti-Bovine Programmed Death-1 Rat-Bovine Chimeric Antibody for Immunotherapy of Bovine Leukemia Virus Infection in Cattle. Front. Immunol. 2017, 8, 650. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.R.; Lim, H.W.; Kang, S.G.; Hillsamer, P.; Kim, C.H. Human CD57+ germinal center-T cells are the major helpers for GC-B cells and induce class switch recombination. BMC Immunol. 2005, 6, 3. [Google Scholar] [CrossRef]
- Ding, Y.; Li, Y.; Wu, Z.; Zhou, Y.; Guo, Y.; Tian, S.; Yu, R.; Deng, C.; Wei, R.; Chen, H.; et al. A novel nanoparticle vaccine displaying multistage tuberculosis antigens confers protection in mice infected with H37Rv. NPJ Vaccines 2025, 10, 173. [Google Scholar] [CrossRef]
- Yu, H.; Lin, L.; Zhang, Z.; Zhang, H.; Hu, H. Targeting NF-κB pathway for the therapy of diseases: Mechanism and clinical study. Signal Transduct. Target. Ther. 2020, 5, 209. [Google Scholar] [CrossRef]
- van Erp, E.A.; Lakerveld, A.J.; de Graaf, E.; Larsen, M.D.; Schepp, R.M.; Ederveen, A.L.H.; Ahout, I.M.; de Haan, C.A.; Wuhrer, M.; Luytjes, W.; et al. Natural killer cell activation by respiratory syncytial virus-specific antibodies is decreased in infants with severe respiratory infections and correlates with Fc-glycosylation. Clin. Transl. Immunol. 2020, 9, e1112. [Google Scholar] [CrossRef]
- Annamalai, B.; Nicholson, C.; Parsons, N.; Stephenson, S.; Atkinson, C.; Jones, B.; Rohrer, B. Immunization Against Oxidized Elastin Exacerbates Structural and Functional Damage in Mouse Model of Smoke-Induced Ocular Injury. Investig. Ophthalmol. Vis. Sci. 2020, 61, 45. [Google Scholar] [CrossRef]
- Lewis, M.J.; Wagner, B.; Irvine, R.M.; Woof, J.M. IgA in the horse: Cloning of equine polymeric Ig receptor and J chain and characterization of recombinant forms of equine IgA. Mucosal Immunol. 2010, 3, 610–621. [Google Scholar] [CrossRef]
- Thomas, S.; Smatti, M.K.; Ouhtit, A.; Cyprian, F.S.; Almaslamani, M.A.; Al Thani, A.; Yassine, H.M. Antibody-Dependent Enhancement (ADE) and the role of complement system in disease pathogenesis. Mol. Immunol. 2022, 152, 172–182. [Google Scholar] [CrossRef]
- Byrne, A.B.; Talarico, L.B. Role of the complement system in antibody-dependent enhancement of flavivirus infections. Int. J. Infect. Dis. 2021, 103, 404–411. [Google Scholar] [CrossRef]
- Castro-Dopico, T.; Clatworthy, M.R. IgG and Fcγ Receptors in Intestinal Immunity and Inflammation. Front. Immunol. 2019, 10, 805. [Google Scholar] [CrossRef]
- Lamptey, H.; Bonney, E.Y.; Adu, B.; Kyei, G.B. Are Fc Gamma Receptor Polymorphisms Important in HIV-1 Infection Outcomes and Latent Reservoir Size? Front. Immunol. 2021, 12, 656894. [Google Scholar] [CrossRef]
- Seeling, M.; Pöhnl, M.; Kara, S.; Horstmann, N.; Riemer, C.; Wöhner, M.; Liang, C.; Brückner, C.; Eiring, P.; Werner, A.; et al. Immunoglobulin G-dependent inhibition of inflammatory bone remodeling requires pattern recognition receptor Dectin-1. Immunity 2023, 56, 1046–1063.e7. [Google Scholar] [CrossRef] [PubMed]
- Vattepu, R.; Sneed, S.L.; Anthony, R.M. Sialylation as an Important Regulator of Antibody Function. Front. Immunol. 2022, 13, 818736. [Google Scholar] [CrossRef] [PubMed]
- Irvine, E.B.; Alter, G. Understanding the role of antibody glycosylation through the lens of severe viral and bacterial diseases. Glycobiology 2020, 30, 241–253. [Google Scholar] [CrossRef]
- Lembo, A.; Molinaro, A.; De Castro, C.; Berti, F.; Biagini, M. Impact of glycosylation on viral vaccines. Carbohydr. Polym. 2024, 342, 122402. [Google Scholar] [CrossRef]
