
Divergent Effects of Topography on Soil Properties and Above-Ground Biomass in Nepal’s Mid-Hill Forests



Abstract
:1. Introduction
2. Materials and Methods
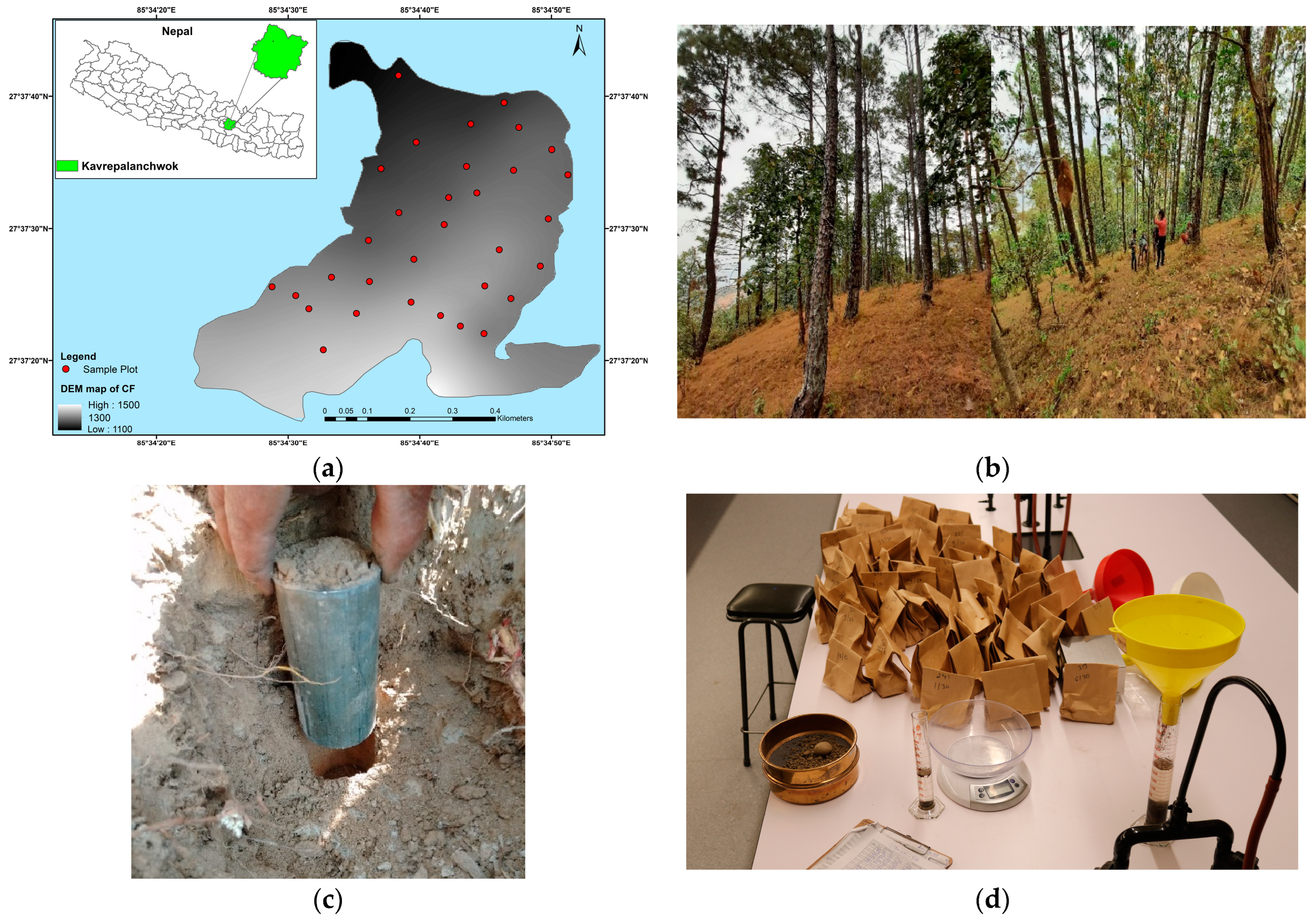
2.1. Study Area
2.2. Design of the Experiment
2.3. Soil Laboratory Analysis
2.4. Above-Ground Vegetation Analysis
2.5. Statistical Analysis
3. Results
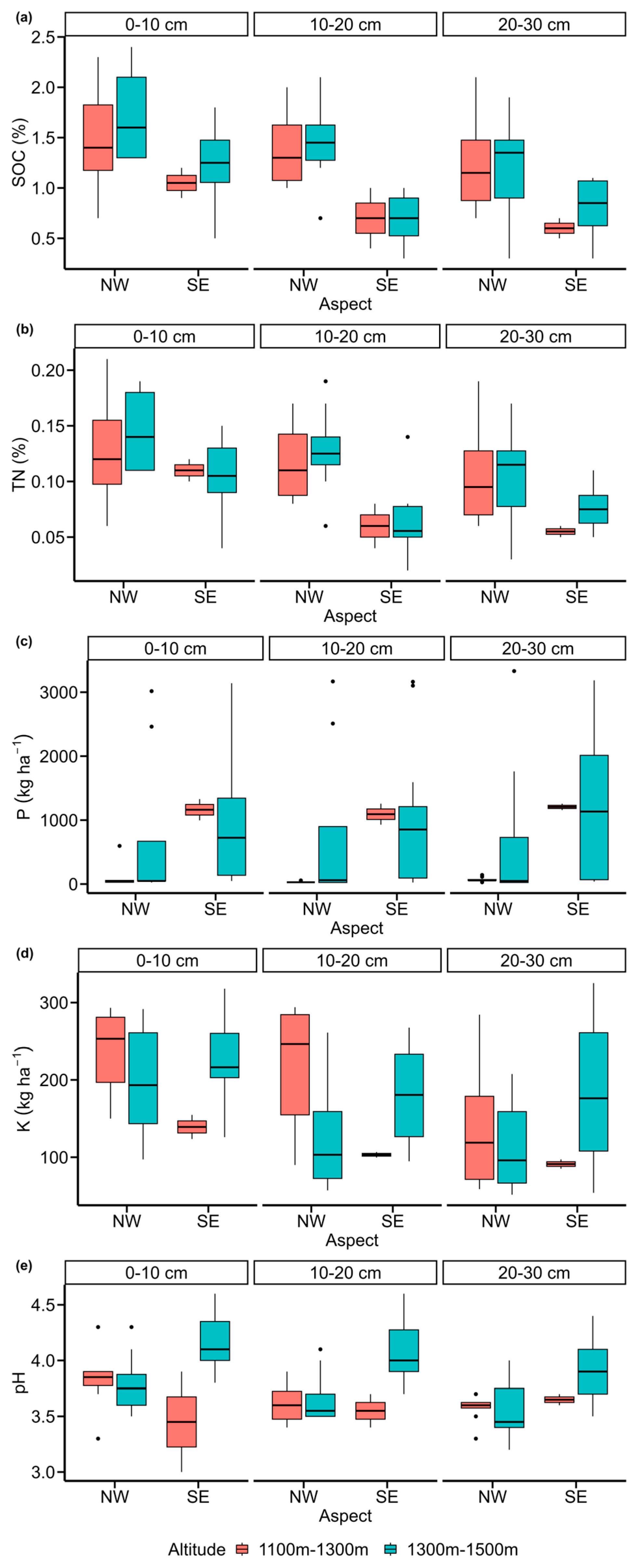
3.1. Effects of Aspect, Altitude, and Soil Depth on Soil Parameters
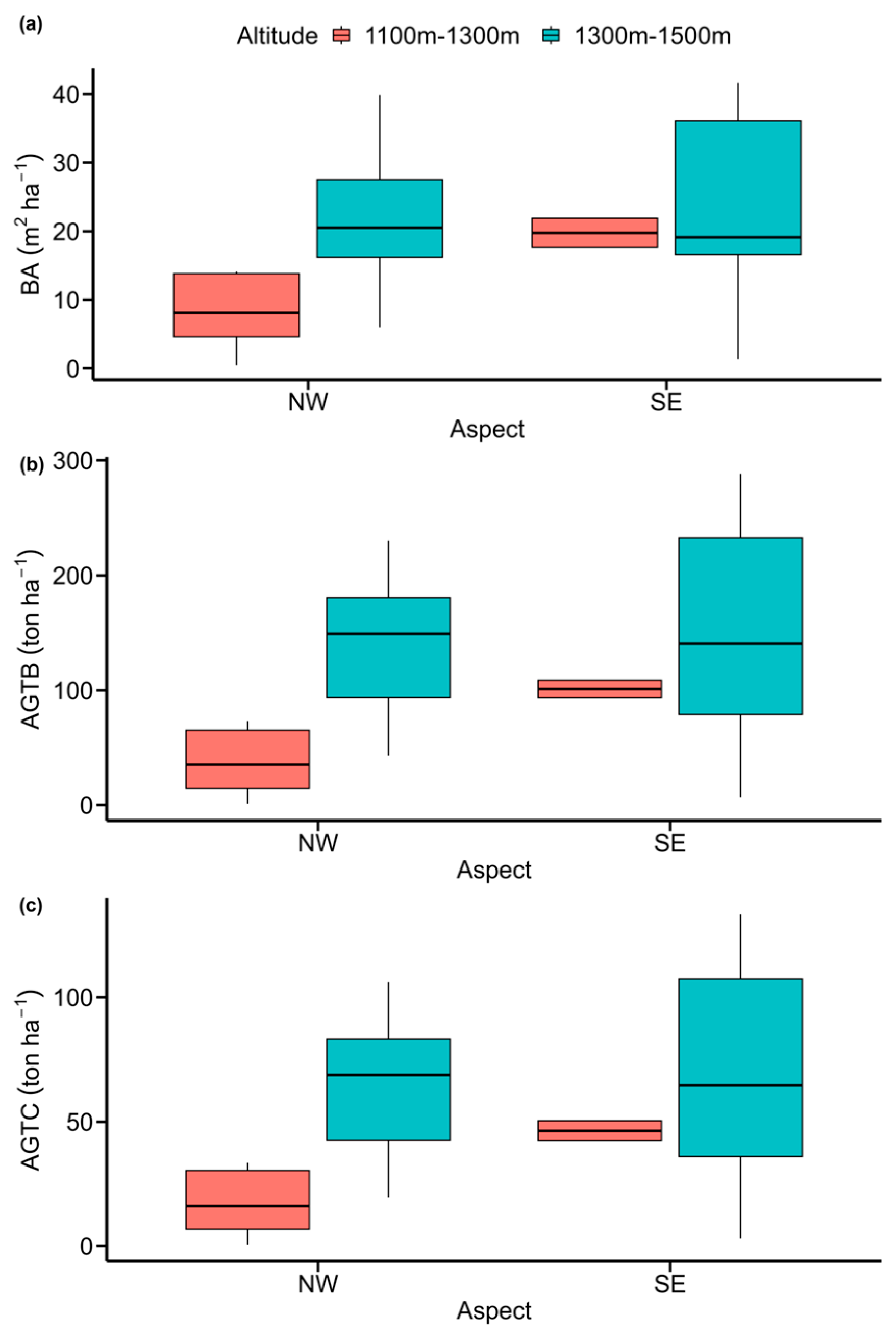
3.2. Effects of Aspect, Altitude, and Species Type on Vegetation Parameters
3.3. Relationships between Above-Ground Variables and Soil Nutrients
4. Discussion
4.1. Effects of Aspect, Altitude, and Soil Depth on Soil Chemical Properties
4.2. Effects of Aspect, Altitude, and Species Type on Vegetation Parameters
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Gairola, S.; Sharma, C.M.; Ghildiyal, S.K.; Suyal, S. Chemical Properties of Soils in Relation to Forest Composition in Moist Temperate Valley Slopes of Garhwal Himalaya, India. Environmentalist 2012, 32, 512–523. [Google Scholar] [CrossRef]
- Lal, R.; Kimble, J.; Follett, R. Land Use and Soil C Pools in Terrestrial Ecosystems. In Management of Carbon Sequestration in Soil; CRC Press: Boca Raton, FL, USA, 1998; pp. 1–10. [Google Scholar]
- Luintel, H.; Bluffstone, R.A.; Scheller, R.M. The Effects of the Nepal Community Forestry Program on Biodiversity Conservation and Carbon Storage. PLoS ONE 2018, 13, e0199526. [Google Scholar] [CrossRef]
- Dearborn, K.D.; Danby, R.K. Aspect and Slope Influence Plant Community Composition More than Elevation across Forest–Tundra Ecotones in Subarctic Canada. J. Veg. Sci. 2017, 28, 595–604. [Google Scholar] [CrossRef]
- Román-Sánchez, A.; Vanwalleghem, T.; Peña, A.; Laguna, A.; Giráldez, J.V. Controls on Soil Carbon Storage from Topography and Vegetation in a Rocky, Semi-Arid Landscapes. Geoderma 2018, 311, 159–166. [Google Scholar] [CrossRef]
