
Systematic Evaluation of the Distribution of Immune Cells following Subcutaneous Administration of Haemophilus Influenzae Type B Vaccine to Mice

Abstract
:1. Introduction
2. Results
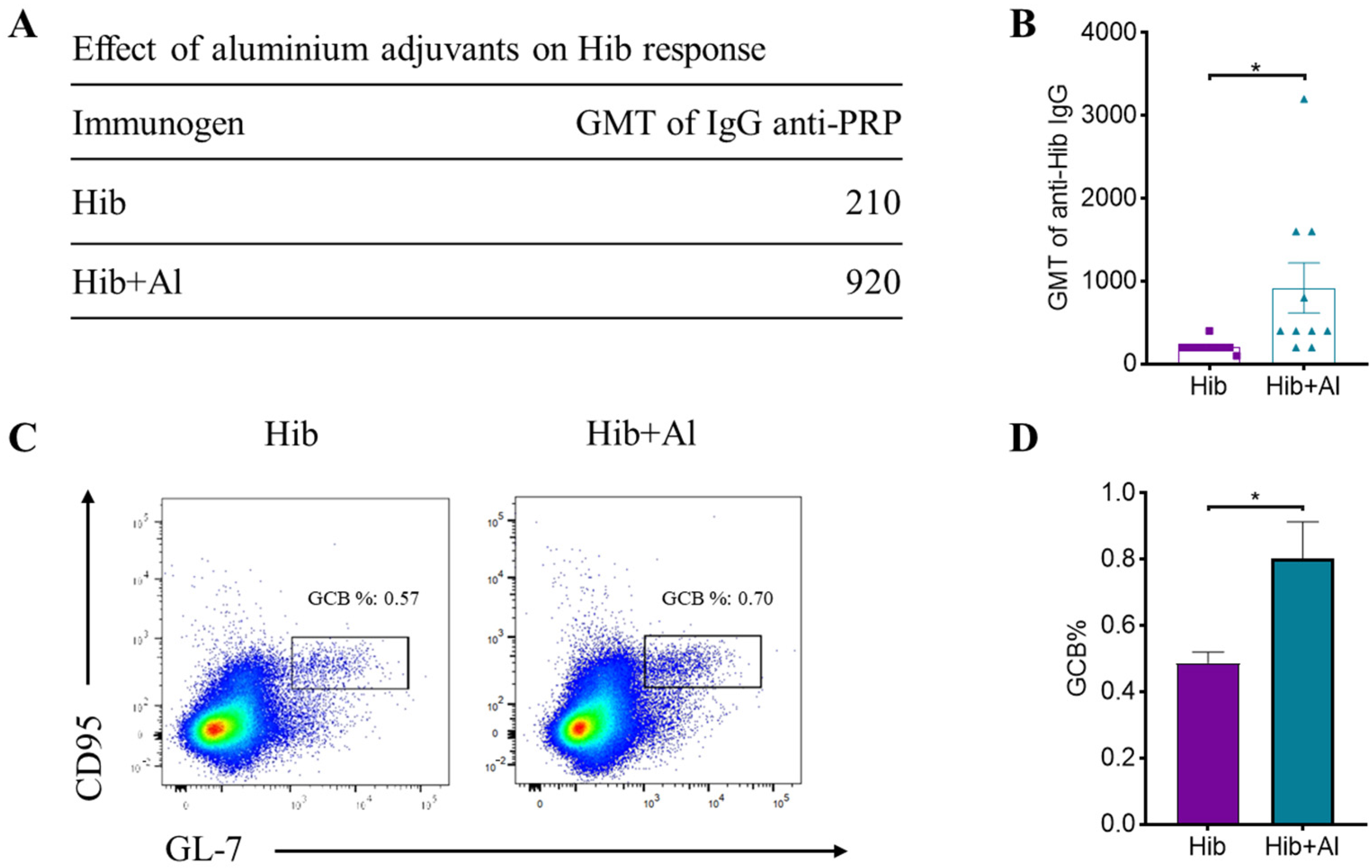
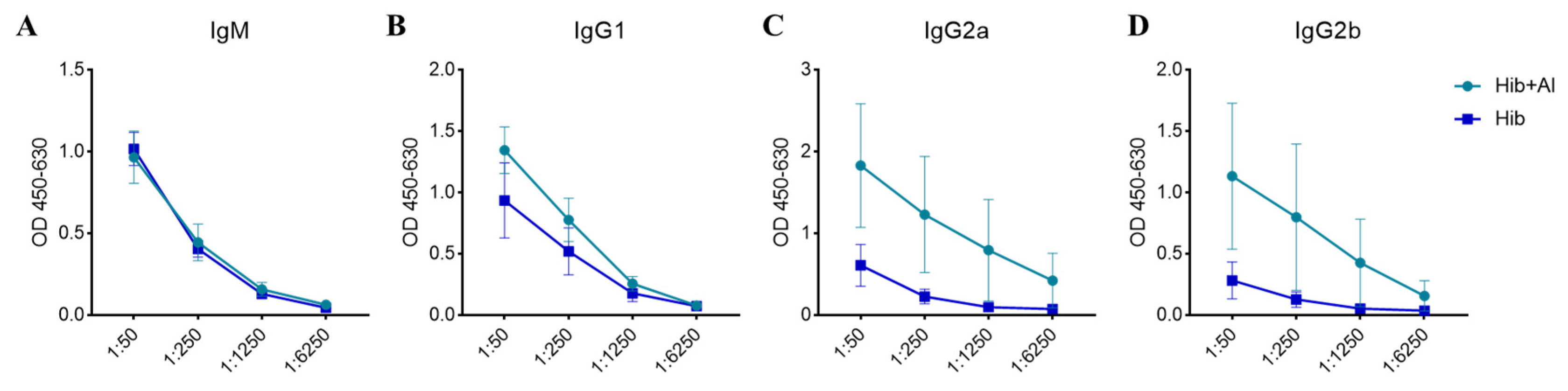
2.1. Hib and Aluminum-Adjuvant-Adsorbed Hib Vaccines Both Induce Humoral Immune Response
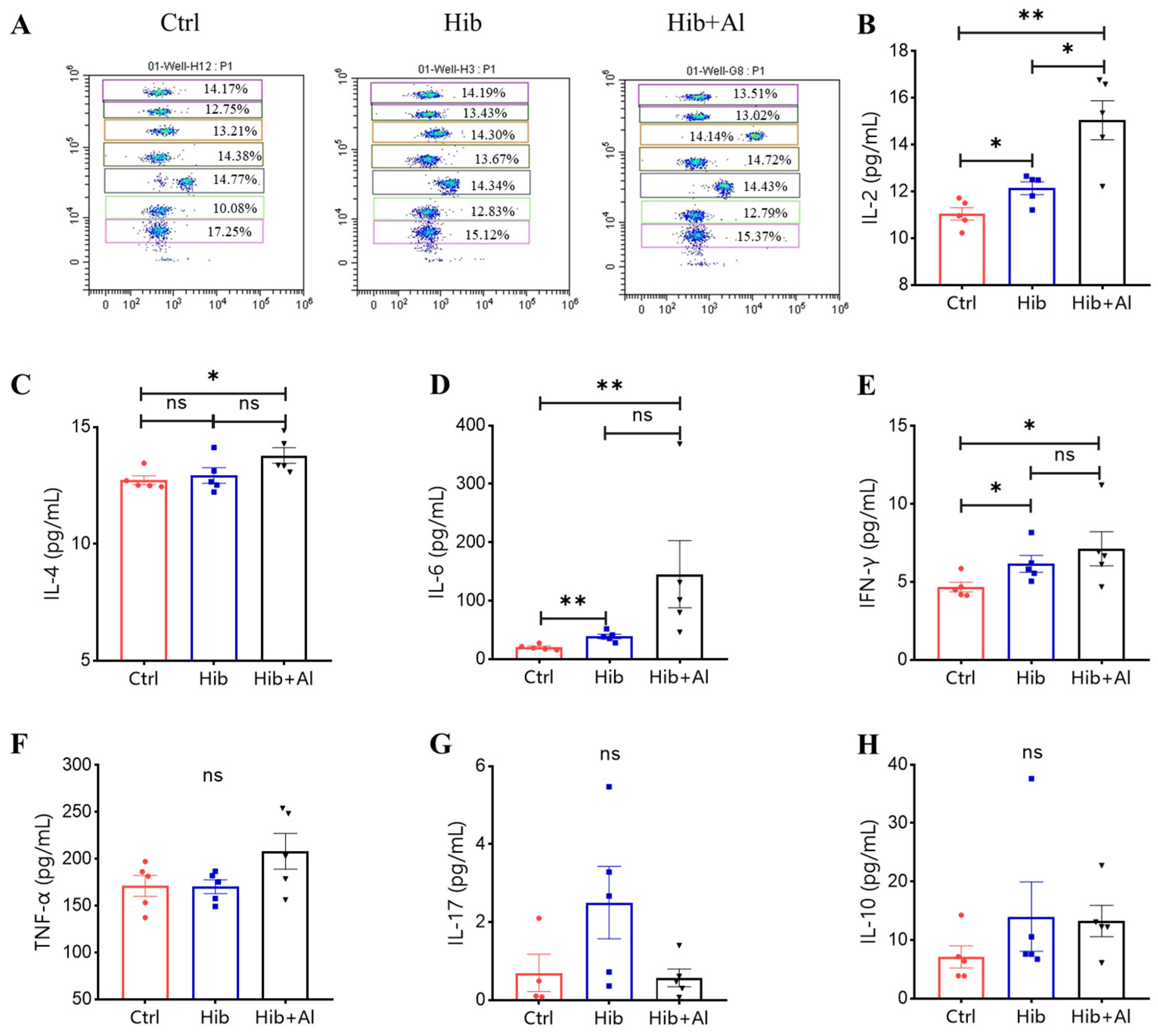
2.2. The Effect of Aluminum Adjuvant on Hib-Induced Cytokine Expression
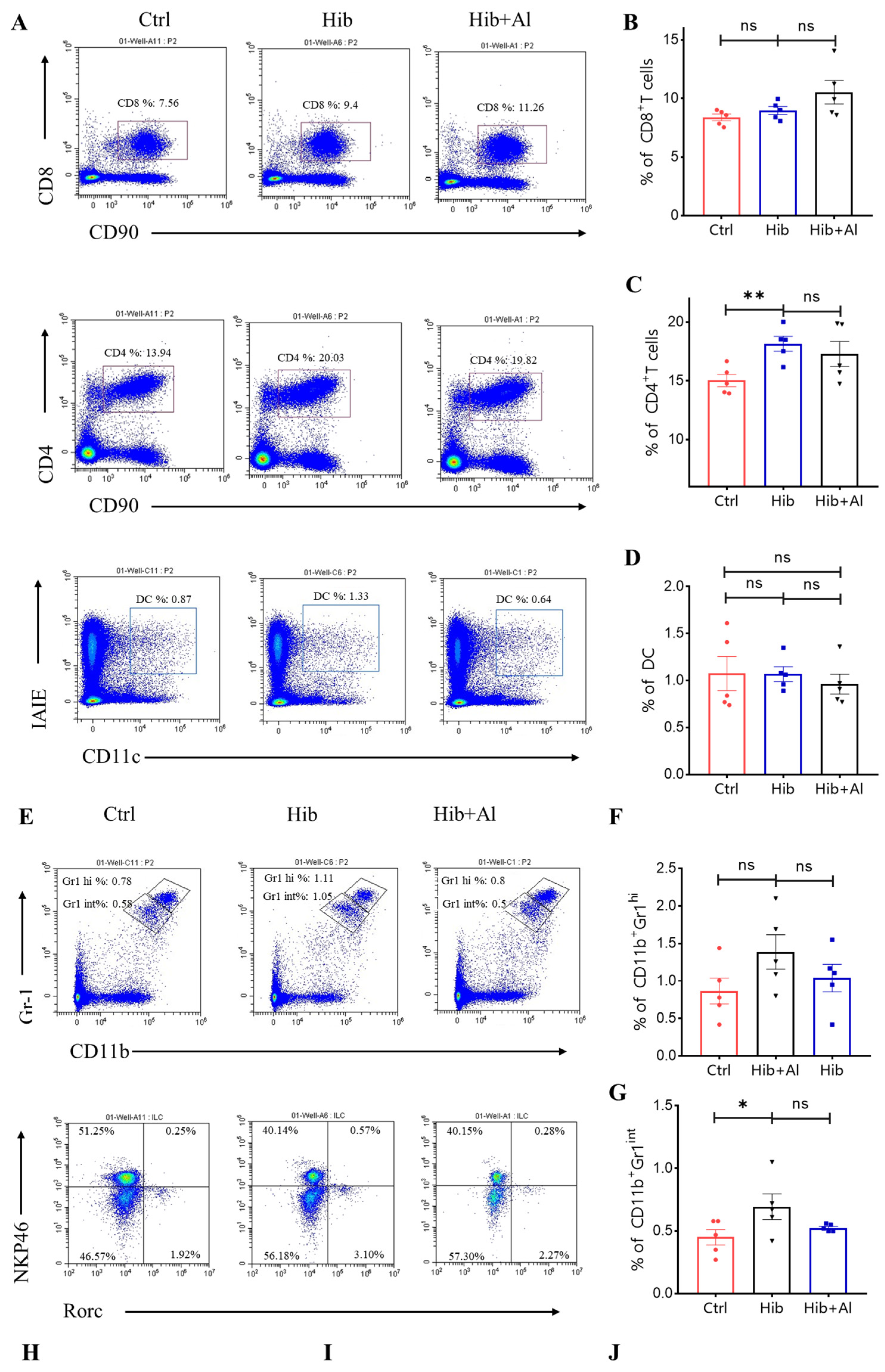
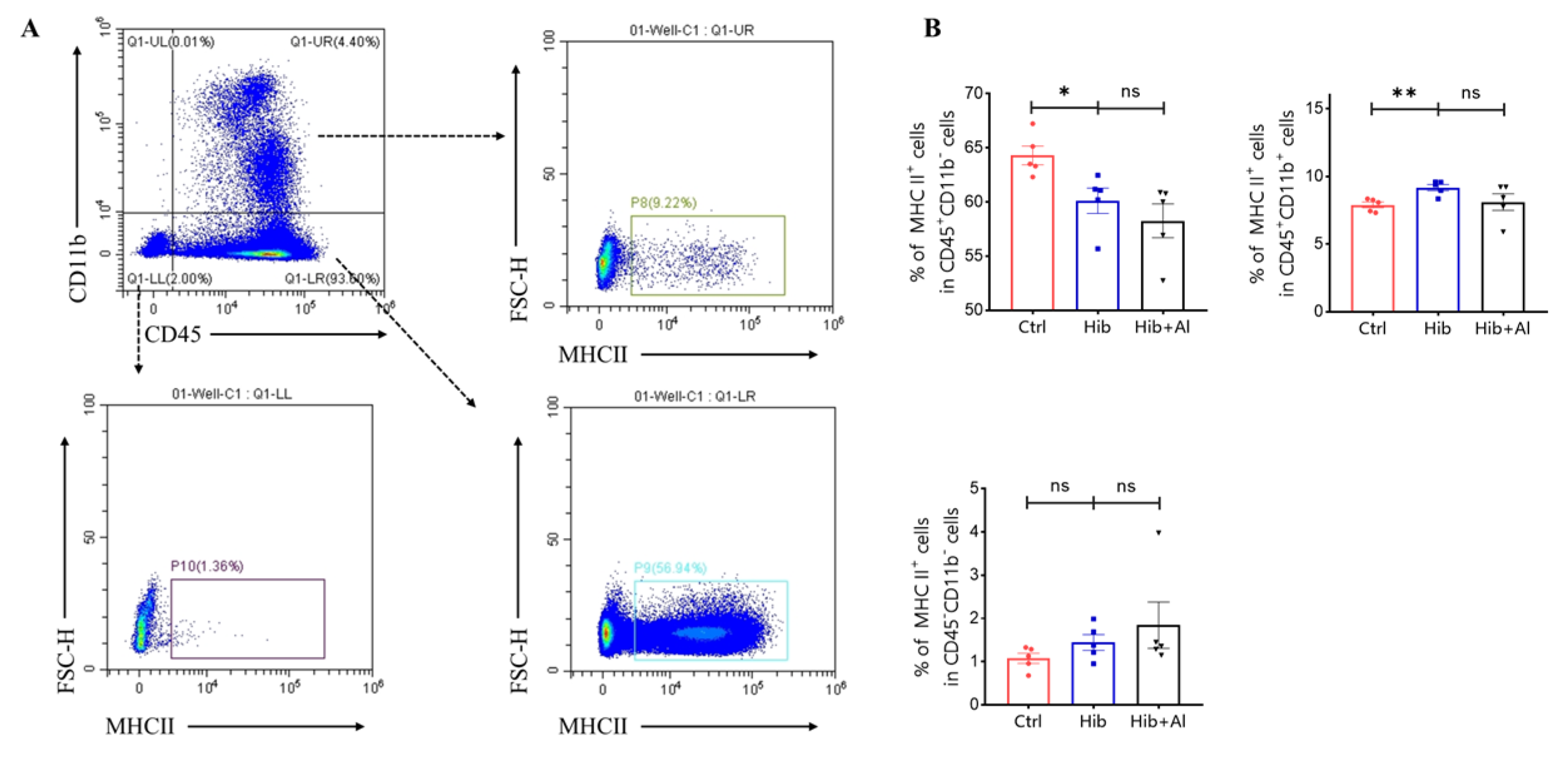
2.3. The Distribution of Immune Cells following Administration of Hib and Hib+Al Vaccine
3. Discussion
4. Materials and Methods
4.1. Vaccines
4.2. Animals and Immunization
4.3. Reagents
4.4. ELISA
4.5. FACS
4.6. ELISpot
4.7. CBA Analysis
4.8. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gilsdorf, J.R. Hib Vaccines: Their Impact on Haemophilus influenzae Type b Disease. J. Infect. Dis. 2021, 224, S321–S330. [Google Scholar] [CrossRef]
- Decker, M.D.; Edwards, K.M. Haemophilus influenzae type b vaccines: History, choice and comparisons. Pediatr. Infect. Dis. J. 1998, 17, S113–S116. [Google Scholar] [CrossRef]
- Mawas, F.; Dickinson, R.; Douglasbardsley, A.; Xing, D.K.; Sesardic, D.; Corbel, M.J. Immune interaction between components of acellular pertussis-diphtheria-tetanus (DTaP) vaccine and Haemophilus influenzae b (Hib) conjugate vaccine in a rat model. Vaccine 2006, 24, 3505–3512. [Google Scholar] [CrossRef]