- Gómez Román, V.R.; Murray, J.C.; Weiner, L.M. Chapter 1—Antibody-Dependent Cellular Cytotoxicity (ADCC). In Antibody Fc; Ackerman, M.E., Nimmerjahn, F., Eds.; Academic Press: Boston, MA, USA, 2014; pp. 1–27. [Google Scholar]
- Niu, Y.-X.; Xu, Z.-X.; Yu, L.-F.; Lu, Y.-P.; Wang, Y.; Wu, C.; Hou, Y.-B.; Li, J.-N.; Huang, S.; Song, X.; et al. Advances of research of Fc-fusion protein that activate NK cells for tumor immunotherapy. Int. Immunopharmacol. 2022, 109, 108783. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, U.; Chakrabarti, A.K.; Kanungo, S.; Dutta, S. Evolutionary dynamics of influenza A/H1N1 virus circulating in India from 2011 to 2021. Infect. Genet. Evol. 2023, 110, 105424. [Google Scholar] [CrossRef]
- He, W.; Tan, G.S.; Mullarkey, C.E.; Lee, A.J.; Lam, M.M.W.; Krammer, F.; Henry, C.; Wilson, P.C.; Ashkar, A.A.; Palese, P.; et al. Epitope specificity plays a critical role in regulating antibody-dependent cell-mediated cytotoxicity against influenza A virus. Proc. Natl. Acad. Sci. USA 2016, 113, 11931–11936. [Google Scholar] [CrossRef]
- Muralidharan, A.; Gravel, C.; Harris, G.; Hashem, A.M.; Zhang, W.; Safronetz, D.; Van Domselaar, G.; Krammer, F.; Sauve, S.; Rosu-Myles, M.; et al. Universal antibody targeting the highly conserved fusion peptide provides cross-protection in mice. Hum. Vaccin Immunother. 2022, 18, 2083428. [Google Scholar] [CrossRef]
- de Taeye, S.W.; Schriek, A.I.; Umotoy, J.C.; Grobben, M.; Burger, J.A.; Sanders, R.W.; Vidarsson, G.; Wuhrer, M.; Falck, D.; Kootstra, N.A.; et al. Afucosylated broadly neutralizing antibodies enhance clearance of HIV-1 infected cells through cell-mediated killing. Commun. Biol. 2024, 7, 964. [Google Scholar] [CrossRef]
- Wang, T.T.; Sewatanon, J.; Memoli, M.J.; Wrammert, J.; Bournazos, S.; Bhaumik, S.K.; Pinsky, B.A.; Chokephaibulkit, K.; Onlamoon, N.; Pattanapanyasat, K.; et al. IgG antibodies to dengue enhanced for FcγRIIIA binding determine disease severity. Science 2017, 355, 395–398. [Google Scholar] [CrossRef]
- Gudelj, I.; Lauc, G.; Pezer, M. Immunoglobulin G glycosylation in aging and diseases. Cell. Immunol. 2018, 333, 65–79. [Google Scholar] [CrossRef] [PubMed]
- Nimmerjahn, F. Role of Antibody Glycosylation in Health, Disease, and Therapy. In Handbook of Experimental Pharmacology; Anmol Publications: Delhi, India, 2025. [Google Scholar]
- Sundaresan, B.; Shirafkan, F.; Ripperger, K.; Rattay, K. The Role of Viral Infections in the Onset of Autoimmune Diseases. Viruses 2023, 15, 782. [Google Scholar] [CrossRef]
- Hargraves, M.M. Discovery of the LE cell and its morphology. Mayo Clin. Proc. 1969, 44, 579–599. [Google Scholar] [PubMed]
- Waaler, E. On the occurrence of a factor in human serum activating the specific agglutintion of sheep blood corpuscles 1939. APMIS 2007, 115, 422–438, discussion 439. [Google Scholar] [PubMed]
- Davies, D.J.; Moran, J.E.; Niall, J.F.; Ryan, G.B. Segmental necrotising glomerulonephritis with antineutrophil antibody: Possible arbovirus aetiology? Br. Med. J. (Clin. Res. Ed.) 1982, 285, 606. [Google Scholar] [CrossRef]
- Bastard, P.; Rosen, L.B.; Zhang, Q.; Michailidis, E.; Hoffmann, H.-H.; Zhang, Y.; Dorgham, K.; Philippot, Q.; Rosain, J.; Béziat, V.; et al. Autoantibodies against type I IFNs in patients with life-threatening COVID-19. Science 2020, 370, eabd4585. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.-K.; Yeo, K.-J.; Chang, S.-H.; Liao, T.-L.; Chou, C.-H.; Lan, J.-L.; Chang, C.-K.; Chen, D.-Y. The detectable anti-interferon-γ autoantibodies in COVID-19 patients may be associated with disease severity. Virol. J. 2023, 20, 33. [Google Scholar] [CrossRef]