- Sanaei, A.; Chahouki, M.A.Z.; Ali, A.; Jafari, M.; Azarnivand, H. Abiotic and Biotic Drivers of Aboveground Biomass in Semi-Steppe Rangelands. Sci. Total Environ. 2018, 615, 895–905. [Google Scholar] [CrossRef] [PubMed]
- Lybrand, R.A.; Rasmussen, C. Quantifying Climate and Landscape Position Controls on Soil Development in Semiarid Ecosystems. Soil Sci. Soc. Am. J. 2015, 79, 104–116. [Google Scholar] [CrossRef]
- Schwanghart, W.; Jarmer, T. Linking Spatial Patterns of Soil Organic Carbon to Topography—A Case Study from South-Eastern Spain. Geomorphology 2011, 126, 252–263. [Google Scholar] [CrossRef]
- Müller, M.; Schickhoff, U.; Scholten, R.P.; Drollinger, S.; Böhner, J.; Chaudhary, R.P. How Do Soil Properties Affect Alpine Treelines? General Principles in a Global Perspective and Novel Findings from Rolwaling Himal, Nepal. Prog. Phys. Geogr. Earth Environ. 2016, 40, 135–160. [Google Scholar] [CrossRef]
- Bajaracharya, R. Land Preparation: An Integral Part of Farming Systems in the Mid-Hills of Nepal. Nepal J. Sci. Technol. 2001, 3, 15–24. [Google Scholar]
- Post, W.M.; Kwon, K.C. Soil Carbon Sequestration and Land-Use Change: Processes and Potential. Glob. Chang. Biol. 2000, 6, 317–327. [Google Scholar] [CrossRef]
- Schreier; Shrestha, B.; Brown, S.; Shah, P.B. Challenges in Mountain Resource Management in Nepal: Processes, Trends, and Dynamics in Middle Mountain Watershed Kathmandu, Nepal; International Centre for Integrated Mountain Development: Kathmandu, Nepal, 1999. [Google Scholar]
- Fontaine, S.; Barot, S.; Barré, P.; Bdioui, N.; Mary, B.; Rumpel, C. Stability of Organic Carbon in Deep Soil Layers Controlled by Fresh Carbon Supply. Nature 2007, 450, 277–280. [Google Scholar] [CrossRef] [PubMed]
- Kewlani, P.; Negi, V.S.; Bhatt, I.D.; Rawal, R.S.; Nandi, S.K. Soil Nutrients Concentration along Altitudinal Gradients in Indian Western Himalaya. Scand. J. For. Res. 2021, 36, 98–104. [Google Scholar] [CrossRef]