- Mawas, F.; Newman, G.; Burns, S.; Corbel, M.J. Suppression and modulation of cellular and humoral immune responses to Haemophilus influenzae type B (Hib) conjugate vaccine in hib–diphtheria-tetanus toxoids–acellular pertussis combination vaccines: A study in a rat model. J. Infect. Dis. 2005, 191, 58–64. [Google Scholar] [CrossRef]
- Mulholland, E.K.; Hoestermann, A.; Ward, J.I.; Maine, N.; Ethevenaux, C.; Greenwood, B.M. The use of Haemophilus influenzae type b-tetanus toxoid conjugate vaccine mixed with diphtheria-tetanus-pertussis vaccine in Gambian infants. Vaccine 1996, 14, 905–909. [Google Scholar] [CrossRef]
- Cherian, T.; Thomas, N.; Raghupathy, P.; Durot, I.; Dutta, A. Safety and immunogenicity of Haemophilus influenzae type B vaccine given in combination with DTwP at 6, 10 and 14 weeks of age. Indian Pediatr. 2002, 39, 427–436. [Google Scholar]
- Tavakoli, M.; Bouzari, S.; Jafari, A.; Oloomi, M.; Karam, M.R.A.; Najar-Peerayeh, S.; Siadat, S.D. Effect of nontypeable Haemophilus influenzae protein E (PE) as a microbial adjuvant on the amount of antibody against PRP of Haemophilus influenzae type b (Hib) in BALB/c mice. Microb. Pathog. 2019, 129, 78–81. [Google Scholar] [CrossRef]
- Gupta, R.K.; Anderson, R.; Cecchini, D.; Rost, B.; Xu, J.; Gendreau, K.; Saroff, D.L.; Marchant, C.; Siber, G.R. Evaluation of a guinea pig model to assess interference in the immunogenicity of different components of a combination vaccine comprising diphtheria, tetanus and acellular pertussis (DTaP) vaccine and haemophilus influenzae type b capsular polysaccharide conjugate vaccine. Biologicals 1999, 27, 167–176. [Google Scholar] [CrossRef]
