
Functional Connectivity of EEG in Encephalitis during Slow Biphasic Complexes



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Methods
2.1. Experimental Data
- 0—normal condition (22 traces, from either controls or patients when they presented no sign of disease);
- 1—mild disease (29 traces);
- 2—moderate disease (25 traces);
- 3—severe condition (22 traces);
- 4—serious pathology (20 traces);
- NC—not classified EEG traces, considered as unmanageable (10 traces).
2.2. Automated Detection of SBCs
2.3. Functional Connectivity Indexes
2.4. Topological Indexes of Network Patterns
- Strength is the sum of the connections to a nodewhere is an element of the adjacency matrix indicating the connection status between nodes i and j. We used the average strength, defined as(where n is the number of nodes in the graph) to provide an overall characterization of how much the network was connected.
- Clustering coefficient of the network [41]where is the geometric mean of triangles around a node iand is the clustering coefficient of the ith node ( = 0 for 2). It is a measure of the degree to which nodes in a graph cluster together.
- Global efficiency of the network [42]where is the efficiency of node i and is the shortest path length between nodes i and jwhere is the geodesic path between nodes i and j (with for disconnected pairs). It measures the efficiency of the network to exchange information.
- Characteristic path length of the network (indicated as lambda in the following)where is the average distance between node i and all other nodes. As for the previous index, it is also a measure of information transport in the network (with the opposite trend with respect to the efficiency), as a short average path length between nodes indicates a better overall connection.
2.5. Statistical Analysis
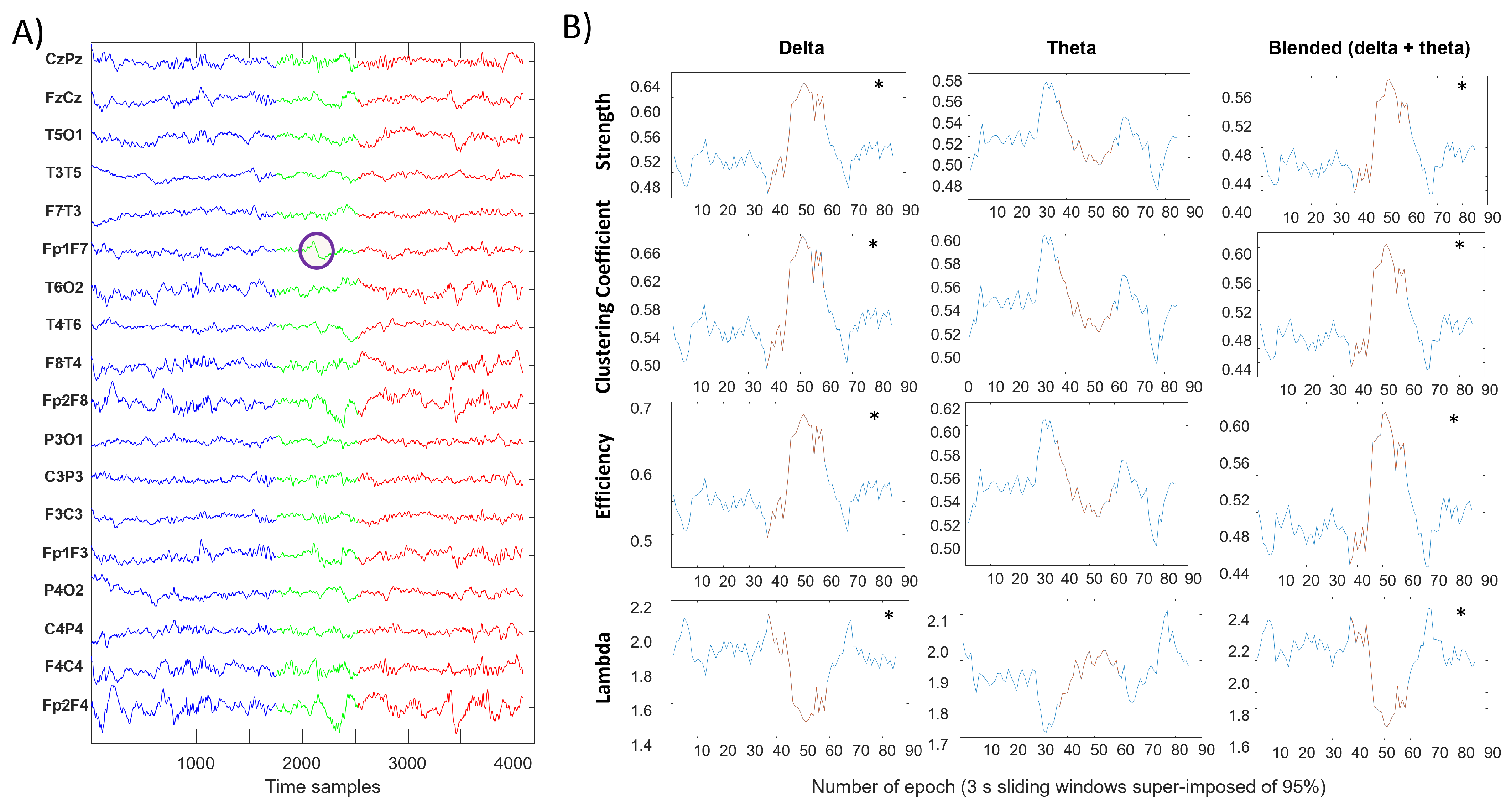
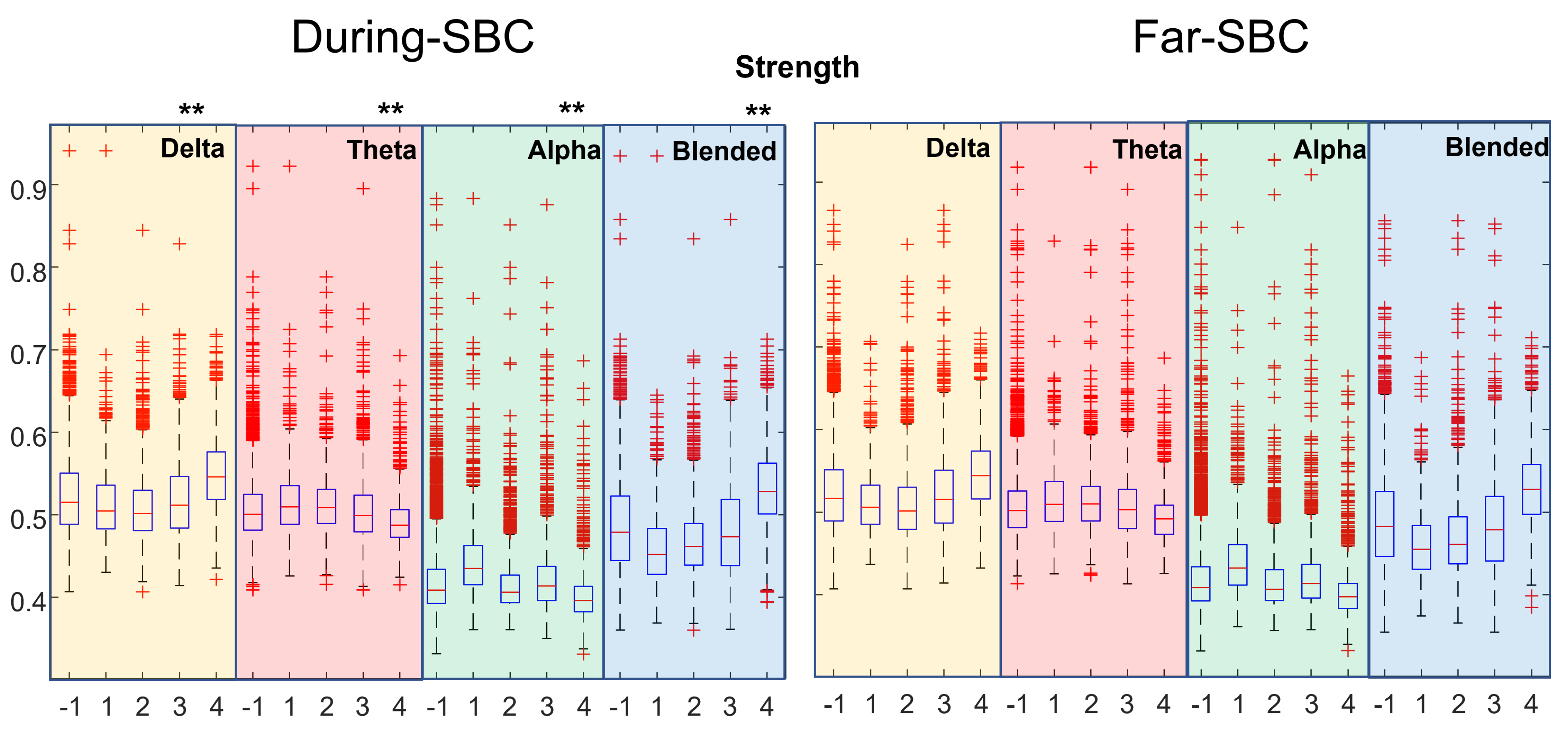
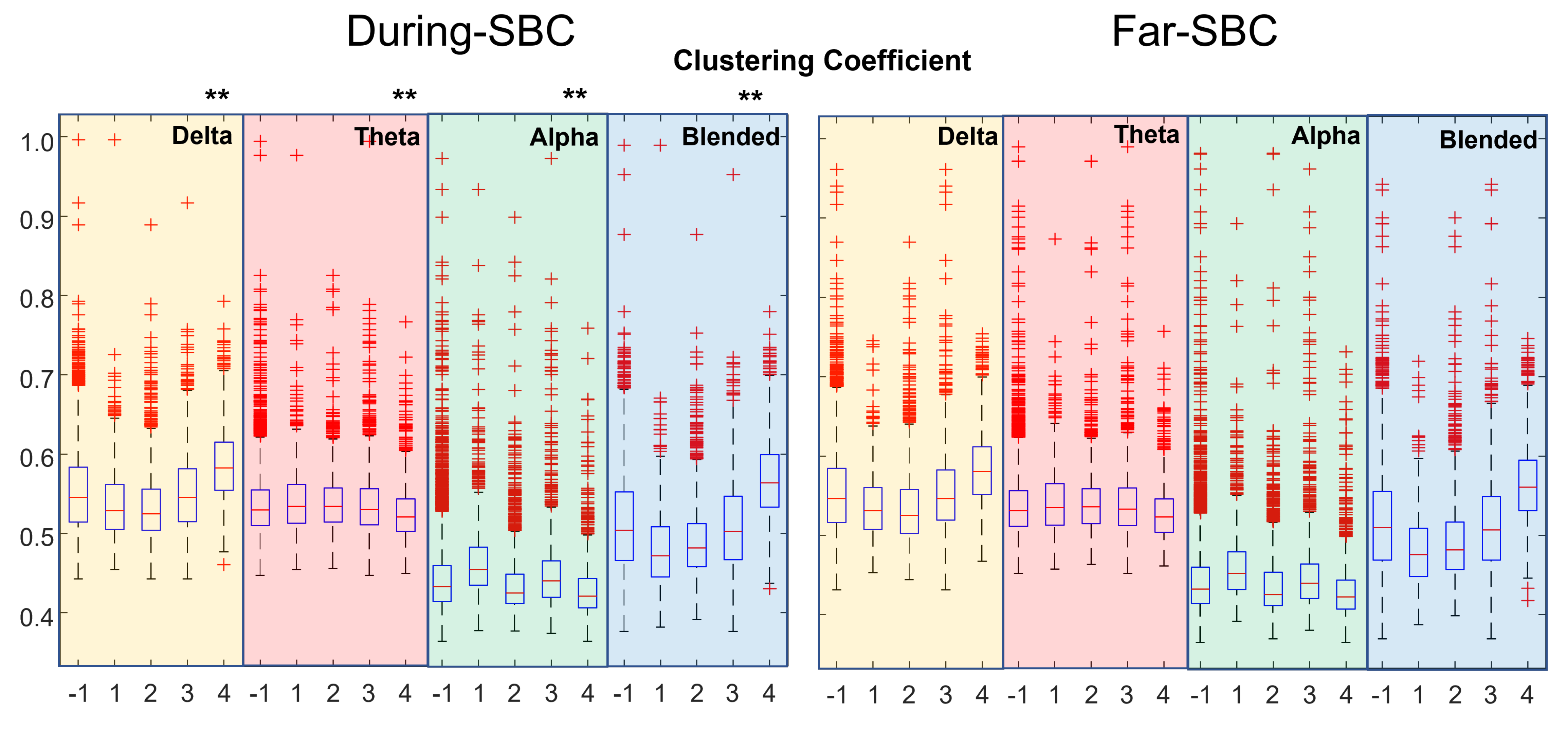
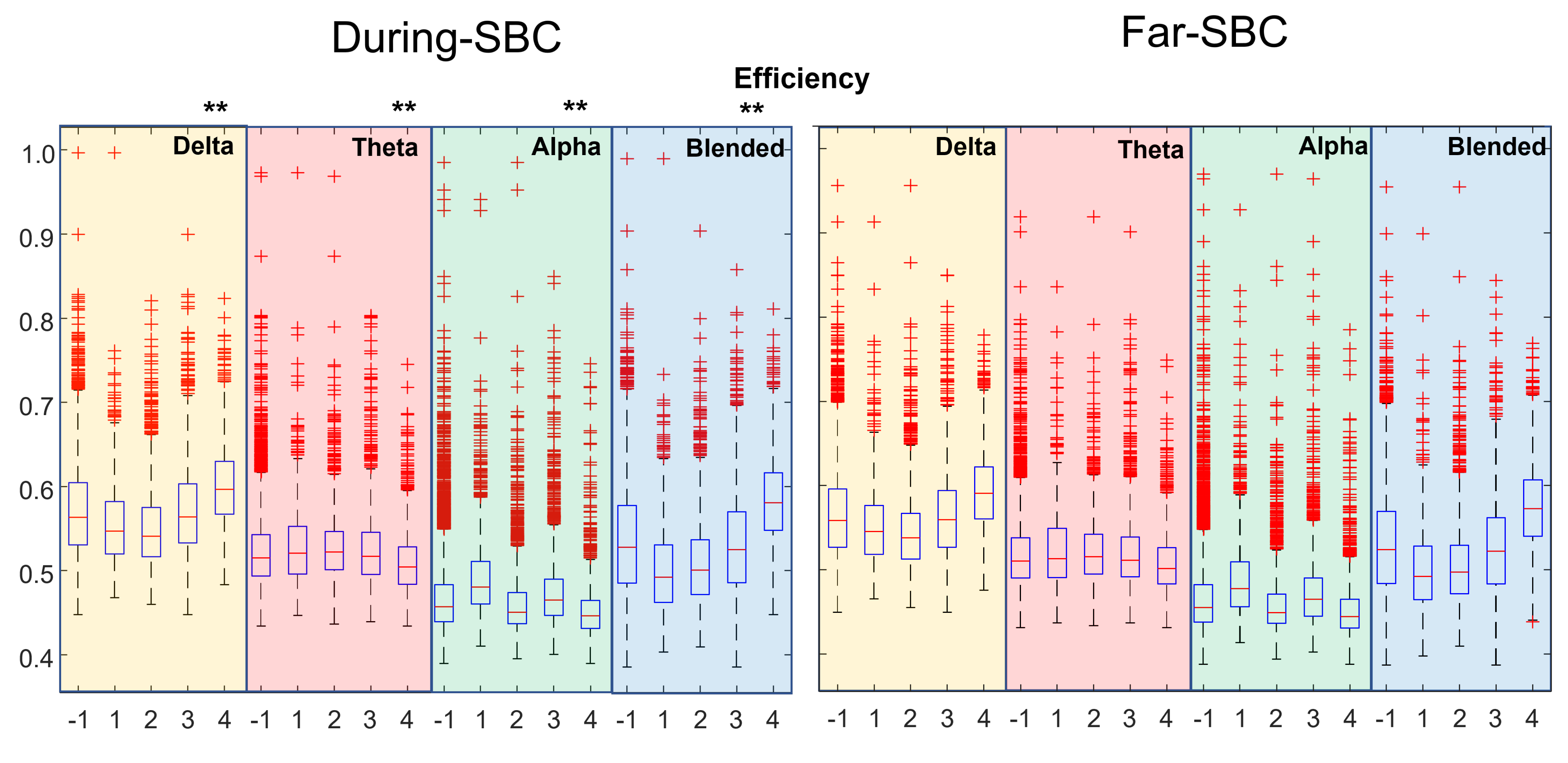
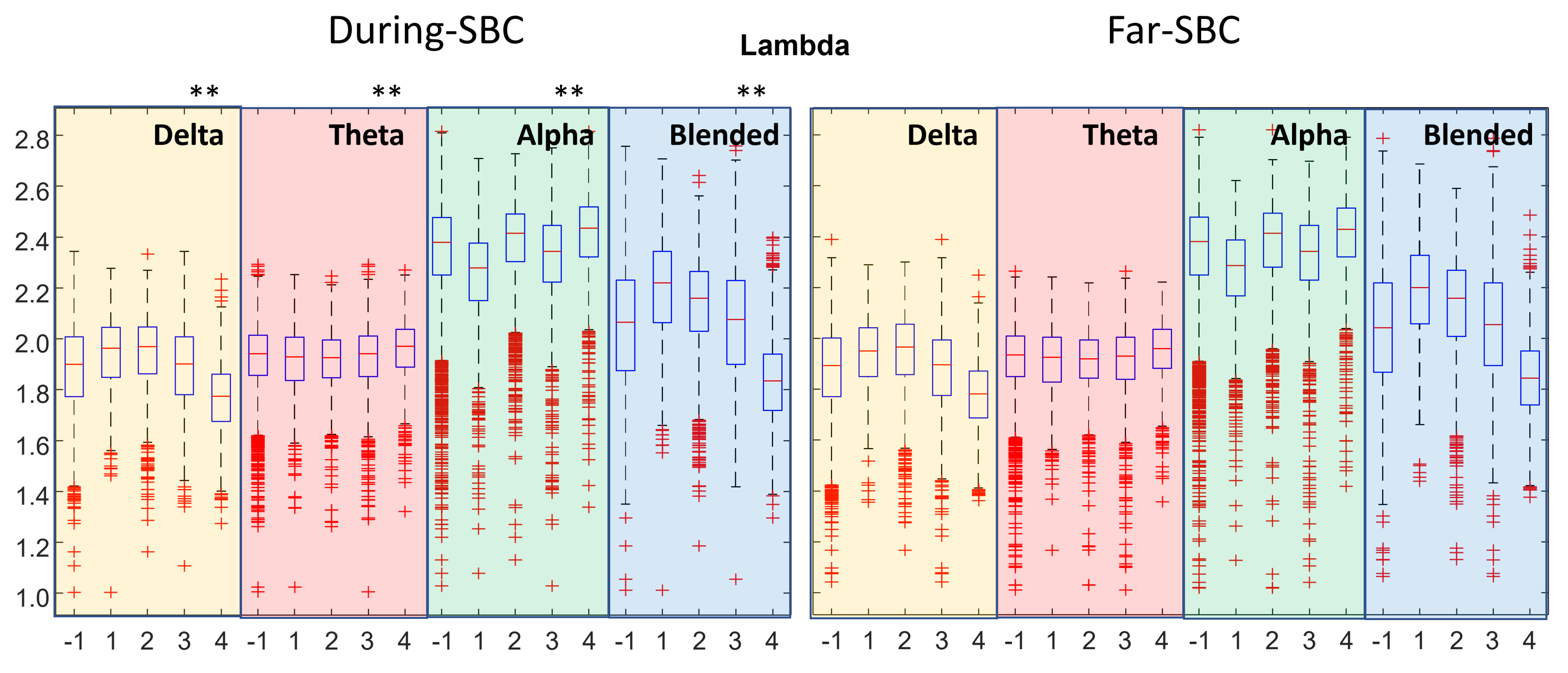
3. Results
4. Discussion
- clustering coefficient, efficiency and strength tend to be higher when closer to an SBC;