- Lanz, T.V.; Brewer, R.C.; Ho, P.P.; Moon, J.-S.; Jude, K.M.; Fernandez, D.; Fernandes, R.A.; Gomez, A.M.; Nadj, G.-S.; Bartley, C.M.; et al. Clonally expanded B cells in multiple sclerosis bind EBV EBNA1 and GlialCAM. Nature 2022, 603, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Tengvall, K.; Huang, J.; Hellström, C.; Kammer, P.; Biström, M.; Ayoglu, B.; Bomfim, I.L.; Stridh, P.; Butt, J.; Brenner, N.; et al. Molecular mimicry between Anoctamin 2 and Epstein-Barr virus nuclear antigen 1 associates with multiple sclerosis risk. Proc. Natl. Acad. Sci. USA 2019, 116, 16955–16960. [Google Scholar] [CrossRef] [PubMed]
- Thomas, O.G.; Bronge, M.; Tengvall, K.; Akpinar, B.; Nilsson, O.B.; Holmgren, E.; Hessa, T.; Gafvelin, G.; Khademi, M.; Alfredsson, L.; et al. Cross-reactive EBNA1 immunity targets alpha-crystallin B and is associated with multiple sclerosis. Sci. Adv. 2023, 9, eadg3032. [Google Scholar] [CrossRef]
- Jog, N.R.; McClain, M.T.; Heinlen, L.D.; Gross, T.; Towner, R.; Guthridge, J.M.; Axtell, R.C.; Pardo, G.; Harley, J.B.; James, J.A. Epstein Barr virus nuclear antigen 1 (EBNA-1) peptides recognized by adult multiple sclerosis patient sera induce neurologic symptoms in a murine model. J. Autoimmun. 2020, 106, 102332. [Google Scholar] [CrossRef]
- Bodansky, A.; Mettelman, R.C.; Sabatino, J.J.; Vazquez, S.E.; Chou, J.; Novak, T.; Moffitt, K.L.; Miller, H.S.; Kung, A.F.; Rackaityte, E.; et al. Molecular mimicry in multisystem inflammatory syndrome in children. Nature 2024, 632, 622–629. [Google Scholar] [CrossRef]
- Zhou, F.; Xu, Y.; Liu, X.; Xu, Y.; Wang, Y.; Jiang, D.; Du, P. Zika and Dengue Virus Autoimmunity: An Overview of Related Disorders and Their Potential Mechanisms. Rev. Med. Virol. 2025, 35, e70014. [Google Scholar] [CrossRef]
- Cao-Lormeau, V.-M.; Blake, A.; Mons, S.; Lastère, S.; Roche, C.; Vanhomwegen, J.; Dub, T.; Baudouin, L.; Teissier, A.; Larre, P.; et al. Guillain-Barré Syndrome outbreak associated with Zika virus infection in French Polynesia: A case-control study. Lancet 2016, 387, 1531–1539. [Google Scholar] [CrossRef]
- Pisetsky, D.S. Pathogenesis of autoimmune disease. Nat. Rev. Nephrol. 2023, 19, 509–524. [Google Scholar] [CrossRef]
- Jiang, W.; Johnson, D.; Adekunle, R.; Heather, H.; Xu, W.; Cong, X.; Wu, X.; Fan, H.; Andersson, L.; Robertson, J.; et al. COVID-19 is associated with bystander polyclonal autoreactive B cell activation as reflected by a broad autoantibody production, but none is linked to disease severity. J. Med. Virol. 2022, 95, e28134. [Google Scholar] [CrossRef]
- Gregoire, C.; Spinelli, L.; Villazala-Merino, S.; Gil, L.; Holgado, M.P.; Moussa, M.; Dong, C.; Zarubica, A.; Fallet, M.; Navarro, J.-M.; et al. Viral infection engenders bona fide and bystander subsets of lung-resident memory B cells through a permissive mechanism. Immunity 2022, 55, 1216–1233.e9. [Google Scholar] [CrossRef]
- Priora, M.; Borrelli, R.; Parisi, S.; Ditto, M.C.; Realmuto, C.; Laganà, A.; Di Vittorio, C.C.; Degiovanni, R.; Peroni, C.L.; Fusaro, E. Autoantibodies and rheumatologic manifestations in hepatitis C virus infection. Biology 2021, 10, 1071. [Google Scholar] [CrossRef]
- Sattarnezhad, N.; Kockum, I.; Thomas, O.G.; Liu, Y.; Ho, P.P.; Barrett, A.K.; Comanescu, A.I.; Wijeratne, T.U.; Utz, P.J.; Alfredsson, L.; et al. Antibody reactivity against EBNA1 and GlialCAM differentiates multiple sclerosis patients from healthy controls. Proc. Natl. Acad. Sci. USA 2025, 122, e2424986122. [Google Scholar] [CrossRef] [PubMed]
- Sundar, K.; Jacques, S.; Gottlieb, P.; Villars, R.; Benito, M.-E.; Taylor, D.K.; Spatz, L.A. Expression of the Epstein-Barr virus nuclear antigen-1 (EBNA-1) in the mouse can elicit the production of anti-dsDNA and anti-Sm antibodies. J. Autoimmun. 2004, 23, 127–140. [Google Scholar] [CrossRef]