- Gupta, R.; Anderson, R.; Cecchini, D.; Rost, B.; Griffin, P.; Benscoter, K.; Xu, J.; Montanez-Ortiz, L.; Siber, G.R. Development of a guinea-pig model for potency/immunogenicity evaluation of diphtheria, tetanus acellular pertussis (DTaP) and Haemophilus influenzae type b polysaccharide conjugate vaccines. Dev. Biol. Stand. 1996, 86, 283–296. [Google Scholar]
- He, X.; Wang, J.; Tang, Y.; Chiang, S.T.; Han, T.; Chen, Q.; Qian, C.; Shen, X.; Li, R.; Ai, X. Recent Advances of Emerging Spleen-Targeting Nanovaccines for Immunotherapy. Adv. Healthc. Mater. 2023, 12, e2300351. [Google Scholar] [CrossRef]
- Wang, Y.; Qu, K.; Lu, W.; Zhao, P.; Wang, Z.; Cui, C.; Liu, Y.; Yang, M.; Yu, Y.; Wang, L. Neutrophils recruited to immunization sites initiating vaccine-induced antibody responses by locally expressing BAFF. iScience 2022, 25, 104453. [Google Scholar] [CrossRef]
- Anderson, P.; Smith, D.H.; Ingram, D.L.; Wilkins, J.; Wehrle, P.F.; Howie, V.M. Antibody of polyribophate of Haemophilus influenzae type b in infants and children: Effect of immunization with polyribophosphate. J. Infect. Dis. 1977, 136, S57–S62. [Google Scholar] [CrossRef] [PubMed]
- Kalies, H.; Grote, V.; Verstraeten, T.; Hessel, L.; Schmitt, H.-J.; von Kries, R. The use of combination vaccines has improved timeliness of vaccination in children. Pediatr. Infect. Dis. J. 2006, 25, 507–512. [Google Scholar] [CrossRef] [PubMed]