- the characteristic path length (lambda) is lower when near an SBC.
- This is a retrospective clinical study, including also data recorded in emergency conditions or quick checkups, considering a different number of channels. Thus, the processed EEGs are not very clean, and the experimental conditions were not kept under control. Moreover, young children with a large range of ages were included. Hence, the investigated brains were under development, with large differences, beyond the effect of the pathology.
- Our automated method for SBC detection is based on the subjective experience of neurologists. The identified waveforms are sometimes small, raising doubts on their correctness, and some of them are not correct in the opinion of the expert neurologist (as shown in some examples reported in [28]). Due to the long duration of the traces, a complete comparison of the automated and manual identification is not available, but based on sampled portions of traces, we could expect that our algorithm makes about 25% of false positive and 10% of false negatives, with a large variability of accuracy depending on the quality of the recording.
- The distributions of topological indexes far from and close to SBCs are quite similar. Only paired comparisons of the indexes during the SBC and in the preceding time window show statistically significant variations. Thus, the brain behavior changes during the SBC with respect to the previous baseline, but the variations between the overall epochs taken during SBCs and far from them are less clear.
- Only the crosscorrelation of EEG rhythms was considered as a measure of functional connectivity.
5. Conclusions and Further Work
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| EEG | Electroencephalogram |
| FC | Functional Connectivity |
| SBC | Slow Biphasic Complex |
| CC | Crosscorrelation |
References
- Barbadoro, P.; Marigliano, A.; Ricciardi, A.; D’Errico, M.M.; Prospero, E. Trend of hospital utilization for encephalitis. Epidemiol. Infect. 2012, 140, 753–764. [Google Scholar] [CrossRef]
