Increased Plasma Levels of Adenylate Cyclase 8 and cAMP Are Associated with Obesity and Type 2 Diabetes: Results from a Cross-Sectional Study

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Demographic Data of Study Population
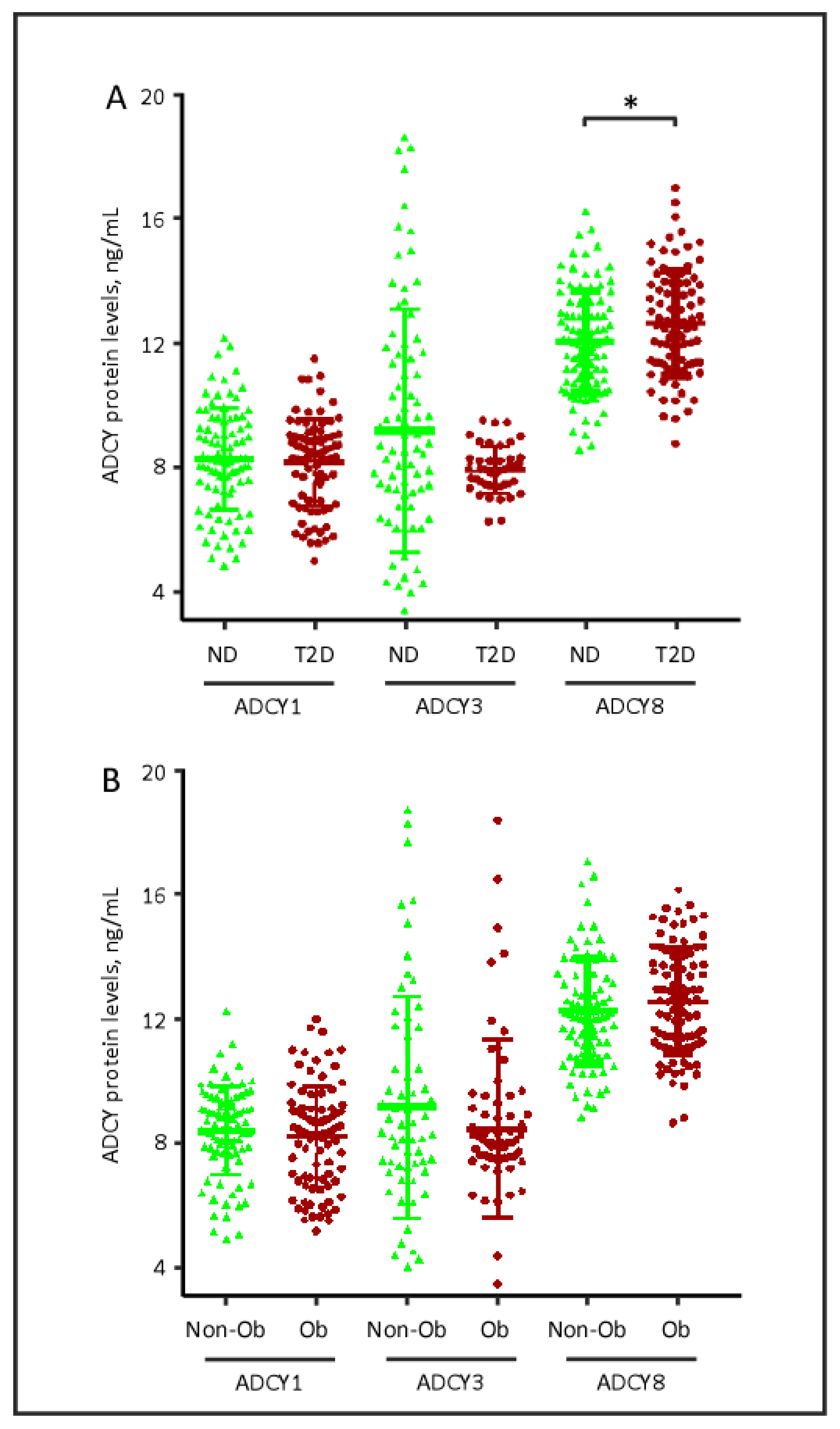
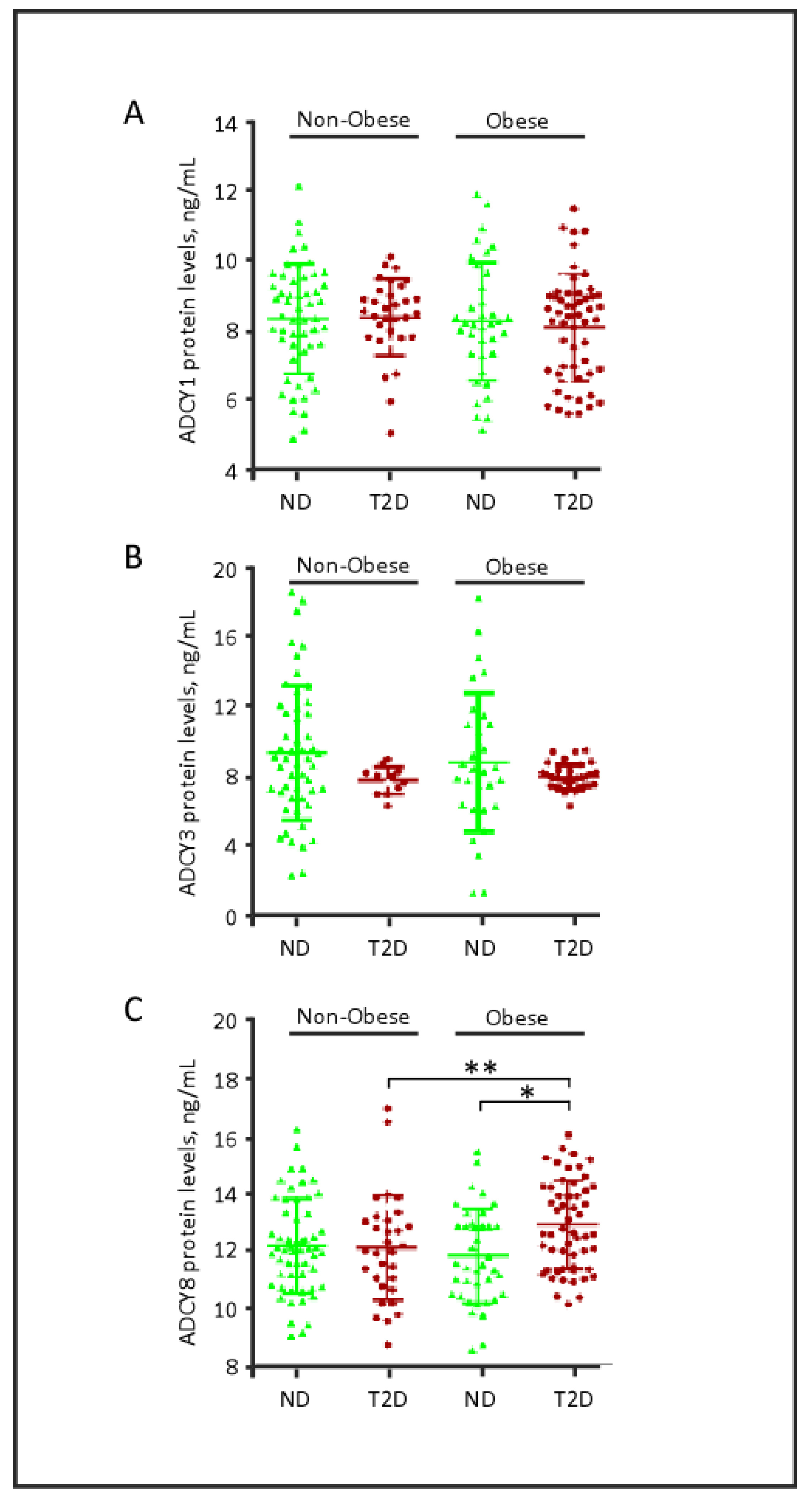
2.2. Plasma Levels of ADCY Proteins, Obesity and Diabetes
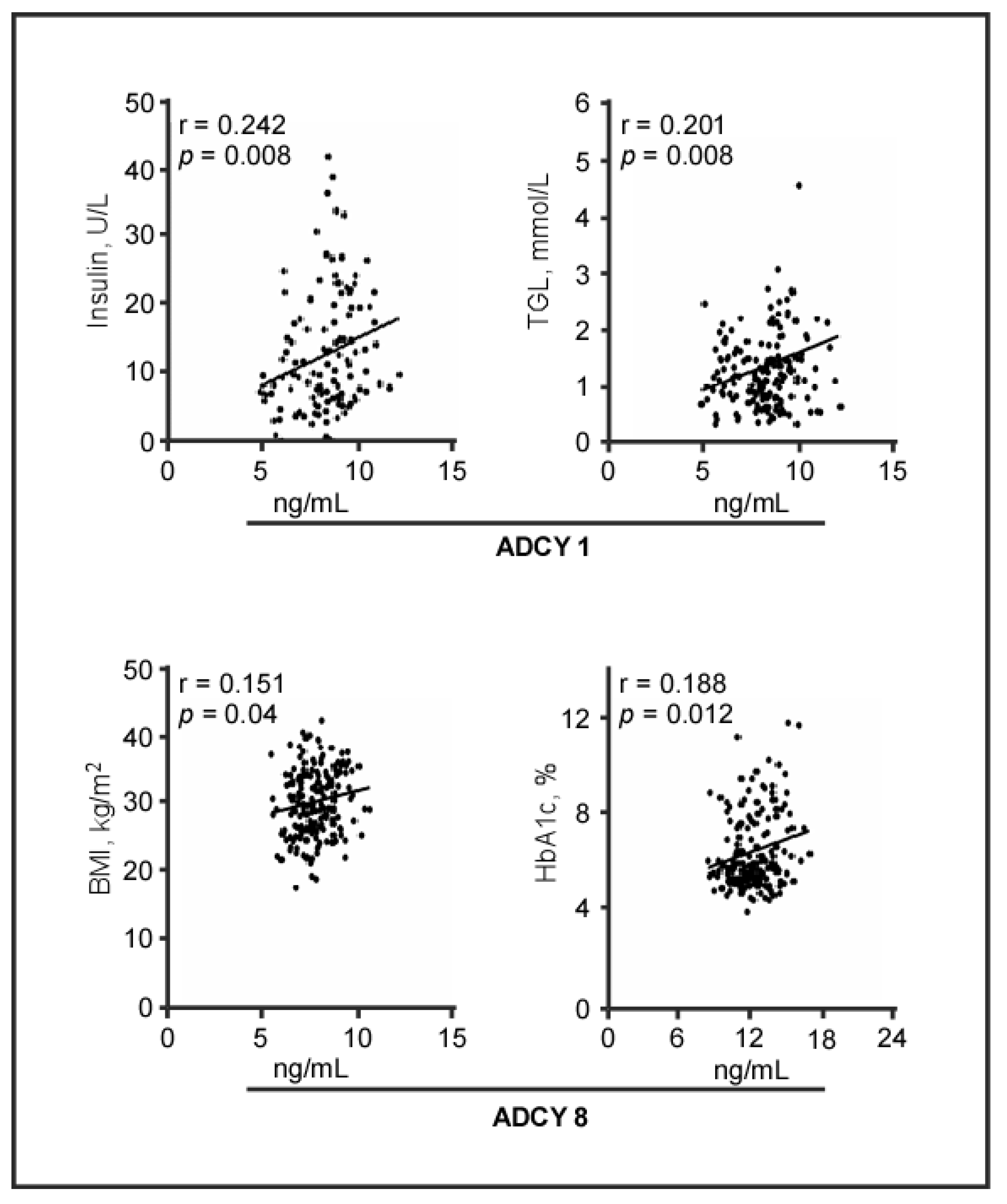
2.3. Plasma Levels of ADCY Proteins and Biomedical Parameters
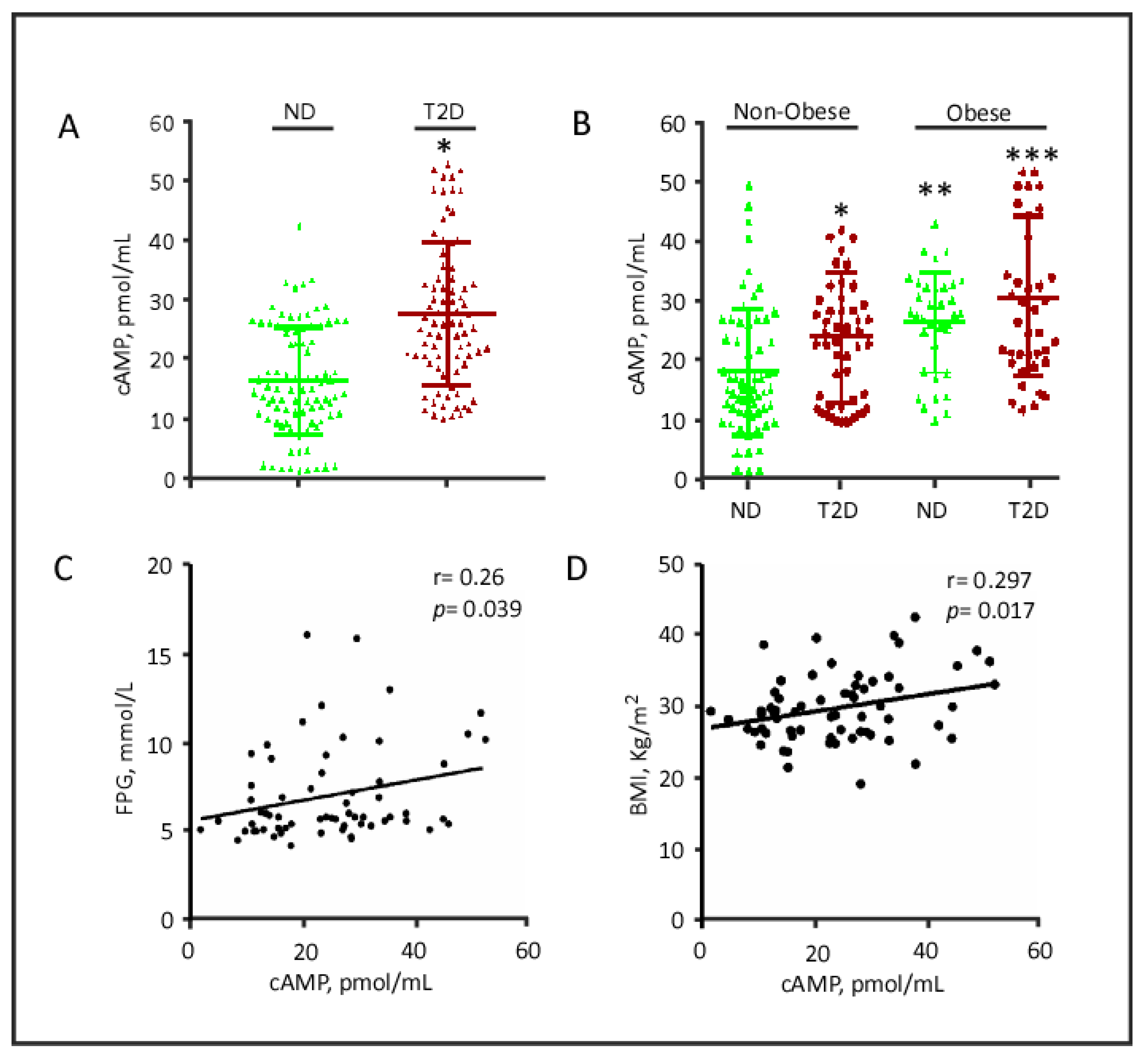
2.4. cAMP Plasma Levels
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Study Design
5.2. Study Population
5.3. Blood Collection, Anthropometric, and Biochemical Measurements
5.4. Quantitative Analysis of Plasma Levels of Adenylate Cyclase Proteins
5.5. Quantitative Analysis of cAMP Plasma Levels
5.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Ethics Approval and Consent to Participate
Abbreviations
| ADCY | adenylate cyclase |
| BMI | body mass index |
| cAMP | cyclic adenosine monophosphate |
| FPG | fasting plasma glucose |
| HbA1c | glycated hemoglobin |
| T2D | type 2 diabetes |
References
- Arroyo-Johnson, C.; Mincey, K. Obesity Epidemiology Worldwide. Gastroenterol. Clin. N. Am. 2016, 45, 571–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelaal, M.; Le Roux, C.W.; Docherty, N.G. Morbidity and mortality associated with obesity. Ann. Transl. Med. 2017, 5, 161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phelan, S.M.; Burgess, D.J.; Yeazel, M.W.; Hellerstedt, W.L.; Griffin, J.M.; Van Ryn, M. Impact of weight bias and stigma on quality of care and outcomes for patients with obesity. Obes. Rev. 2015, 16, 319–326. [Google Scholar] [CrossRef]
- de Luca, C.; Olefsky, J.M. Inflammation and insulin resistance. FEBS Lett. 2008, 582, 97–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matthews, D.R. Insulin resistance and beta-cell function--a clinical perspective. Diabetes Obes. Metab. 2001, 3, 28–33. [Google Scholar] [CrossRef]