- Malek, T.R. The biology of interleukin-2. Annu. Rev. Immunol. 2008, 26, 453–479. [Google Scholar] [CrossRef]
- Poolman, J.; Kaufhold, A.; De Grave, D.; Goldblatt, D. Clinical relevance of lower Hib response in DTPa-based combination vaccines. Vaccine 2001, 19, 2280–2285. [Google Scholar] [CrossRef] [PubMed]
- Slack, M.H.; Schapira, D.; Thwaites, R.J.; Burrage, M.; Southern, J.; Andrews, N.; Borrow, R.; Goldblatt, D.; Miller, E. Immune response of premature infants to meningococcal serogroup C and combined diphtheria-tetanus toxoids-acellular pertussis-Haemophilus influenzae type b conjugate vaccines. J. Infect. Dis. 2001, 184, 1617–1620. [Google Scholar] [CrossRef]
- Bernstein, H.H.; Seyferth, E.R. Hib antibody responses in infants following diphtheria, tetanus, acellular pertussis, and conjugated Haemophilus influenzae type b (Hib) combination vaccines with decreasing amounts of tetanus toxoid. Vaccine 2017, 35, 6707–6711. [Google Scholar] [CrossRef]
- Tafreshi, S.-Y.H. Efficacy, safety, and formulation issues of the combined vaccines. Expert Rev. Vaccines 2020, 19, 949–958. [Google Scholar] [CrossRef]
- Konini, A.; Kang, M.; Moghadas, S.M. Simulating Immune Interference on the Effect of a Bivalent Glycoconjugate Vaccine against Haemophilus influenzae Serotypes “a” and “b”. Can. J. Infect. Dis. Med. Microbiol. 2016, 2016, 5486869. [Google Scholar] [CrossRef]