- Shedayi, A.A.; Xu, M.; Naseer, I.; Khan, B. Altitudinal Gradients of Soil and Vegetation Carbon and Nitrogen in a High Altitude Nature Reserve of Karakoram Ranges. Springerplus 2016, 5, 320. [Google Scholar] [CrossRef]
- Matus, F.; Rumpel, C.; Neculman, R.; Panichini, M.; Mora, M.L. Soil Carbon Storage and Stabilisation in Andic Soils: A Review. Catena 2014, 120, 102–110. [Google Scholar] [CrossRef]
- Cao, J.; Wang, X.; Tian, Y.; Wen, Z.; Zha, T. Pattern of Carbon Allocation across Three Different Stages of Stand Development of a Chinese Pine (Pinus tabulaeformis) Forest. Ecol. Res. 2012, 27, 883–892. [Google Scholar] [CrossRef]
- Kumar, A.; Sharma, M.P. Estimation of Carbon Stocks of Balganga Reserved Forest, Uttarakhand, India. For. Sci. Technol. 2015, 11, 177–181. [Google Scholar] [CrossRef]
- Leuschner, C.; Moser, G.; Bertsch, C.; Röderstein, M.; Hertel, D. Large Altitudinal Increase in Tree Root/Shoot Ratio in Tropical Mountain Forests of Ecuador. Basic Appl. Ecol. 2007, 8, 219–230. [Google Scholar] [CrossRef]
- Mekonnen, M.; Keesstra, S.D.; Stroosnijder, L.; Baartman, J.E.M.; Maroulis, J. Soil Conservation through Sediment Trapping: A Review. Land Degrad. Dev. 2015, 26, 544–556. [Google Scholar] [CrossRef]
- Wang, B.; Xue, S.; Liu, G.B.; Zhang, G.H.; Li, G.; Ren, Z.P. Changes in Soil Nutrient and Enzyme Activities under Different Vegetations in the Loess Plateau Area, Northwest China. Catena 2012, 92, 186–195. [Google Scholar] [CrossRef]
- Carroll, J.A.; Caporn, S.J.M.; Johnson, D.; Morecroft, M.D.; Lee, J.A. The Interactions between Plant Growth, Vegetation Structure and Soil Processes in Semi-Natural Acidic and Calcareous Grasslands Receiving Long-Term Inputs of Simulated Pollutant Nitrogen Deposition. Environ. Pollut. 2003, 121, 363–376. [Google Scholar] [CrossRef]