- Ashcroft, F.M.; Rorsman, P. Diabetes mellitus and the beta cell: The last ten years. Cell 2012, 148, 1160–1171. [Google Scholar] [CrossRef] [Green Version]
- Rhodes, C.J. Type 2 diabetes-a matter of beta-cell life and death? Science 2005, 307, 380–384. [Google Scholar] [CrossRef]
- Ogurtsova, K.; Fernandes, J.D.R.; Huang, Y.; Linnenkamp, U.; Guariguata, L.; Cho, N.; Cavan, D.; Shaw, J.; Makaroff, L.E. IDF Diabetes Atlas: Global estimates for the prevalence of diabetes for 2015 and 2040. Diabetes Res. Clin. Pract. 2017, 128, 40–50. [Google Scholar] [CrossRef] [Green Version]
- Guariguata, L.; Whiting, D.; Hambleton, I.; Beagley, J.; Linnenkamp, U.; Shaw, J. Global estimates of diabetes prevalence for 2013 and projections for 2035. Diabetes Res. Clin. Pract. 2014, 103, 137–149. [Google Scholar] [CrossRef]
- Tong, T.; Shen, Y.; Lee, H.-W.; Yu, R.; Park, T. Adenylyl cyclase 3 haploinsufficiency confers susceptibility to diet-induced obesity and insulin resistance in mice. Sci. Rep. 2016, 6, 34179. [Google Scholar] [CrossRef] [Green Version]
- Hanoune, J.; Defer, N. Regulation and role of adenylyl cyclase isoforms. Annu. Rev. Pharmacol. Toxicol. 2001, 41, 145–174. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, M.-G.; Seuwen, K. Characterization of the human adenylyl cyclase gene family: cDNA, gene structure, and tissue distribution of the nine isoforms. J. Recept. Signal Transduct. 2002, 22, 79–110. [Google Scholar] [CrossRef] [PubMed]
- Tengholm, A.; Gylfe, E. cAMP signalling in insulin and glucagon secretion. Diabetes Obes. Metab. 2017, 19, 42–53. [Google Scholar] [CrossRef] [Green Version]
- Choi, L.J.; Jenikova, G.; Hanson, E.; Spehlmann, M.E.; Boehling, N.S.; Kirstein, S.L.; Bundey, R.A.; Smith, J.R.; Insel, P.A.; Eckmann, L. Coordinate Down-regulation of Adenylyl Cyclase Isoforms and the Stimulatory G Protein (Gs) in Intestinal Epithelial Cell Differentiation. J. Biol. Chem. 2010, 285, 12504–12511. [Google Scholar] [CrossRef] [Green Version]