- Falchek, S.J. Encephalitis in the Pediatric Population. Pediatr. Rev. 2012, 33, 122–133. [Google Scholar] [CrossRef][Green Version]
- Piquet, A.L.; Cho, T.A. The Clinical Approach to Encephalitis. Curr. Neurol. Neurosci. Rep. 2016, 16, 45. [Google Scholar] [CrossRef]
- Thompson, C.; Kneen, R.; Riordan, A.; Kelly, D.; Pollard, A.J. Encephalitis in children. Arch. Dis. Child. 2012, 97, 150–161. [Google Scholar] [CrossRef]
- Venkatesan, A. Epidemiology and outcomes of acute encephalitis. Curr. Opin. Neurol. 2015, 28, 277–282. [Google Scholar] [CrossRef]
- Boucher, A.; Herrmann, J.L.; Morand, P.; Buzelé, R.; Crabol, Y.; Stahl, J.P.; Mailles, A. Epidemiology of infectious encephalitis causes in 2016. Med. Mal. Infect. 2017, 47, 221–235. [Google Scholar] [CrossRef] [PubMed]
- Weingarten, L.; Enarson, P.; Klassen, T. Encephalitis. Pediatr. Emerg. Care 2013, 29, 235–241. [Google Scholar] [CrossRef]
- Vora, N.M.; Holman, R.C.; Mehal, J.M.; Steiner, C.A.; Blanton, J.; Sejvar, J. Burden of encephalitis-associated hospitalizations in the United States, 1998–2010. Neurology 2014, 82, 443–451. [Google Scholar] [CrossRef]
- Kneen, R.; Michael, B.D.; Menson, E.; Mehta, B.; Easton, A.; Hemingway, C.; Klapper, P.E.; Vincent, A.; Lim, M.; Carrol, E.; et al. National Encephalitis Guidelines Development and Stakeholder Groups. Management of suspected viral encephalitis in children—Association of British Neurologists and British Paediatric Allergy, Immunology and Infection Group national guidelines. J. Infect. 2012, 64, 449–477. [Google Scholar] [CrossRef] [PubMed]
- Drislane, F.W. Overlap of encephalopathies and epileptic seizures. J. Clin. Neurophysiol. 2013, 30, 468–476. [Google Scholar] [CrossRef] [PubMed]
- Sutter, R.; Kaplan, P.W. Clinical and electroencephalographic correlates of acute encephalopathy. J. Clin. Neurophysiol. 2013, 30, 443–453. [Google Scholar] [CrossRef]