- Ceacero, C.J.; Díaz-Hernández, J.L.; del Campo, A.D.; Navarro-Cerrillo, R.M. Interactions between Soil Gravel Content and Neighboring Vegetation Control Management in Oak Seedling Establishment Success in Mediterranean Environments. For. Ecol. Manag. 2012, 271, 10–18. [Google Scholar] [CrossRef]
- Naeth, M.A.; Chanasyk, D.S.; Burgers, T.D. Vegetation and Soil Water Interactions on a Tailings Sand Storage Facility in the Athabasca Oil Sands Region of Alberta Canada. Phys. Chem. Earth 2011, 36, 19–30. [Google Scholar] [CrossRef]
- Benbi, D.K.; Chand, M. Quantifying the Effect of Soil Organic Matter on Indigenous Soil N Supply and Wheat Productivity in Semiarid Sub-Tropical India. Nutr. Cycl. Agroecosyst. 2007, 79, 103–112. [Google Scholar] [CrossRef]
- Liao, J.D.; Boutton, T.W.; Jastrow, J.D. Storage and Dynamics of Carbon and Nitrogen in Soil Physical Fractions following Woody Plant Invasion of Grassland. Soil Biol. Biochem. 2006, 38, 3184–3196. [Google Scholar] [CrossRef]
- Xia, Y.Q.; Shao, M.A. Soil Water Carrying Capacity for Vegetation: A Hydrologic and Biogeochemical Process Model Solution. Ecol. Model. 2008, 214, 112–124. [Google Scholar] [CrossRef]
- Méndez-Toribio, M.; Ibarra-Manríquez, G.; Navarrete-Segueda, A.; Paz, H. Topographic Position, but Not Slope Aspect, Drives the Dominance of Functional Strategies of Tropical Dry Forest Trees. Environ. Res. Lett. 2017, 12, 85002. [Google Scholar] [CrossRef]
- Bajracharya, R.M.; Sitaula, B.; Shrestha, B.M.; Awasthi, K.D. Soil Organic Carbon Status and Dynamics in the Central Nepal Middle Mountains. Forestry 2004, 12, 28–44. [Google Scholar]
- Begum, F.; Bajracharya, R.M.; Sharma, S.; Sitaula, B.K. Influence of Slope Aspect on Soil Physico-Chemical and Biological Properties in the Mid Hills of Central Nepal. Int. J. Sustain. Dev. World Ecol. 2010, 17, 438–443. [Google Scholar] [CrossRef]
- Department of Forest and Soil Conservation. Forest Management in Nepal; Department of Forest and Soil Conservation: Kathmandu, Nepal, 2020.