- Cooper, D.M.F. Regulation and organization of adenylyl cyclases and cAMP. Biochem. J. 2003, 375, 517–529. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Halim, S.M.; Guenifi, A.; He, B.; Yang, B.; Mustafa, M.; Hojeberg, B.; Hillert, J.; Bakhiet, M.; Efendic, S. Mutations in the promoter of adenylyl cyclase (AC)-III gene, overexpression of AC-III mRNA, and enhanced cAMP generation in islets from the spontaneously diabetic GK rat model of type 2 diabetes. Diabetes 1998, 47, 498–504. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Li, V.; Chan, G.C.K.; Phan, T.; Nudelman, A.S.; Xia, Z.; Storm, D.R. Adult Type 3 Adenylyl Cyclase–Deficient Mice Are Obese. PLoS ONE 2009, 4, e6979. [Google Scholar] [CrossRef] [Green Version]
- Pitman, J.L.; Wheeler, M.C.; Lloyd, D.J.; Walker, J.R.; Glynne, R.J.; Gekakis, N. A Gain-of-Function Mutation in Adenylate Cyclase 3 Protects Mice from Diet-Induced Obesity. PLoS ONE 2014, 9, e110226. [Google Scholar] [CrossRef]
- Yang, B.; He, B.; Abdel-Halim, S.M.; Tibell, A.; Brendel, M.D.; Bretzel, R.G.; Efendić, S.; Hillert, J. Molecular Cloning of a Full-Length cDNA for Human Type 3 Adenylyl Cyclase and Its Expression in Human Islets. Biochem. Biophys. Res. Commun. 1999, 254, 548–551. [Google Scholar] [CrossRef]
- Saeed, S.; Bonnefond, A.; Tamanini, F.; Mirza, M.U.; Manzoor, J.; Janjua, M.Q.; Din, S.M.; Gaitan, J.; Milochau, A.; Durand, E.; et al. Loss-of-function mutations in ADCY3 cause monogenic severe obesity. Nat. Genet. 2018, 50, 175–179. [Google Scholar] [CrossRef]
- Tian, Y.; Peng, B.; Fu, X.-H. New ADCY3 Variants Dance in Obesity Etiology. Trends Endocrinol. Metab. 2018, 29, 361–363. [Google Scholar] [CrossRef]
- Guenifi, A.; Portela-Gomes, G.M.; Grimelius, L.; Efendić, S.; Abdel-Halim, S.M. Adenylyl cyclase isoform expression in non-diabetic and diabetic Goto-Kakizaki (GK) rat pancreas. Evidence for distinct overexpression of type-8 adenylyl cyclase in diabetic GK rat islets. Histochem. Cell Biol. 2000, 113, 81–89. [Google Scholar] [CrossRef]
- Portela-Gomes, G.M.; Abdel-Halim, S.M. Overexpression of Gs Proteins and Adenylyl Cyclase in Normal and Diabetic Islets. Pancreas 2002, 25, 176–181. [Google Scholar] [CrossRef] [PubMed]