- Tauber, S.C.; Eiffert, H.; Bruck, W.; Nau, R. Septic encephalopathy and septic encephalitis. Expert Rev. Anti Infect. Ther. 2017, 15, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Sutter, R.; Kaplan, P.W.; Cervenka, M.C.; Thakur, K.T.; Asemota, A.O.; Venkatesan, A.; Geocadin, R.G. Electroencephalography for diagnosis and prognosis of acute encephalitis. Clin. Neurophysiol. 2015, 126, 1524–1531. [Google Scholar] [CrossRef] [PubMed]
- Mesin, L.; Costa, P. Prognostic value of EEG indexes for the Glasgow outcome scale of comatose patients in the acute phase. J. Clin. Monit. Comput. 2014, 28, 377–385. [Google Scholar] [CrossRef]
- Jacob, J.E.; Nair, G.K.; Iype, T.; Cherian, A. Diagnosis of encephalopathy based on energies of EEG subbands using discrete wavelet transform and support vector machine. Neurol. Res. Int. 2018, 2, 1613456. [Google Scholar] [CrossRef] [PubMed]
- Burquier, V.; Koralnik, I.J.; Vibert, D.; Burkhard, P.; Beaumanoir, A.; Jallon, P.; Mayer, E.; Hirschel, B. Effect of antiretroviral treatment on early electroencephalographic and otoneurologic manifestations in HIV infection and prognostic importance of verified perturbations. Neurophysiol. Clin. 1997, 27, 508–519. (In French) [Google Scholar] [CrossRef]
- Chen, L.; Zhu, M.; Zhou, H.; Liang, J. Clinical study of West syndrome with PS and late-onset epileptic spasms. Epilepsy. Res. 2010, 89, 82–88. [Google Scholar] [CrossRef]
- Tanoue, K.; Oguni, H.; Nakayama, N.; Sasaki, K.; Ito, Y.; Imai, K.; Osawa, M. Focal epileptic spasms, involving one leg, manifesting during the clinical course of west syndrome (WS). Brain Dev. 2008, 30, 155–159. [Google Scholar] [CrossRef]
- Beaumanoir, A.; Grioni, D.; Kullmann, G.; Tiberti, A.; Valseriati, D. EEG anomalies in the prodromic phase of Rasmussen’s syndrome. Report of two cases. Neurophysiol. Clin. 1997, 27, 25–32. (In French) [Google Scholar] [CrossRef]
- Bien, C.G.; Granata, T.; Antozzi, C.; Cross, J.H.; Dulac, O.; Kurthen, M.; Lassmann, H.; Mantegazza, R.; Villemure, J.G.; Spreafico, R.; et al. Pathogenesis, diagnosis and treatment of Rasmussen encephalitis: A European consensus statement. Brain 2005, 128, 454–471. [Google Scholar] [CrossRef]
- Capovilla, G.; Paladin, F.; Dalla Bernardina, B. Rasmussen’s syndrome: Longitudinal EEG study from the first seizure to epilepsia partialis continua. Epilepsia 1997, 38, 483–488. [Google Scholar] [CrossRef]