- Pagan, J.D.; Kitaoka, M.; Anthony, R.M. Engineered Sialylation of Pathogenic Antibodies In Vivo Attenuates Autoimmune Disease. Cell 2018, 172, 564–577.e13. [Google Scholar] [CrossRef]
- Sthoeger, Z.; Zinger, H.; Sharabi, A.; Asher, I.; Mozes, E. The tolerogenic peptide, hCDR1, down-regulates the expression of interferon-α in murine and human systemic lupus erythematosus. PLoS ONE 2013, 8, e60394. [Google Scholar] [CrossRef]
- Gazitt, T.; Eviatar, T.; Shear, J.; Meidan, R.; Furer, V.; Feld, J.; Haddad, A.; Elias, M.; Hijazi, N.; Stein, N.; et al. Development of Autoantibodies Following BNT162b2 mRNA COVID-19 Vaccination and Their Association with Disease Flares in Adult Patients with Autoimmune Inflammatory Rheumatic Diseases (AIIRD) and the General Population: Results of 1-Year Prospective Follow-Up Study. Vaccines 2023, 11, 476. [Google Scholar] [PubMed]
- Ulff-Møller, C.J.; Nielsen, N.M.; Rostgaard, K.; Hjalgrim, H.; Frisch, M. Epstein-Barr virus-associated infectious mononucleosis and risk of systemic lupus erythematosus. Rheumatology 2010, 49, 1706–1712. [Google Scholar] [CrossRef] [PubMed]
- Hansen, N.; Lipp, M.; Vogelgsang, J.; Vukovich, R.; Zindler, T.; Luedecke, D.; Gingele, S.; Malchow, B.; Frieling, H.; Kühn, S.; et al. Autoantibody-associated psychiatric symptoms and syndromes in adults: A narrative review and proposed diagnostic approach. Brain Behav. Immun. Health 2020, 9, 100154. [Google Scholar] [CrossRef]
- Sepúlveda-Delgado, J.; Llorente, L.; Hernández-Doño, S. A Comprehensive Review of Fc Gamma Receptors and Their Role in Systemic Lupus Erythematosus. Int. J. Mol. Sci. 2025, 26, 1851. [Google Scholar] [CrossRef]
- van de Rijn, M.; Cleary, M.L.; Variakojis, D.; Warnke, R.A.; Chang, P.P.; Kamel, O.W. Epstein-Barr virus clonality in lymphomas occurring in patients with rheumatoid arthritis. Arthritis Rheum 1996, 39, 638–642. [Google Scholar] [CrossRef]
- Almanzar, G.; Schmalzing, M.; Trippen, R.; Höfner, K.; Weißbrich, B.; Geissinger, E.; Meyer, T.; Liese, J.; Tony, H.-P.; Prelog, M. Significant IFNγ responses of CD8+ T cells in CMV-seropositive individuals with autoimmune arthritis. J. Clin. Virol. 2016, 77, 77–84. [Google Scholar] [CrossRef]
- Dong, X.; Zheng, Z.; Lin, P.; Fu, X.; Li, F.; Jiang, J.; Zhu, P. ACPAs promote IL-1β production in rheumatoid arthritis by activating the NLRP3 inflammasome. Cell. Mol. Immunol. 2019, 17, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Yosri, M.; Dokhan, M.; Aboagye, E.; Al Moussawy, M.; Abdelsamed, H.A. Mechanisms governing bystander activation of T cells. Front. Immunol. 2024, 15, 1465889. [Google Scholar] [CrossRef] [PubMed]
- Noonan, T.P.; Konstantinov, K.N.; Echevarria, L. Epstein-Barr virus reactivation induced myeloperoxidase-specific antineutrophil cytoplasmic antibody (MPO-ANCA)-associated vasculitis. BMJ Case. Rep. 2021, 14, e245059. [Google Scholar] [CrossRef]
- Nemoto, M.; Nishioka, K.; Fukuoka, J.; Aoshima, M. Hepatitis B Virus-associated Vasculitis: Multiple Cavitary Masses in the Lung Mimicking Granulomatous Polyangiitis. Intern. Med. 2019, 58, 3013–3017. [Google Scholar] [CrossRef]
- von Vietinghoff, S.; Choi, M.; Rolle, S.; Luft, F.C.; Kettritz, R. Febrile temperatures control antineutrophil cytoplasmic autoantibody-induced neutrophil activation via inhibition of phosphatidylinositol 3-kinase/Akt. Arthritis Rheum. 2007, 56, 3149–3158. [Google Scholar] [CrossRef]