- Portela-Gomes, G.M.; Grimelius, L.; Johansson, H.; Efendic, S.; Wester, K.; Abdel-Halim, S.M. Increased expression of adenylyl cyclase isoforms in the adrenal gland of diabetic Goto-Kakizaki rat. Appl. Immunohistochem. Mol. Morphol. 2002, 10, 387–392. [Google Scholar] [CrossRef] [PubMed]
- Roger, B.; Papin, J.; Vacher, P.; Raoux, M.; Mulot, A.; Dubois, M.; Kerr-Conte, J.; Voy, B.H.; Pattou, F.; Charpentier, G.; et al. Adenylyl cyclase 8 is central to glucagon-like peptide 1 signalling and effects of chronically elevated glucose in rat and human pancreatic beta cells. Diabetologia 2010, 54, 390–402. [Google Scholar] [CrossRef] [Green Version]
- Raoux, M.; Vacher, P.; Papin, J.; Picard, A.; Kostrzewa, E.; Devin, A.; Gaitan, J.; Limon, I.; Kas, M.J.; Magnan, C.; et al. Multilevel control of glucose homeostasis by adenylyl cyclase 8. Diabetologia 2015, 58, 749–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hodson, D.J.; Mitchell, R.; Marselli, L.; Pullen, T.J.; Brias, S.G.; Semplici, F.; Everett, K.L.; Cooper, D.M.; Bugliani, M.; Marchetti, P.; et al. ADCY5 Couples Glucose to Insulin Secretion in Human Islets. Diabetes 2014, 63, 3009–3021. [Google Scholar] [CrossRef] [Green Version]
- Eizirik, D.L.; Sammeth, M.; Bouckenooghe, T.; Bottu, G.; Sisino, G.; Igoillo-Esteve, M.; Ortis, F.; Santin, I.; Colli, M.L.; Barthson, J.; et al. The Human Pancreatic Islet Transcriptome: Expression of Candidate Genes for Type 1 Diabetes and the Impact of Pro-Inflammatory Cytokines. PLoS Genet. 2012, 8, e1002552. [Google Scholar] [CrossRef]
- Guenifi, A.; Abdel-Halim, S.M. Glucose Enhances Adenylyl Cyclase Responses in Normal But Not Diabetic GK Rat Islets. Pancreas 2001, 22, 65–68. [Google Scholar] [CrossRef]
- Grarup, N.; Moltke, I.; Andersen, M.K.; Dalby, M.; Vitting-Seerup, K.; Kern, T.; Mahendran, Y.; Jørsboe, E.; Larsen, C.V.L.; Dahl-Petersen, I.K.; et al. Loss-of-function variants in ADCY3 increase risk of obesity and type 2 diabetes. Nat. Genet. 2018, 50, 172–174. [Google Scholar] [CrossRef]