- Benne, C.; Harmsen, M.; van der Graaff, W.; Verheul, A.; Snippe, H.; Kraaijeveld, C. Influenza virus neutralizing antibodies and IgG isotype profiles after immunization of mice with influenza A subunit vaccine using various adjuvants. Vaccine 1997, 15, 1039–1044. [Google Scholar] [CrossRef]
- Hocart, M.J.; Mackenzie, J.S.; Stewart, G.A. The immunoglobulin G subclass responses of mice to influenza a virus: The effect of mouse strain, and the neutralizing abilities of individual protein a-purified subclass antibodies. J. Gen. Virol. 1989, 70 Pt 9, 2439–2448. [Google Scholar] [CrossRef]
- Snapper, C.M.; Paul, W.E. Interferon-gamma and B cell stimulatory factor-1 reciprocally regulate Ig isotype production. Science 1987, 236, 944–947. [Google Scholar] [CrossRef]
- Mbulaiteye, S.; Kemp, T.; Gage, J.; Ajenifuja, K.; Kiruthu, C.; Wentzensen, N.; Adepiti, C.; Wacholder, S.; Burk, R.; Schiffman, M.; et al. Plasma cytokine levels and human papillomavirus infection at the cervix in rural Nigerian women. Cytokine 2013, 64, 146–151. [Google Scholar] [CrossRef]
- Tchalla, E.Y.; Bhalla, M.; Wohlfert, E.A.; Ghanem, E.N.B. Neutrophils Are Required during Immunization with the Pneumococcal Conjugate Vaccine for Protective Antibody Responses and Host Defense Against Infection. J. Infect. Dis. 2020, 222, 1363–1370. [Google Scholar] [CrossRef]