- MPFS. MPFS: Master Plan for the Forestry Sector, Nepal; Government of Nepal: Kathmandu, Nepal, 1989.
- Department of Hydrology and Meteorology. Observed Climate Trend Analysis of Nepal (1971–2014); Department of Hydrology and Meteorology: Kathmandu, Nepal, 2017.
- Division Forest Office. Annual Progress Report for the Fiscal Year 2019–20; Division Forest Office: Kavre, Nepal, 2020.
- Bremner, J.M. Total Nitrogen, Methods of Soil Analysis: Part 2 Chemical and Microbiological Properties; ASA: St. Louis, MO, USA, 1965; pp. 1149–1178. [Google Scholar]
- Page, A.L.; Miller, R.H.; Keeney, D.R. Methods of Soil Analysis. Part 2. American Society of Agronomy. Soil Sci. Soc. Am. J. 1982, 4, 167–179. [Google Scholar]
- Olsen, S.R. Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate; US Department of Agriculture: Washington, DC, USA, 1954; p. 939.
- Morwin, H.D.; Peach, P.M. Exchangeability of Soil Potassium in and, Silt and Clay Fractions as Influenced by the Nature of Complementary Exchangeable Cations. Soil Sci. Soc. Am. J. 1951, 15, 125–128. [Google Scholar] [CrossRef]
- Sharma, E.R.; Pukkala, T. Volume Equations and Biomass Prediction of Forest Trees in Nepal; Ministry of Forests and Soil Conservation, Forest Survey and Statistics Division: Kathmandu, Nepal, 1990.
- Jackson, J.K. Manual of Afforestation in Nepal; Forest Research and Survey Centre: Kathmandu, Nepal, 1994.
- Negi, J.D.S.; Manhas, R.K.; Chauhan, P.S. Carbon Allocation in Different Components of Some Tree Species of India: A New Approach for Carbon Estimation. Curr. Sci. 2003, 85.11, 1528–1531. [Google Scholar]
- MacDicken, K.G. A Guide to Monitoring Carbon Storage in Forestry and Agroforestry Projects; Winrock International Institute of Agricultural Development, Forest Carbon Monitoring Program: Arlington, VA, USA, 1997. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; R Core Team. Linear and Nonlinear Mixed Effects Models (R Package Version 3.1-137); R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: http://cran.r-project.org/web/packages/nlme (accessed on 7 November 2023).
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Springer: New York, NY, USA, 2009; Volume 574, p. 574. [Google Scholar]
- Bangroo, S.A.; Najar, G.R.; Rasool, A. Effect of Altitude and Aspect on Soil Organic Carbon and Nitrogen Stocks in the Himalayan Mawer Forest Range. Catena 2017, 158, 63–68. [Google Scholar] [CrossRef]
- Pang, X.Y.; Bao, W.K.; Zang, Y.M. Evaluation of Soil Fertility under Different Cupressus Chengiana Forests Using Multivariate Approach. Pedosphere 2006, 16, 602–615. [Google Scholar] [CrossRef]
- Måren, I.E.; Karki, S.; Prajapati, C.; Yadav, R.K.; Shrestha, B.B. Facing North or South: Does Slope Aspect Impact Forest Stand Characteristics and Soil Properties in a Semiarid Trans-Himalayan Valley? J. Arid Environ. 2015, 121, 112–123. [Google Scholar] [CrossRef]
- Smith, J.L.; Halvorson, J.J.; Bolton, H. Soil Properties and Microbial Activity across a 500 m Elevation Gradient in a Semi-Arid Environment. Soil Biol. Biochem. 2002, 34, 1749–1757. [Google Scholar] [CrossRef]
- Zhang, S.; Chen, D.; Sun, D.; Wang, X.; Smith, J.L.; Du, G. Impacts of Altitude and Position on the Rates of Soil Nitrogen Mineralization and Nitrification in Alpine Meadows on the Eastern Qinghai-Tibetan Plateau, China. Biol. Fertil. Soils 2012, 48, 393–400. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, X.; Huffman, T.; Liu, X.; Yang, Y. Influence of Topography and Land Management on Soil Nutrients Variability in Northeast China. Nutr. Cycl. Agroecosyst. 2011, 89, 427–438. [Google Scholar] [CrossRef]