- Granata, T.; Gobbi, G.; Spreafico, R.; Vigevano, F.; Capovilla, G.; Ragona, F.; Freri, E.; Chiapparini, L.; Bernasconi, P.; Giordano, L.; et al. Rasmussen’s encephalitis: Early characteristics allow diagnosis. Neurology 2003, 60, 422–425. [Google Scholar] [CrossRef]
- Longaretti, F.; Dunkley, C.; Varadkar, S.; Vargha–Khadem, F.; Boyd, S.G.; Cross, J.H. Evolution of the EEG in children with Rasmussen’s syndrome. Epilepsia 2012, 53, 1539–1545. [Google Scholar] [CrossRef]
- Granata, T.; Andermann, F. Rasmussen encephalitis. Handb Clin Neurol. 2013, 111, 511–519. [Google Scholar]
- Beaumanoir, A.; Burkhard, P.; Gauthier, G.; Le Floch-Rohr, J.; Ochsner, F.; Waldvogel, F. EEG recordings in 19 cases of AIDS with encephalic involvement. Neurophysiol. Clin. 1988, 18, 313–322. (In French) [Google Scholar] [CrossRef]
- Beaumanoir, A.; Nahory, A. EEG in HIV infection. Neurophysiol. Clin. 1992, 22, 355–368. (In French) [Google Scholar] [CrossRef]
- Koralnik, I.J.; Beaumanoir, A.; Hausler, R.; Kohler, A.; Safran, A.B.; Delacoux, R.; Vibert, D.; Mayer, E.; Burkhard, P.; Nahory, A.; et al. A controlled study of early neurologic abnormalities in men with asymptomatic human immunodeficiency virus infection. N. Engl. J. Med. 1990, 323, 864–870, Erratum in 1990, 323, 1716. [Google Scholar] [CrossRef] [PubMed]
- Mesin, L.; Valerio, M.; Beaumanoir, A.; Capizzi, G. Automatic identification of Slow Biphasic Complexes in EEG: An effective tool to detect Encephalitis. Biomed. Phys. Eng. Express 2019, 5, 045006. [Google Scholar] [CrossRef]
- Mesin, L.; Valerio, M.; Capizzi, G. Detection and Assessment of Encephalitis from EEG. In Proceedings of the IEEE Conference on Computational Intelligence in Bioinformatics and Computational Biology, Siena, Italy, 9–11 July 2019. [Google Scholar]
- Valerio, M.; Rivera, S.; Mesin, L. Relation between lesions and localization of sources of slow biphasic complexes in encephalitis. Neuroimmunol. Neuroinflamm. 2020, 8, 42–49. [Google Scholar] [CrossRef]