- Hepworth, M.R.; Monticelli, L.A.; Fung, T.C.; Ziegler, C.G.K.; Grunberg, S.; Sinha, R.; Mantegazza, A.R.; Ma, H.-L.; Crawford, A.; Angelosanto, J.M.; et al. Innate lymphoid cells regulate CD4+ T-cell responses to intestinal commensal bacteria. Nature 2013, 498, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Sonnenberg, G.F.; Fung, T.C.; Goc, J.; Wang, X.; Hepworth, M.R. Group 3 innate lymphoid cells mediate intestinal selection of commensal bacteria-specific CD4+ T cells. J. Immunol. 2015, 348, 1031–1035. [Google Scholar] [CrossRef]
- Wang, W.; Li, Y.; Hao, J.; He, Y.; Dong, X.; Fu, Y.-X.; Guo, X. The Interaction between Lymphoid Tissue Inducer-Like Cells and T Cells in the Mesenteric Lymph Node Restrains Intestinal Humoral Immunity. Cell Rep. 2020, 32, 107936. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Tan, W.; Lou, Z.; Huang, B.; Zhou, W.; Zhao, Y.; Zhang, J.; Liang, H.; Li, N.; Zhu, X.; et al. Immunogenicity Evaluating of the Multivalent COVID-19 Inactivated Vaccine against the SARS-CoV-2 Variants. Vaccines 2022, 10, 956. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment | Group | Total Number | Tissue |
|---|---|---|---|
| Immunization experiments | Hib+Al | 10 | / |
| Hib | 10 | ||
| Ctrl | 10 | ||
| Total anti-PRP IgG analysis | Hib+Al | 10 | Serum |
| Hib | 10 | ||
| Antibody analysis of different subtypes | Hib+Al | 5 | Serum |
| Hib | 5 | ||
| CBA | Hib+Al | 5 | Serum |
| Hib | 5 | ||
| Ctrl | 5 | ||
| ELISpot | Hib+Al | 5 | Spleen |
| Hib | 5 | ||
| Ctrl | 5 | ||
| FACS | Other immune cells analysis | 5 | Spleen |
| GCB analysis | 4 |
| Reagents | Source | Cat# |