- Kato, T.; Tang, Y.; Gu, S.; Hirota, M.; Du, M.; Li, Y.; Zhao, X. Temperature and Biomass Influences on Interannual Changes in CO2 Exchange in an Alpine Meadow on the Qinghai-Tibetan Plateau. Glob. Chang. Biol. 2006, 12, 1285–1298. [Google Scholar] [CrossRef]
- Tripathi, B. Review of Acid Soil and Its Management in Nepal. In Lumle Seminar Paper; Agricultural Research Station: Lumle, Nepal, 1999; Volume 99, pp. 1–11. [Google Scholar]
- Paul, E.A. Soil Microbiology, Ecology, and Biochemistry in Perspective. In Soil Microbiology, Ecology and Biochemistry, 3rd ed.; Academic Press: Cambridge, MA, USA, 2007; pp. 3–24. [Google Scholar]
- Yebo, B. Integrated Soil Fertility Management for Better Crop Production in Ethiopia. Int. J. Soil Sci. 2015, 10, 1–16. [Google Scholar] [CrossRef]
- Dahlgren, R.A.; Boettinger, J.L.; Huntington, G.L.; Amundson, R.G. Soil Development along an Elevational Transect in the Western Sierra Nevada, California. Geoderma 1997, 78, 207–236. [Google Scholar] [CrossRef]
- Bhattacharyya, T.; Pal, D.K.; Chandran, P.; Ray, S.K.; Mandal, C.; Telpande, B. Soil Carbon Storage Capacity as a Tool to Prioritize Areas for Carbon Sequestration. Curr. Sci. 2008, 95, 482–494. [Google Scholar]
- Müller, M.; Oelmann, Y.; Schickhoff, U.; Böhner, J.; Scholten, T. Himalayan Treeline Soil and Foliar C:N:P Stoichiometry Indicate Nutrient Shortage with Elevation. Geoderma 2017, 291, 21–32. [Google Scholar] [CrossRef]
- Strong, C.L.; Boulter, S.L.; Laidlaw, M.J.; Maunsell, S.C.; Putland, D.; Kitching, R.L. The Physical Environment of an Altitudinal Gradient in the Rainforest of Lamington National Park, Southeast Queensland. Mem. Qld. Mus. 2011, 55, 251–270. [Google Scholar]
- Vieira, S.A.; Alves, L.F.; Duarte-Neto, P.J.; Martins, S.C.; Veiga, L.G.; Scaranello, M.A.; Picollo, M.C.; Camargo, P.B.; do Carmo, J.B.; Neto, E.S.; et al. Stocks of Carbon and Nitrogen and Partitioning between Above-and Belowground Pools in the Brazilian Coastal Atlantic Forest Elevation Range. Ecol. Evol. 2011, 1, 421–434. [Google Scholar] [CrossRef]
- Kou, D.; Ma, W.; Ding, J.; Zhang, B.; Fang, K.; Hu, H.; Yu, J.; Wang, T.; Qin, S.; Zhao, X.; et al. Dryland Soils in Northern China Sequester Carbon during the Early 2000s Warming Hiatus Period. Funct. Ecol. 2018, 32, 1620–1630. [Google Scholar] [CrossRef]
- García, E.; Diéguez-Aranda, U.; Cunha, M.; Rodríguez-Soalleiro, R. Comparison of Harvest-Related Removal of Aboveground Biomass, Carbon and Nutrients in Pedunculate Oak Stands and in Fast-Growing Tree Stands in NW Spain. For. Ecol. Manag. 2016, 365, 119–127. [Google Scholar] [CrossRef]
- Sharma, C.M.; Baduni, N.P.; Gairola, S.; Ghildiyal, S.K.; Suyal, S. Effects of Slope Aspects on Forest Compositions, Community Structures and Soil Properties in Natural Temperate Forests of Garhwal Himalaya. J. For. Res. 2010, 21, 331–337. [Google Scholar] [CrossRef]
- Mohan, K.C.; Mason, E.G.; Bown, H.E. Linking above-Ground Biomass Production to below-Ground Carbon Fluxes across Stocking, Clone, Fertilization, and Understory Elimination in Pinus Radiata D.Don Plantations, New Zealand. For. Ecol. Manag. 2020, 477, 118469. [Google Scholar]
- Motsinger, J.R.; Kabrick, J.M.; Dey, D.C.; Henderson, D.E.; Zenner, E.K. Effect of Midstory and Understory Removal on the Establishment and Development of Natural and Artificial Pin Oak Advance Reproduction in Bottomland Forests. New For. 2010, 39, 195–213. [Google Scholar] [CrossRef]