- van Langelaar, J.; Wierenga-Wolf, A.F.; Samijn, J.P.A.; Luijks, C.J.; Siepman, T.A.; van Doorn, P.A.; Bell, A.; van Zelm, M.C.; Smolders, J.; van Luijn, M.M. The association of Epstein-Barr virus infection with CXCR3(+) B-cell development in multiple sclerosis: Impact of immunotherapies. Eur. J. Immunol. 2021, 51, 626–633. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Abrigo, J.; Chen, Q.; Au, C.; Ng, A.; Fan, P.; Mok, V.; Qiu, W.; Kermode, A.G.; Lau, A.Y. Advanced MRI features in relapsing multiple sclerosis patients with and without CSF oligoclonal IgG bands. Sci. Rep. 2020, 10, 13703. [Google Scholar] [CrossRef]
- Croxford, J.L.; Olson, J.K.; Miller, S.D. Epitope spreading and molecular mimicry as triggers of autoimmunity in the Theiler’s virus-induced demyelinating disease model of multiple sclerosis. Autoimmun. Rev. 2002, 1, 251–260. [Google Scholar] [CrossRef]
- Takahashi, K.; Kunishige, M.; Shinohara, M.; Kubo, K.; Inoue, H.; Yoshino, H.; Asano, A.; Honda, S.; Matsumoto, T.; Mitsui, T. Guillain-Barré syndrome and hemophagocytic lymphohistiocytosis in a patient with severe chronic active Epstein–Barr virus infection syndrome. Clin. Neurol. Neurosurg. 2005, 108, 80–83. [Google Scholar] [CrossRef]
- Ma, Y.; Feng, J.; Qi, Y.; Dou, X.G. An immunocompetent adult patient with hepatitis and guillain-barré syndrome after cytomegalovirus infection. Virol. J. 2011, 8, 95. [Google Scholar] [CrossRef]
- Mosora, O.; Barcutean, L.; Balasa, R.; Fodor, R.; Maier, S.; Bajko, Z.; Stoian, A.; Motataianu, A. Severe Acute Motor Axonal Neuropathy Associated with Influenza-A (H1N1) Infection and Prolonged Respiratory Failure—A Case Report. J. Crit. Care Med. 2021, 7, 302–307. [Google Scholar] [CrossRef] [PubMed]
- Ghorai, T.; Sarkar, A.; Roy, A.; Bhowmick, B.; Nayak, D.; Das, S. Role of auto-antibodies in the mechanisms of dengue pathogenesis and its progression: A comprehensive review. Arch. Microbiol. 2024, 206, 214. [Google Scholar] [CrossRef]
- Gorodetskiy, V.R.; Probatova, N.A.; Konovalov, D.M.; Ryzhikova, N.V.; Sidorova, Y.V.; Sudarikov, A.B.; Mukhortova, O.V. Composite Epstein-Barr virus-positive mucosa-associated lymphoid tissue lymphoma and Epstein-Barr virus-negative diffuse large B-cell lymphoma in the parotid salivary gland of a patient with Sjögren’s syndrome and rheumatoid arthritis: A case report. J. Med. Case Rep. 2020, 14, 12. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Gallo, L.; Oke, V.; Lundström, E.; Elvin, K.; Wu, Y.L.; Eketjäll, S.; Zickert, A.; Gustafsson, J.T.; Jönsen, A.; Leonard, D.; et al. Four Systemic Lupus Erythematosus Subgroups, Defined by Autoantibodies Status, Differ Regarding HLA-DRB1 Genotype Associations and Immunological and Clinical Manifestations. ACR Open Rheumatol. 2022, 4, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.Y.S.; Lin, M.W. Serological intermolecular epitope spreading in a patient with primary Sjögren’s syndrome. BMJ Case Rep. 2023, 16, e254632. [Google Scholar] [CrossRef]
- Wang, T.; Desai, A.A.; Thurber, G.M.; Tessier, P.M. Maximizing activity and selectivity of antibody-mediated effector functions using antibody mixtures. MAbs 2025, 17, 2480666. [Google Scholar] [CrossRef]
- Li, D.; Lou, Y.; Zhang, Y.; Liu, S.; Li, J.; Tao, J. Sialylated immunoglobulin G: A promising diagnostic and therapeutic strategy for autoimmune diseases. Theranostics 2021, 11, 5430–5446. [Google Scholar] [CrossRef]
- Lerner, A.; Benzvi, C.; Vojdani, A. SARS-CoV-2 Gut-Targeted Epitopes: Sequence Similarity and Cross-Reactivity Join Together for Molecular Mimicry. Biomedicines 2023, 11, 1937. [Google Scholar] [CrossRef]
- Randall, K.L. Rituximab in autoimmune diseases. Aust. Prescr. 2016, 39, 131–134. [Google Scholar] [CrossRef]