|---|---|---|
| Purified anti-mouse CD16/32 Antibody | BioLegend (San Diego, CA, USA) | 101302 |
| PE/Cyanine7 anti-mouse CD3ε Antibody | BioLegend (San Diego, CA, USA) | 100320 |
| APC/Cyanine7 anti-mouse CD45 Antibody | BioLegend (San Diego, CA, USA) | 103116 |
| Alexa Fluor 700 anti-mouse CD4 Antibody | BioLegend (San Diego, CA, USA) | 100430 |
| Brilliant Violet 421 anti-mouse/human CD45R/B220 Antibody | BioLegend (San Diego, CA, USA) | 103240 |
| FITC anti-mouse/human GL7 Antigen Antibody | BioLegend (San Diego, CA, USA) | 144604 |
| PE anti-mouse CD95 (Fas) Antibody | BioLegend (San Diego, CA, USA) | 152608 |
| Alexa Fluor 700 anti-mouse CD45 Antibody | BioLegend (San Diego, CA, USA) | 103128 |
| FITC anti-mouse/human CD11b Antibody | BioLegend (San Diego, CA, USA) | 101206 |
| PE anti-mouse Ly-6G/Ly-6C (Gr-1) Antibody | BioLegend (San Diego, CA, USA) | 108408 |
| PE/Cyanine7 anti-mouse CD11c Antibody | BioLegend (San Diego, CA, USA) | 117318 |
| PE anti-mouse CD90.2 (Thy-1.2) Antibody | BioLegend (San Diego, CA, USA) | 140308 |
| FITC anti-mouse CD3 Antibody | BioLegend (San Diego, CA, USA) | 100204 |
| Brilliant Violet 421 anti-mouse NKp46 Antibody | BioLegend (San Diego, CA, USA) | 137611 |
| ROR gamma (t) PE-Cyanine7 | ThermoFisher (Waltham, MA, USA) | 562607 |
| IL-4 ELISpot | Dakewe Group (Shenzhen, China) | 221042 |
| IFN-g ELISpot | Dakewe Group (Shenzhen, China) | 221005 |
| Mouse Th1/Th2/Th17 CBA Kit | BD (San Jose, CA, USA) | 560485 |