- Mesin, L.; Valerio, M.; Capizzi, G. Automated diagnosis of encephalitis in pediatric patients using EEG rhythms and slow biphasic complexes. Phys. Eng. Sci. Med. 2020, 43, 997–1006. [Google Scholar] [CrossRef]
- Bastos, A.M.; Schoffelen, J.M. A Tutorial Review of Functional Connectivity Analysis Methods and Their Interpretational Pitfalls. Front. Syst. Neurosci. 2016, 9, 175. [Google Scholar] [CrossRef] [PubMed]
- Lehembre, R.; Marie-Aurélie, B.; Vanhaudenhuyse, A.; Chatelle, C.; Cologan, V.; Leclercq, Y.; Soddu, A.; Macq, B.; Laureys, S.; Noirhomme, Q. Resting-state EEG study of comatose patients: A connectivity and frequency analysis to find differences between vegetative and minimally conscious states. Funct. Neurol. 2012, 27, 41–47. [Google Scholar] [PubMed]
- Lachaux, J.P.; Rodriguez, E.; Martinerie, J.; Varela, F.J. Measuring phase synchrony in brain signals. Hum. Brain Mapp. 1999, 8, 194–208. [Google Scholar] [CrossRef]
- Granger, C.W.J. Investigating causal relations by econometric models and cross-spectral methods. Econometrica 1969, 37, 424–438. [Google Scholar] [CrossRef]
- Wang, H.E.; Bénar, C.G.; Quilichini, P.P.; Friston, K.J.; Jirsa, V.K.; Bernard, C. A systematic framework for functional connectivity measures. Front. Neurosci. 2014, 8, 405. [Google Scholar] [CrossRef]
- Ismail, L.E.; Karwowski, W. A Graph Theory-Based Modeling of Functional Brain Connectivity Based on EEG: A Systematic Review in the Context of Neuroergonomics. IEEE Access 2020, 8, 155103–155135. [Google Scholar] [CrossRef]
- Boashash, B.; Azem, G. A review of time–frequency matched filter design with application to seizure detection in multichannel newborn EEG. Digit. Signal Process. 2014, 28, 28–38. [Google Scholar] [CrossRef]
- Stamoulis, C.; Richardson, A.G. Application of matched filtering to identify behavioral modulation of brain oscillations. J. Comput. Neurosci. 2010, 29, 63–72. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Rubinov, M.; Sporns, O. Complex network measures of brain connectivity: Uses and interpretations. Neuroimage 2010, 52, 1059–1069. [Google Scholar] [CrossRef]