- Rafiezadeh, Z.; Aboutalebian, S.; Baradaran, A.; Hoveida, L.; Hejazi, S.H. Examining miR-196a, miR-196b and NF-κBIα gene expression in colitis mice model. AMB Express 2025, 15, 98. [Google Scholar] [CrossRef]
- Halstead, S.B.; O’Rourke, E.J.; Allison, A.C. Dengue viruses and mononuclear phagocytes. II. Identity of blood and tissue leukocytes supporting in vitro infection. J. Exp. Med. 1977, 146, 218–229. [Google Scholar] [CrossRef] [PubMed]
- Narayan, R.; Tripathi, S. Intrinsic ADE: The Dark Side of Antibody Dependent Enhancement During Dengue Infection. Front. Cell. Infect. Microbiol. 2020, 10, 580096. [Google Scholar] [CrossRef]
- Edgar, J.E.; Bournazos, S. Fc-FcγR interactions during infections: From neutralizing antibodies to antibody-dependent enhancement. Immunol. Rev. 2024, 328, 221–242. [Google Scholar] [CrossRef]
- Marisa, S.F.; Afsar, N.S.; Sami, C.A.; Hasan, I.; Ali, N.; Rahman, M.M.; Mohona, S.Q.; Ahmed, R.U.; Ehsan, A.; Baset, F. Dengue Myocarditis: A Retrospective Study From 2019 to 2023 in a Tertiary Care Hospital in Bangladesh. Cureus 2025, 17, e84371. [Google Scholar] [CrossRef]
- Anderson, K.B.; Gibbons, R.V.; Thomas, S.J.; Rothman, A.L.; Nisalak, A.; Berkelman, R.L.; Libraty, D.H.; Endy, T.P. Preexisting Japanese encephalitis virus neutralizing antibodies and increased symptomatic dengue illness in a school-based cohort in Thailand. PLoS Neglected Trop. Dis. 2011, 5, e1311. [Google Scholar] [CrossRef]
- Saito, Y.; Moi, M.L.; Takeshita, N.; Lim, C.-K.; Shiba, H.; Hosono, K.; Saijo, M.; Kurane, I.; Takasaki, T. Japanese encephalitis vaccine-facilitated dengue virus infection-enhancement antibody in adults. BMC Infect. Dis. 2016, 16, 578. [Google Scholar] [CrossRef]
- Zhang, W.; Xu, Y.; Zhao, F.; Tarbe, M.; Zhou, S.; Wang, W.; Zhang, S.; Zhang, W.; Xu, Q.; Shi, L.; et al. The pre-existing cellular immunity to Japanese encephalitis virus heterotypically protects mice from Zika virus infection. Sci. Bull. 2020, 65, 402–409. [Google Scholar] [CrossRef]
- Shukla, R.; Beesetti, H.; Brown, J.A.; Ahuja, R.; Ramasamy, V.; Shanmugam, R.K.; Poddar, A.; Batra, G.; Krammer, F.; Lim, J.K.; et al. Dengue and Zika virus infections are enhanced by live attenuated dengue vaccine but not by recombinant DSV4 vaccine candidate in mouse models. eBioMedicine 2020, 60, 102991. [Google Scholar] [CrossRef]
- Garg, H.; Yeh, R.; Watts, D.M.; Mehmetoglu-Gurbuz, T.; Resendes, R.; Parsons, B.; Gonzales, F.; Joshi, A. Enhancement of Zika virus infection by antibodies from West Nile virus seropositive individuals with no history of clinical infection. BMC Immunol. 2021, 22, 5. [Google Scholar] [CrossRef]
- Wen, J.; Cheng, Y.; Ling, R.; Dai, Y.; Huang, B.; Huang, W.; Zhang, S.; Jiang, Y. Antibody-dependent enhancement of coronavirus. Int. J. Infect. Dis. 2020, 100, 483–489. [Google Scholar] [CrossRef]
- Krilov, L.R.; Anderson, L.J.; Marcoux, L.; Bonagura, V.R.; Wedgwood, J.F. Antibody-mediated enhancement of respiratory syncytial virus infection in two monocyte/macrophage cell lines. J. Infect. Dis. 1989, 160, 777–782. [Google Scholar] [CrossRef]
- Polack, F.P. Atypical measles and enhanced respiratory syncytial virus disease (ERD) made simple. Pediatr. Res. 2007, 62, 111–115. [Google Scholar] [CrossRef]
- Kuzmina, N.A.; Younan, P.; Gilchuk, P.; Santos, R.I.; Flyak, A.I.; Ilinykh, P.A.; Huang, K.; Lubaki, N.M.; Ramanathan, P.; Crowe, J.E.; et al. Antibody-Dependent Enhancement of Ebola Virus Infection by Human Antibodies Isolated from Survivors. Cell. Rep. 2018, 24, 1802–1815.e5. [Google Scholar] [CrossRef]