| Anti-IgG2a antibody | Abcam (Cambridge, UK) | ab170489 |
| Anti-IgG2b antibody | Abcam (Cambridge, UK) | ab170322 |
| Goat Anti-Mouse IgG1 | Abcam (Cambridge, UK) | ab97240 |
| Anti-IgM antibody | Abcam (Cambridge, UK) | ab106742 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, Y.; Zhao, Y.; Liang, H.; Wang, X.; Lan, H.; Tian, D.; Li, Y.; Wang, H. Systematic Evaluation of the Distribution of Immune Cells following Subcutaneous Administration of Haemophilus Influenzae Type B Vaccine to Mice. Diseases 2023, 11, 139. https://doi.org/10.3390/diseases11040139
He Y, Zhao Y, Liang H, Wang X, Lan H, Tian D, Li Y, Wang H. Systematic Evaluation of the Distribution of Immune Cells following Subcutaneous Administration of Haemophilus Influenzae Type B Vaccine to Mice. Diseases. 2023; 11(4):139. https://doi.org/10.3390/diseases11040139
Chicago/Turabian StyleHe, Yao, Yuxiu Zhao, Hongyang Liang, Xue Wang, Haoyue Lan, Dongyang Tian, Yan Li, and Hui Wang. 2023. "Systematic Evaluation of the Distribution of Immune Cells following Subcutaneous Administration of Haemophilus Influenzae Type B Vaccine to Mice" Diseases 11, no. 4: 139. https://doi.org/10.3390/diseases11040139
APA StyleHe, Y., Zhao, Y., Liang, H., Wang, X., Lan, H., Tian, D., Li, Y., & Wang, H. (2023). Systematic Evaluation of the Distribution of Immune Cells following Subcutaneous Administration of Haemophilus Influenzae Type B Vaccine to Mice. Diseases, 11(4), 139. https://doi.org/10.3390/diseases11040139