- Nordman, S.; Abulaiti, A.; Hilding, A.; Långberg, E.-C.; Humphreys, K.; Östenson, C.-G.; Efendić, S.; Gu, H.F. Genetic variation of the adenylyl cyclase 3 (AC3) locus and its influence on type 2 diabetes and obesity susceptibility in Swedish men. Int. J. Obes. 2007, 32, 407–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, Y.; Li, Z.; Liang, S.; Li, Y.; Yang, L.; Lu, M.; Gu, H.F.; Xia, N. Hepatic adenylate cyclase 3 is upregulated by Liraglutide and subsequently plays a protective role in insulin resistance and obesity. Nutr. Diabetes 2016, 6, e191. [Google Scholar] [CrossRef] [PubMed]
- Sung, Y.J.; Pérusse, L.; Sarzynski, M.A.; Fornage, M.; Sidney, S.; Sternfeld, B.; Rice, T.; Terry, J.G.; Jacobs, D.R.; Katzmarzyk, P.T.; et al. Genome-wide association studies suggest sex-specific loci associated with abdominal and visceral fat. Int. J. Obes. 2015, 40, 662–674. [Google Scholar] [CrossRef] [Green Version]
- Delmeire, D.; Flamez, D.; Hinke, S.A.; Cali, J.J.; Pipeleers, D.; Schuit, F. Type VIII adenylyl cyclase in rat beta cells: Coincidence signal detector/generator for glucose and GLP-1. Diabetologia 2003, 46, 1383–1393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smolarz, M.; Pietrowska, M.; Matysiak, N.; Mielańczyk, Ł; Widlak, P. Proteome Profiling of Exosomes Purified from a Small Amount of Human Serum: The Problem of Co-Purified Serum Components. Proteomes 2019, 7, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Kojima, K.; Mobley, J.A.; West, A.B. Proteomic analysis of urinary extracellular vesicles reveal biomarkers for neurologic disease. EBioMedicine 2019, 45, 351–361. [Google Scholar] [CrossRef] [Green Version]
- Madsen, L.; Kristiansen, K. The importance of dietary modulation of cAMP and insulin signaling in adipose tissue and the development of obesity. Ann. N. Y. Acad. Sci. 2010, 1190, 1–14. [Google Scholar] [CrossRef]
- Rogne, M.; Taskén, K. Compartmentalization of cAMP Signaling in Adipogenesis, Lipogenesis, and Lipolysis. Horm. Metab. Res. 2014, 46, 833–840. [Google Scholar] [CrossRef]