- Lum, F.-M.; Couderc, T.; Chia, B.-S.; Ong, R.-Y.; Her, Z.; Chow, A.; Leo, Y.-S.; Kam, Y.-W.; Rénia, L.; Lecuit, M.; et al. Antibody-mediated enhancement aggravates chikungunya virus infection and disease severity. Sci. Rep. 2018, 8, 1860. [Google Scholar] [CrossRef]
- Lidbury, B.A.; Mahalingam, S. Specific ablation of antiviral gene expression in macrophages by antibody-dependent enhancement of Ross River virus infection. J. Virol. 2000, 74, 8376–8381. [Google Scholar] [CrossRef]
- Saboowala, H. Exploring the Recent Findings on Virally Induced Antibody Dependent Enhancement (ADE) and Potential Mechanisms Leading to This Condition; Notion Press: Chennai, India, 2022. [Google Scholar]
- Kotaki, T.; Nagai, Y.; Yamanaka, A.; Konishi, E.; Kameoka, M. Japanese Encephalitis DNA Vaccines with Epitope Modification Reduce the Induction of Cross-Reactive Antibodies against Dengue Virus and Antibody-Dependent Enhancement of Dengue Virus Infection. Vaccines 2022, 10, 1411. [Google Scholar] [CrossRef]
- Tay, M.Z.; Wiehe, K.; Pollara, J. Antibody-Dependent Cellular Phagocytosis in Antiviral Immune Responses. Front. Immunol. 2019, 10, 332. [Google Scholar] [CrossRef]
- Mayr, L.M.; Su, B.; Moog, C. Non-Neutralizing Antibodies Directed against HIV and Their Functions. Front. Immunol. 2017, 8, 1590. [Google Scholar] [CrossRef]
- Zanella, I.; Degli Antoni, M.; Marchese, V.; Castelli, F.; Quiros-Roldan, E. Non-neutralizing antibodies: Deleterious or propitious during SARS-CoV-2 infection? Int. Immunopharmacol. 2022, 110, 108943. [Google Scholar] [CrossRef]
- He, J.; Kam, Y.-W. Insights from Avian Influenza: A Review of Its Multifaceted Nature and Future Pandemic Preparedness. Viruses 2024, 16, 458. [Google Scholar] [CrossRef] [PubMed]
- Burton, D.R.; Poignard, P.; Stanfield, R.L.; Wilson, I.A. Broadly Neutralizing Antibodies Present New Prospects to Counter Highly Antigenically Diverse Viruses. Science 2012, 337, 183–186. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Kong, W.; Yang, L.; Ding, Y.; Cui, H. Immunity and Immune Evasion Mechanisms of Epstein-Barr Virus. Viral. Immunol. 2023, 36, 303–317. [Google Scholar] [CrossRef] [PubMed]
- Jenks, J.A.; Goodwin, M.L.; Permar, S.R. The roles of host and viral antibody fc receptors in herpes simplex virus (HSV) and human cytomegalovirus (HCMV) infections and immunity. Front. Immunol. 2019, 10, 2110. [Google Scholar] [CrossRef]
- Crispin, M.; Ward, A.B.; Wilson, I.A. Structure and immune recognition of the HIV glycan shield. Annu. Rev. Biophys. 2018, 47, 499–523. [Google Scholar] [CrossRef]
- Walls, A.C.; Park, Y.-J.; Tortorici, M.A.; Wall, A.; McGuire, A.T.; Veesler, D. Structure, Function, and Antigenicity of the SARS-CoV-2 Spike Glycoprotein. Cell 2020, 181, 281–292.e6. [Google Scholar] [CrossRef]
- Engelbrecht, E.; Rodriguez, O.L.; Lees, W.; Vanwinkle, Z.; Shields, K.; Schultze, S.; Gibson, W.S.; Smith, D.R.; Jana, U.; Saha, S.; et al. Germline polymorphism in the immunoglobulin kappa and lambda loci explain variation in the expressed light chain antibody repertoire. bioRxiv 2025. [Google Scholar] [CrossRef]
- Posteraro, B.; Pastorino, R.; Di Giannantonio, P.; Ianuale, C.; Amore, R.; Ricciardi, W.; Boccia, S. The link between genetic variation and variability in vaccine responses: Systematic review and meta-analyses. Vaccine 2014, 32, 1661–1669. [Google Scholar] [CrossRef]
- Garretson, T.A.; Liu, J.; Li, S.H.; Scher, G.; Santos, J.J.S.; Hogan, G.; Vieira, M.C.; Furey, C.; Atkinson, R.K.; Ye, N.; et al. Immune history shapes human antibody responses to H5N1 influenza viruses. Nat. Med. 2025, 31, 1454–1458. [Google Scholar] [CrossRef]