- Watts, D.; Strogatz, S. Collective dynamics of ‘small-world’ networks. Nature 1998, 393, 440–442. [Google Scholar] [CrossRef]
- Latora, V.; Marchiori, M. Efficient behavior of small-world networks. Phys. Rev. Lett. 2001, 87, 198701. [Google Scholar] [CrossRef]
- Scheirer, C.J.; Ray, W.S.; Hare, N. The analysis of ranked data derived from completely randomized factorial designs. Biometrics 1976, 32, 429–434. [Google Scholar] [CrossRef]
- Adamczyk, M.; Genzel, L.; Dresler, M.; Steiger, A.; Friess, F. Automatic sleep spindle detection and genetic influence estimation using continuous wavelet transform. Front. Hum. Neurosci. 2015, 19, 624. [Google Scholar] [CrossRef]
- Amodio, P.; Marchetti, P.; Del Piccolo, F.; de Tourtchaninoff, M.; Varghese, P.; Zuliani, C.; Campo, G.; Gatta, A.; Guérit, J.M. Spectral versus visual EEG analysis in mild hepatic encephalopathy. Clin. Neurophysiol. 1999, 110, 1334–1344. [Google Scholar] [CrossRef]
- Azuma, H.; Hori, S.; Nakanishi, M.; Fujimoto, S.; Ichikawa, N.; Furukawa, T.A. An intervention to improve the interrater reliability of clinical EEG interpretations. Psychiatry Clin. Neurosci. 2003, 57, 485–489. [Google Scholar] [CrossRef]
- Ji, Z.; Sugi, T.; Goto, S.; Wang, X.; Ikeda, A.; Nagamine, T.; Shibasaki, H.; Nakamura, M. An automatic spike detection system based on elimination of false positives using the large–area context in the scalp EEG. IEEE Trans. Biomed. Eng. 2011, 58, 2478–2488. [Google Scholar]
- Mohammad, S.S.; Soe, S.M.; Pillai, S.C.; Nosadini, M.; Barnes, E.H.; Gill, D.; Dale, R.C. Etiological associations and outcome predictors of acute electroencephalography in childhood encephalitis. Clin. Neurophysiol. 2016, 127, 3217–3224. [Google Scholar] [CrossRef]
- Scheuer, M.L.; Bagic, A.; Wilson, S.B. Spike detection: Inter–reader agreement and a statistical Turing test on a large data set. Clin. Neurophysiol. 2017, 128, 243–250. [Google Scholar] [CrossRef]
- Stevenson, N.J.; Korotchikova, I.; Temko, A.; Lightbody, G.; Marnane, W.P.; Boylan, G.B. An automated system for grading EEG abnormality in term neonates with hypoxic-ischaemic encephalopathy. Ann. Biomed. Eng. 2013, 41, 775–785. [Google Scholar] [CrossRef]