- Cao, W.; Daniel, K.W.; Robidoux, J.; Puigserver, P.; Medvedev, A.; Bai, X.; Floering, L.M.; Spiegelman, B.M.; Collins, S. p38 Mitogen-Activated Protein Kinase Is the Central Regulator of Cyclic AMP-Dependent Transcription of the Brown Fat Uncoupling Protein 1 Gene. Mol. Cell. Biol. 2004, 24, 3057–3067. [Google Scholar] [CrossRef] [Green Version]
- Robinson, R.T.; Harris, N.D.; Ireland, R.H.; Lee, S.; Newman, C.; Heller, S.R. Mechanisms of abnormal cardiac repolarization during insulin-induced hypoglycemia. Diabetes 2003, 52, 1469–1474. [Google Scholar] [CrossRef] [Green Version]
- Landstrom, A.P.; Dobrev, B.; Wehrens, X.H. Calcium Signaling and Cardiac Arrhythmias. Circ. Res. 2017, 120, 1969–1993. [Google Scholar] [CrossRef] [PubMed]
- Abubaker, J.; Tiss, A.; Abu-Farha, M.; Al-Ghimlas, F.; Al-Khairi, I.; Baturcam, E.; Cherian, P.; Elkum, N.; Hammad, M.; John, J.; et al. DNAJB3/HSP-40 cochaperone is downregulated in obese humans and is restored by physical exercise. PLoS ONE 2013, 87, e69217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abu-Farha, M.; Tiss, A.; Abubaker, J.; Khadir, A.; Al-Ghimlas, F.; Al-Khairi, I.; Baturcam, E.; Cherian, P.; Elkum, N.; Hammad, M.; et al. Proteomics analysis of human obesity reveals the epigenetic factor HDAC4 as a potential target for obesity. PLoS ONE 2013, 8, e75342. [Google Scholar] [CrossRef] [PubMed]
- Cherian, P.T.; Al-Khairi, I.; Sriraman, D.; Al-Enezi, A.; Al-Sultan, D.; AlOtaibi, M.; Al-Enezi, S.; Tuomilehto, J.; Al-Mulla, F.; Abubaker, J.A.; et al. Increased Circulation and Adipose Tissue Levels of DNAJC27/RBJ in Obesity and Type 2-Diabetes. Front. Endocrinol. Lausanne 2018, 9, 423. [Google Scholar] [CrossRef] [PubMed]
- Abu-Farha, M.; Cherian, P.; Qaddoumi, M.G.; AlKhairi, I.; Sriraman, D.; Alanbaei, M.; Abubaker, J. Increased plasma and adipose tissue levels of ANGPTL8/Betatrophin and ANGPTL4 in people with hypertension. Lipids Health Dis. 2018, 17, 35. [Google Scholar] [CrossRef] [Green Version]