- Saremi, H.; Kumar, L.; Turner, R.; Stone, C.; Melville, G. Impact of Local Slope and Aspect Assessed from LiDAR Records on Tree Diameter in Radiata Pine (Pinus Radiata D. Don) Plantations. Ann. For. Sci. 2014, 71, 771–780. [Google Scholar] [CrossRef]
- Auslander, M.; Nevo, E.; Inbar, M. The Effects of Slope Orientation on Plant Growth, Developmental Instability and Susceptibility to Herbivores. J. Arid Environ. 2003, 55, 405–416. [Google Scholar] [CrossRef]
- Rohner, B.; Bugmann, H.; Bigler, C. Estimating the Age-Diameter Relationship of Oak Species in Switzerland Using Nonlinear Mixed-Effects Models. Eur. J. For. Res. 2013, 132, 751–764. [Google Scholar] [CrossRef]
- King, D.A. The Adaptive Significance of Tree Height. Am. Nat. 1990, 135, 809–828. [Google Scholar] [CrossRef]
- Koch, G.W.; Stillet, S.C.; Jennings, G.M.; Davis, S.D. The Limits to Tree Height. Nature 2004, 428, 851–854. [Google Scholar] [CrossRef]
- Domec, J.C.; Lachenbruch, B.; Meinzer, F.C.; Woodruff, D.R.; Warren, J.M.; McCulloh, K.A. Maximum Height in a Conifer Is Associated with Conflicting Requirements for Xylem Design. Proc. Natl. Acad. Sci. USA 2008, 105, 12069–12074. [Google Scholar] [CrossRef]
- Petit, G.; Anfodillo, T.; Carraro, V.; Grani, F.; Carrer, M. Hydraulic Constraints Limit Height Growth in Trees at High Altitude. New Phytol. 2011, 189, 241–252. [Google Scholar] [CrossRef]
- Thokchom, A.; Yadava, P.S. Biomass and Carbon Stock along an Altitudinal Gradient in the Forest of Manipur, Northeast India. Trop. Ecol. 2017, 58, 389–396. [Google Scholar]
- Peichl, M.; Arain, M.A. Above- and Belowground Ecosystem Biomass and Carbon Pools in an Age-Sequence of Temperate Pine Plantation Forests. Agric. For. Meteorol. 2006, 140, 51–63. [Google Scholar] [CrossRef]
- Bhandari, J.; Zhang, Y. Effect of Altitude and Soil Properties on Biomass and Plant Richness in the Grasslands of Tibet, China, and Manang District, Nepal. Ecosphere 2019, 10, e02915. [Google Scholar] [CrossRef]
- Riihimäki, H.; Heiskanen, J.; Luoto, M. The Effect of Topography on Arctic-Alpine Aboveground Biomass and NDVI Patterns. Int. J. Appl. Earth Obs. Geoinf. 2017, 56, 44–53. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Min | Max | Mean | SE |
|---|---|---|---|---|
| Soil organic carbon (%) | 0.30 | 2.40 | 1.16 | 0.05 |
| Total nitrogen (%) | 0.02 | 0.21 | 0.10 | 0.00 |
| Total phosphorus (kg ha−1) | 24.10 | 3330.30 | 727.00 | 100.34 |
| Potassium (kg ha−1) | 51.60 | 325.20 | 178.46 | 8.04 |
| Soil pH | 3.00 | 4.60 | 3.83 | 0.03 |
| Diameter at breast height (cm) | 5.25 | 29.83 | 21.45 | 0.60 |
| Tree height (m) | 4.50 | 17.67 | 12.38 | 0.35 |
| Tree basal area (m2 ha−1) | 0.43 | 41.68 | 18.63 | 1.19 |
| Above-ground total tree biomass (ton ha−1) | 0.99 | 288.52 | 116.27 | 8.28 |
| Above-ground total tree carbon (ton ha−1) | 0.45 | 133.29 | 53.54 | 3.83 |
| Factors | df | Soil Parameters | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| SOC | TN | P | K | pH | |||||||
| F | p | F | p | F | p | F | p | F | p | ||
| Aspect | 1,28 | 22.79 | 0.0001 | 16.93 | 0.0003 | 4.20 | 0.0499 | 16.43 | 0.0004 | ||
| Altitude | 1,28 | ||||||||||
| Depth | 2,56 | 18.66 | <0.0001 | 16.61 | <0.0001 | 17.01 | <0.0001 | 12.13 | <0.0001 | ||
| Species | 3,28 | ||||||||||
| Factors | df | Vegetation Parameters | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| DBH | Ht | BA | AGTB | AGTC | |||||||
| F | p | F | p | F | p | F | p | F | p | ||
| Aspect | 1,28 | 13.2962 | 0.0011 | 16.6391 | 0.0003 | 4.51651 | 0.0425 | 4.49901 | 0.0429 | ||
| Altitude | 1,28 | 8.8149 | 0.0061 | 13.4725 | 0.0010 | 5.94251 | 0.0214 | 8.84873 | 0.0060 | 8.78523 | 0.0061 |