- Stroink, H.; Schimsheimer, R.J.; de Weerd, A.W.; Geerts, A.T.; Arts, W.F.; Peeters, E.A.; Brouwer, O.F.; Boudewijn Peters, A.; van Donselaar, C.A. Interobserver reliability of visual interpretation of electroencephalograms in children with newly diagnosed seizures. Dev. Med. Child. Neurol. 2006, 48, 374–377. [Google Scholar] [CrossRef]
- Cooray, G.K.; Sengupta, B.; Douglas, P.; Englund, M.; Wickstrom, R.; Friston, K. Characterising seizures in anti–NMDA–receptor encephalitis with dynamic causal modelling. Neuroimage 2015, 118, 508–519. [Google Scholar] [CrossRef][Green Version]
- Dobleand, M.; Narayan, S.K. Mathematical analysis of EEG of patients with non–fatal nonspecific diffuse encephalitis. Int. J. Biol. Med. Sci. 2008, 3, 4. [Google Scholar]
- Beaumanoir, A.; Magistris, M.; Nahory, A. Sporadic slow biphasic complex: Description and clinical correlations. Electroenceph. Clin. Neurophysiol. 1985, 61, S142. [Google Scholar] [CrossRef]
- Bassett, D.S.; Bullmore, E.T. Small-World Brain Networks Revisited. Neuroscientist 2017, 23, 499–516. [Google Scholar] [CrossRef]
- Liao, X.; Vasilakos, A.V.; He, Y. Small-world human brain networks: Perspectives and challenges. Neurosci. Biobehav. Rev. 2017, 77, 286–300. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiarion, G.; Mesin, L. Functional Connectivity of EEG in Encephalitis during Slow Biphasic Complexes. Electronics 2021, 10, 2978. https://doi.org/10.3390/electronics10232978
Chiarion G, Mesin L. Functional Connectivity of EEG in Encephalitis during Slow Biphasic Complexes. Electronics. 2021; 10(23):2978. https://doi.org/10.3390/electronics10232978
Chicago/Turabian StyleChiarion, Giovanni, and Luca Mesin. 2021. "Functional Connectivity of EEG in Encephalitis during Slow Biphasic Complexes" Electronics 10, no. 23: 2978. https://doi.org/10.3390/electronics10232978
APA StyleChiarion, G., & Mesin, L. (2021). Functional Connectivity of EEG in Encephalitis during Slow Biphasic Complexes. Electronics, 10(23), 2978. https://doi.org/10.3390/electronics10232978