- Abu-Farha, M.; Cherian, P.; Al-Khairi, I.; Madhu, D.; Tiss, A.; Warsam, S.; Alhubail, A.; Sriraman, D.; Al-Refaei, F.; Abubaker, J. Plasma and adipose tissue level of angiopoietin-like 7 (ANGPTL7) are increased in obesity and reduced after physical exercise. PLoS ONE 2017, 12, e0173024. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phenotype | T2D (n) | ND (n) | p-value |
|---|---|---|---|
| Age, years | 52.13 ± 9.15 (92) | 43.3 ± 12.782 (96) | <0.001 |
| Gender (F/M) | 43/49 | 69/27 | 0.143 |
| Height, m | 1.65 ± 0.08 (91) | 1.63 ± 0.10 (96) | 0.073 |
| Weight, Kg | 87.06 ± 14.82 (91) | 78.36 ± 18.78 (96) | 0.001 |
| BMI, Kg/m2 | 31.60 ± 4.32 (92) | 29.26 ± 5.36 (96) | 0.001 |
| WHR | 0.96 ± 0.19 (64) | 0.85 ± 0.09 (67) | <0.001 |
| FPG, mmol/L | 8.35 ± 3.03 (91) | 5.30 ± 0.82 (94) | <0.001 |
| HbA1c, % | 7.75 ± 1.84 (90) | 5.58 ± 0.50 (92) | <0.001 |
| Total Cholesterol, mmol/L | 4.96 ± 1.35 (90) | 5.20 ± 0.94 (95) | 0.158 |
| HDL, mmol/L | 1.19 ± 0.48 (89) | 1.41 ± 0.41 (95) | 0.001 |
| LDL, mmol/L | 3.08 ± 1.15 (88) | 3.27 ± 0.87 (95) | 0.226 |
| TGL, mmol/L | 1.63 ± 1.20 (90) | 1.13 ± 0.67 (94) | 0.001 |
| Insulin, U/L | 14.86 ± 10.02 (70) | 9.70 ± 5.44 (57) | <0.001 |
| Phenotype | T2D | ND | ||||
|---|---|---|---|---|---|---|
| Non-Obese (n) | Obese (n) | p-value | Non-obese (n) | Obese (n) | p-value | |
| Age, years | 51.85 ± 9.37 (34) | 52.29 ± 9.10 (58) | 0.827 | 42.00 ± 12.30 (56) | 45.13 ± 13.37 (40) | 0.247 |
| Gender (F/M) | 14/20 | 29/29- | 0.517 | 42/14 | 27/13 | 0.492 |
| Height, m | 1.65 ± 0.07 (33) | 1.66 ± 0.09 (58) | 0.821 | 1.62 ± 0.09 (56) | 1.65 ± 0.11 (40) | 0.155 |
| Weight, Kg | 73.69 ± 9.00 (33) | 94.66 ± 11.79 (58) | <0.001 | 67.12 ± 10.95 (56) | 94.10 ± 15.96 (40) | <0.001 |
| BMI, Kg/m2 | 26.80 ± 2.33 (34) | 34.41 ± 2.22 (58) | <0.001 | 25.57 ± 2.89 (56) | 34.43 ± 3.33 (40) | <0.001 |
| WHR | 0.91 ± 0.06 (22) | 0.98 ± 0.23 (42) | 0.077 | 0.83 ± 0.10 (39) | 0.88 ± 0.07 (28) | 0.020 |
| FPG, mmol/L | 7.44 ± 2.35 (33) | 8.87 ± 3.26 (58) | 0.018 | 5.10 ± 0.43 (54) | 5.57 ± 1.11 (40) | 0.016 |
| HbA1c, % | 6.76 ± 1.26 (32) | 8.30 ± 1.88 (58) | <0.001 | 5.54 ± 0.42 (52) | 5.63 ± 0.59 (40) | 0.447 |