- Auladell, M.; Phuong, H.V.M.; Mai, L.T.Q.; Tseng, Y.-Y.; Carolan, L.; Wilks, S.; Thai, P.Q.; Price, D.; Duong, N.T.; Hang, N.L.K.; et al. Influenza virus infection history shapes antibody responses to influenza vaccination. Nat. Med. 2022, 28, 363–372. [Google Scholar] [CrossRef] [PubMed]
- Lv, H.; Teo, Q.W.; Lee, C.-C.D.; Liang, W.; Choi, D.; Mao, K.J.; Ardagh, M.R.; Gopal, A.B.; Mehta, A.; Szlembarski, M.; et al. Differential antigenic imprinting effects between influenza H1N1 hemagglutinin and neuraminidase in a mouse model. J. Virol. 2025, 99, e0169524. [Google Scholar] [CrossRef] [PubMed]
- Bigdelou, B.; Sepand, M.R.; Najafikhoshnoo, S.; Negrete, J.A.T.; Sharaf, M.; Ho, J.Q.; Sullivan, I.; Chauhan, P.; Etter, M.; Shekarian, T.; et al. COVID-19 and Preexisting Comorbidities: Risks, Synergies, and Clinical Outcomes. Front. Immunol. 2022, 13, 890517. [Google Scholar] [CrossRef] [PubMed]
- Nimmerjahn, F.; Ravetch, J.V. Fcγ receptors as regulators of immune responses. Nat. Rev. Immunol. 2008, 8, 34–47. [Google Scholar] [CrossRef]
- Kang, M.; Lin, F.; Jiang, Z.; Tan, X.; Lin, X.; Liang, Z.; Xiao, C.; Xia, Y.; Guan, W.; Yang, Z.; et al. The impact of pre-existing influenza antibodies and inflammatory status on the influenza vaccine responses in older adults. Influenza Other Respir. Viruses 2023, 17, e13172. [Google Scholar] [CrossRef]
- Rossouw, C.; Ryan, F.J.; Lynn, D.J. The role of the gut microbiota in regulating responses to vaccination: Current knowledge and future directions. FEBS J. 2025, 292, 1480–1499. [Google Scholar] [CrossRef]
- Levy, M.; Kolodziejczyk, A.A.; Thaiss, C.A.; Elinav, E. Dysbiosis and the immune system. Nat. Rev. Immunol. 2017, 17, 219–232. [Google Scholar] [CrossRef]
- Fernandes, K.A.; Lim, A.I. Maternal-driven immune education in offspring. Immunol. Rev. 2024, 323, 288–302. [Google Scholar] [CrossRef]
- Visalli, G.; Laganà, A.; Giudice, D.L.; Calimeri, S.; Caccamo, D.; Trainito, A.; Di Pietro, A.; Facciolà, A. Towards a Future of Personalized Vaccinology: Study on Individual Variables Influencing the Antibody Response to the COVID-19 Vaccine. Vaccines 2023, 11, 217. [Google Scholar] [CrossRef]
- Wrynla, X.H.; Bates, T.A.; Trank-Greene, M.; Wahedi, M.; Hinchliff, A.; Curlin, M.E.; Tafesse, F.G. Immune imprinting and vaccine interval determine antibody responses to monovalent XBB.1.5 COVID-19 vaccination. Commun. Med. 2025, 5, 182. [Google Scholar] [CrossRef]
- Aranow, C. Vitamin D and the Immune System. J. Investig. Med. 2011, 59, 881–886. [Google Scholar] [CrossRef] [PubMed]
- McSorley, H.J.; Maizels, R.M. Helminth infections and host immune regulation. Clin. Microbiol. Rev. 2012, 25, 585–608. [Google Scholar] [CrossRef] [PubMed]
- Li, X.X.; Chen, J.X.; Wang, L.X.; Sun, J.; Chen, S.H.; Chen, J.H.; Zhang, X.Y.; Zhou, X.N. Profiling B and T cell immune responses to co-infection of Mycobacterium tuberculosis and hookworm in humans. Infect. Dis. Poverty 2015, 4, 20. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, J.; Kam, Y.-W.; Lum, F.-M. The Yin and Yang of Antibodies in Viral Infectious Diseases. Diseases 2025, 13, 341. https://doi.org/10.3390/diseases13100341
He J, Kam Y-W, Lum F-M. The Yin and Yang of Antibodies in Viral Infectious Diseases. Diseases. 2025; 13(10):341. https://doi.org/10.3390/diseases13100341
Chicago/Turabian StyleHe, Jianning, Yiu-Wing Kam, and Fok-Moon Lum. 2025. "The Yin and Yang of Antibodies in Viral Infectious Diseases" Diseases 13, no. 10: 341. https://doi.org/10.3390/diseases13100341
APA StyleHe, J., Kam, Y.-W., & Lum, F.-M. (2025). The Yin and Yang of Antibodies in Viral Infectious Diseases. Diseases, 13(10), 341. https://doi.org/10.3390/diseases13100341