| Depth | 2,56 | ||||||||||
| Species | 3,28 | 16.0605 | <0.0001 | 18.5343 | <0.0001 | 21.89512 | <0.0001 | 21.44612 | <0.0001 | 21.58518 | <0.0001 |
| Category | Levels | Soil Parameters | ||||
|---|---|---|---|---|---|---|
| SOC | TN | P | K | pH | ||
| Aspect | SE | 0.85 ± 0.11 a | 0.08 ± 0.01 a | 1095 ± 338 a | 154 ± 21.7 a | 3.80 ± 0.08 a |
| NW | 1.42 ± 0.08 b | 0.12 ± 0.01 b | 403 ± 223 b | 171 ± 14.3 a | 3.67 ± 0.05 a | |
| Altitude | A1 | 1.08 ± 0.12 a | 0.09 ± 0.01 a | 612 ± 353 a | 154 ± 22.6 a | 3.61 ± 0.09 a |
| A2 | 1.20 ± 0.07 a | 0.10 ± 0.01 a | 886 ± 198 a | 171 ± 12.7 a | 3.86 ± 0.05 b | |
| Soil depth (cm) | D1 | 1.38 ± 0.09 b | 0.12 ± 0.01 b | 732 ± 215 a | 201 ± 15.9 b | 3.81 ± 0.06 a |
| D2 | 1.06 ± 0.09 a | 0.09 ± 0.01 a | 716 ± 215 a | 157 ± 15.9 a | 3.73 ± 0.06 a | |
| D3 | 0.97 ± 0.09 a | 0.08 ± 0.01 a | 800 ± 215 a | 130 ± 15.9 a | 3.68 ± 0.06 a | |
| Category | Levels | Vegetation Parameters | ||||
|---|---|---|---|---|---|---|
| DBH | Ht | BA | AGTB | AGTC | ||
| Aspect | SE | 23.60 ± 1.73 b | 13.10 ± 0.94 b | 21.00 ± 3.98 a | 124.90 ± 26.60 a | 57.50 ± 12.30 a |
| NW | 18.50 ± 1.17 a | 10.60 ± 0.63 a | 15.30 ± 2.63 a | 89.80 ± 17.60 a | 41.30 ± 8.14 a | |
| Altitude | A1 | 18.70 ± 1.81 a | 10.10 ± 0.99 a | 14.00 ± 4.17 a | 69.30 ± 27.80 a | 31.80 ± 12.86 a |
| A2 | 23.40 ± 1.02 b | 13.60 ± 0.55 b | 22.40 ± 2.33 a | 145.00 ± 15.60 b | 67.00 ± 7.21 b | |
| Species | Pinus wallichiana | 27.00 ± 1.41 b | 15.21 ± 0.71 c | 19.86 ± 1.62 b | 129.87 ± 11.10 b | 59.97 ± 5.13 b |
| Schima Wallichii | 16.30 ± 1.35 a | 9.67 ± 0.68 b | 3.91 ± 1.55 a | 22.19 ± 10.70 a | 9.99 ± 4.92 a | |
| Alnus nepalensis | 17.20 ± 2.38 a | 10.52 ± 1.20 b | 4.13 ± 2.75 a | 17.58 ± 18.90 a | 8.26 ± 8.70 a | |
| Gravellia robusta | 10.20 ± 2.55 a | 5.64 ± 1.29 a | 1.24 ± 2.93 a | 5.47 ± 20.20 a | 2.57 ± 9.30 a | |
| Variables | SOC | TN | P | K | pH | DBH | Ht | BA | AGTB | AGTC |
|---|---|---|---|---|---|---|---|---|---|---|
| SOC | 1 | 0.95 *** | −0.07 | 0.25 * | 0.03 | −0.29 ** | −0.39 *** | −0.11 | −0.15 | −0.16 |
| TN | 1 | −0.08 | 0.21 * | 0.00 | −0.27 ** | −0.37 *** | −0.03 | −0.09 | −0.09 | |
| P | 1 | 0.17 | 0.18 | 0.38 *** | 0.28 ** | 0.40 *** | 0.37 *** | 0.36 *** | ||
| K | 1 | 0.26 * | 0.13 | 0.06 | 0.21 * | 0.19 | 0.19 | |||
| pH | 1 | 0.21 * | 0.23 * | 0.02 | 0.05 | 0.05 | ||||
| DBH | 1 | 0.82 *** | 0.73 *** | 0.71 *** | 0.71 *** | |||||
| Ht | 1 | 0.66 *** | 0.75 *** | 0.75 *** | ||||||
| BA | 1 | 0.97 *** | 0.97 *** | |||||||
| AGTB | 1 | 1.00 *** | ||||||||
| AGTC | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nepal, S.; KC, M.; Pudasaini, N.; Adhikari, H. Divergent Effects of Topography on Soil Properties and Above-Ground Biomass in Nepal’s Mid-Hill Forests. Resources 2023, 12, 136. https://doi.org/10.3390/resources12110136
Nepal S, KC M, Pudasaini N, Adhikari H. Divergent Effects of Topography on Soil Properties and Above-Ground Biomass in Nepal’s Mid-Hill Forests. Resources. 2023; 12(11):136. https://doi.org/10.3390/resources12110136
Chicago/Turabian StyleNepal, Sandhya, Mohan KC, Nabaraj Pudasaini, and Hari Adhikari. 2023. "Divergent Effects of Topography on Soil Properties and Above-Ground Biomass in Nepal’s Mid-Hill Forests" Resources 12, no. 11: 136. https://doi.org/10.3390/resources12110136
APA StyleNepal, S., KC, M., Pudasaini, N., & Adhikari, H. (2023). Divergent Effects of Topography on Soil Properties and Above-Ground Biomass in Nepal’s Mid-Hill Forests. Resources, 12(11), 136. https://doi.org/10.3390/resources12110136