| Total Cholesterol, mmol/L | 4.94 ± 1.65 (33) | 4.97 ± 1.15 (57) | 0.919 | 5.23 ± 0.93 (55) | 5.17 ± 0.95 (40) | 0.770 |
| HDL, mmol/L | 1.22 ± 0.63 (33) | 1.17 ± 0.36 (56) | 0.658 | 1.46 ± 0.45 (55) | 1.35 ± 0.36 (40) | 0.178 |
| LDL, mmol/L | 3.12 ± 1.33 (33) | 3.06 ± 1.04 (55) | 0.836 | 3.25 ± 0.86 (55) | 3.29 ± 0.89 (40) | 0.799 |
| TGL, mmol/L | 1.56 ± 1.11 (33) | 1.67 ± 1.26 (57) | 0.652 | 1.06 ± 0.72 (54) | 1.22 ± 0.61 (40) | 0.252 |
| Insulin, U/L | 15.68 ± 11.05 (27) | 14.34 ± 9.41 (43) | 0.603 | 9.39 ± 5.94 (39) | 10.38 ± 4.23 (18) | 0.474 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdel-Halim, S.M.; Al Madhoun, A.; Nizam, R.; Melhem, M.; Cherian, P.; Al-Khairi, I.; Haddad, D.; Abu-Farha, M.; Abubaker, J.; Bitar, M.S.; et al. Increased Plasma Levels of Adenylate Cyclase 8 and cAMP Are Associated with Obesity and Type 2 Diabetes: Results from a Cross-Sectional Study. Biology 2020, 9, 244. https://doi.org/10.3390/biology9090244
Abdel-Halim SM, Al Madhoun A, Nizam R, Melhem M, Cherian P, Al-Khairi I, Haddad D, Abu-Farha M, Abubaker J, Bitar MS, et al. Increased Plasma Levels of Adenylate Cyclase 8 and cAMP Are Associated with Obesity and Type 2 Diabetes: Results from a Cross-Sectional Study. Biology. 2020; 9(9):244. https://doi.org/10.3390/biology9090244
Chicago/Turabian StyleAbdel-Halim, Samy M., Ashraf Al Madhoun, Rasheeba Nizam, Motasem Melhem, Preethi Cherian, Irina Al-Khairi, Dania Haddad, Mohamed Abu-Farha, Jehad Abubaker, Milad S. Bitar, and et al. 2020. "Increased Plasma Levels of Adenylate Cyclase 8 and cAMP Are Associated with Obesity and Type 2 Diabetes: Results from a Cross-Sectional Study" Biology 9, no. 9: 244. https://doi.org/10.3390/biology9090244
APA StyleAbdel-Halim, S. M., Al Madhoun, A., Nizam, R., Melhem, M., Cherian, P., Al-Khairi, I., Haddad, D., Abu-Farha, M., Abubaker, J., Bitar, M. S., & Al-Mulla, F. (2020). Increased Plasma Levels of Adenylate Cyclase 8 and cAMP Are Associated with Obesity and Type 2 Diabetes: Results from a Cross-Sectional Study. Biology, 9(9), 244. https://doi.org/10.3390/biology9090244