Phosphoinositide Lipids in Ocular Tissues



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
2.2.1. Isolation of Rod Outer Segments by Discontinuous Sucrose Density Gradient Centrifugation
2.2.2. Isolation of Bovine Retinal Pigment Epithelium
2.2.3. Preparation of Bovine Cornea for the Isolation of Phosphoinositides
2.2.4. Protein-Lipid Overlay Assay
2.2.5. Extraction of Phosphoinositides from Bovine Cornea, Retina, and RPE
2.2.6. The Phosphoinositide ELISA Assay
2.2.7. Recombinant Enzyme ELISA
2.3. Statistical Analysis
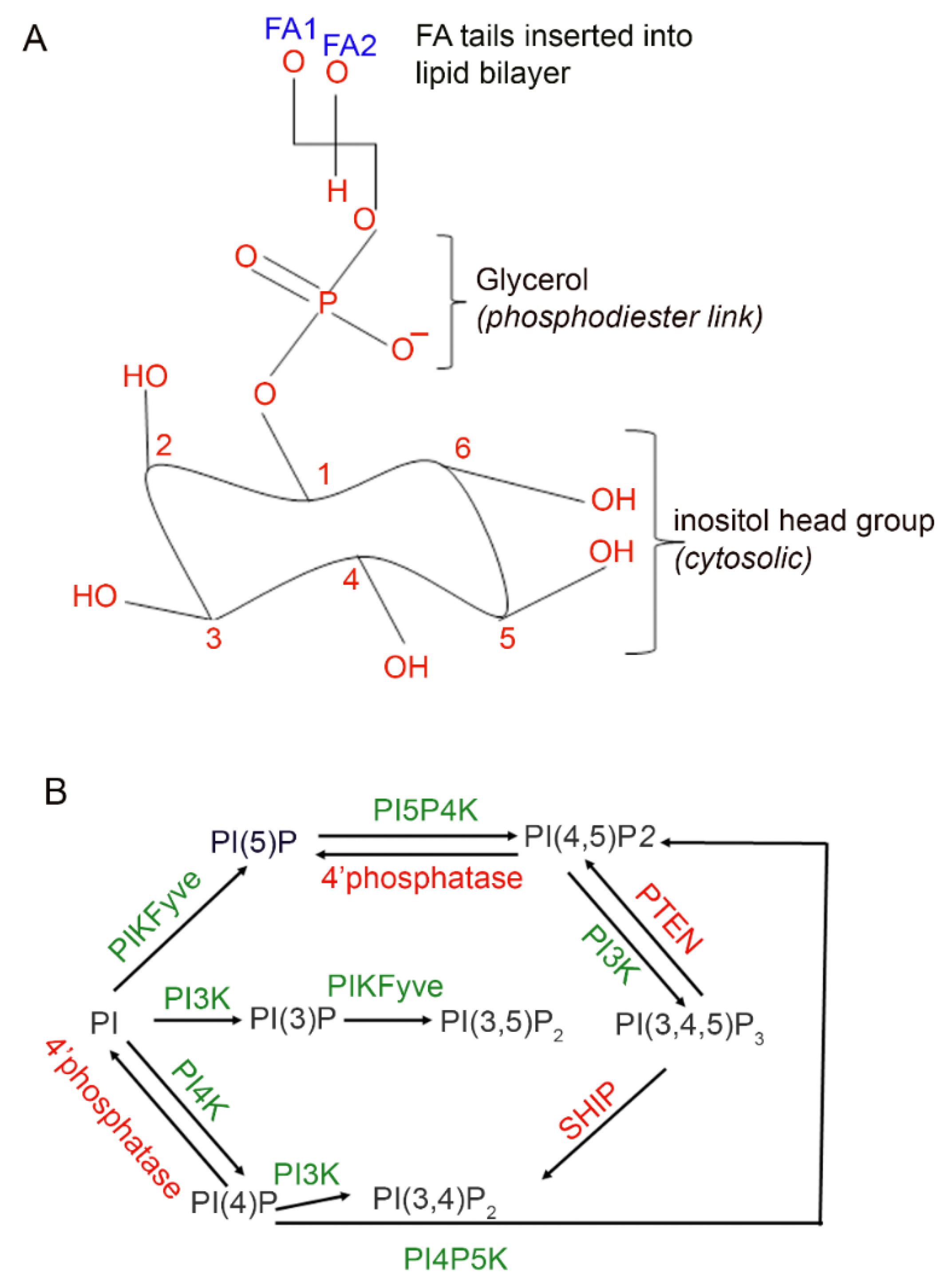
3. Results
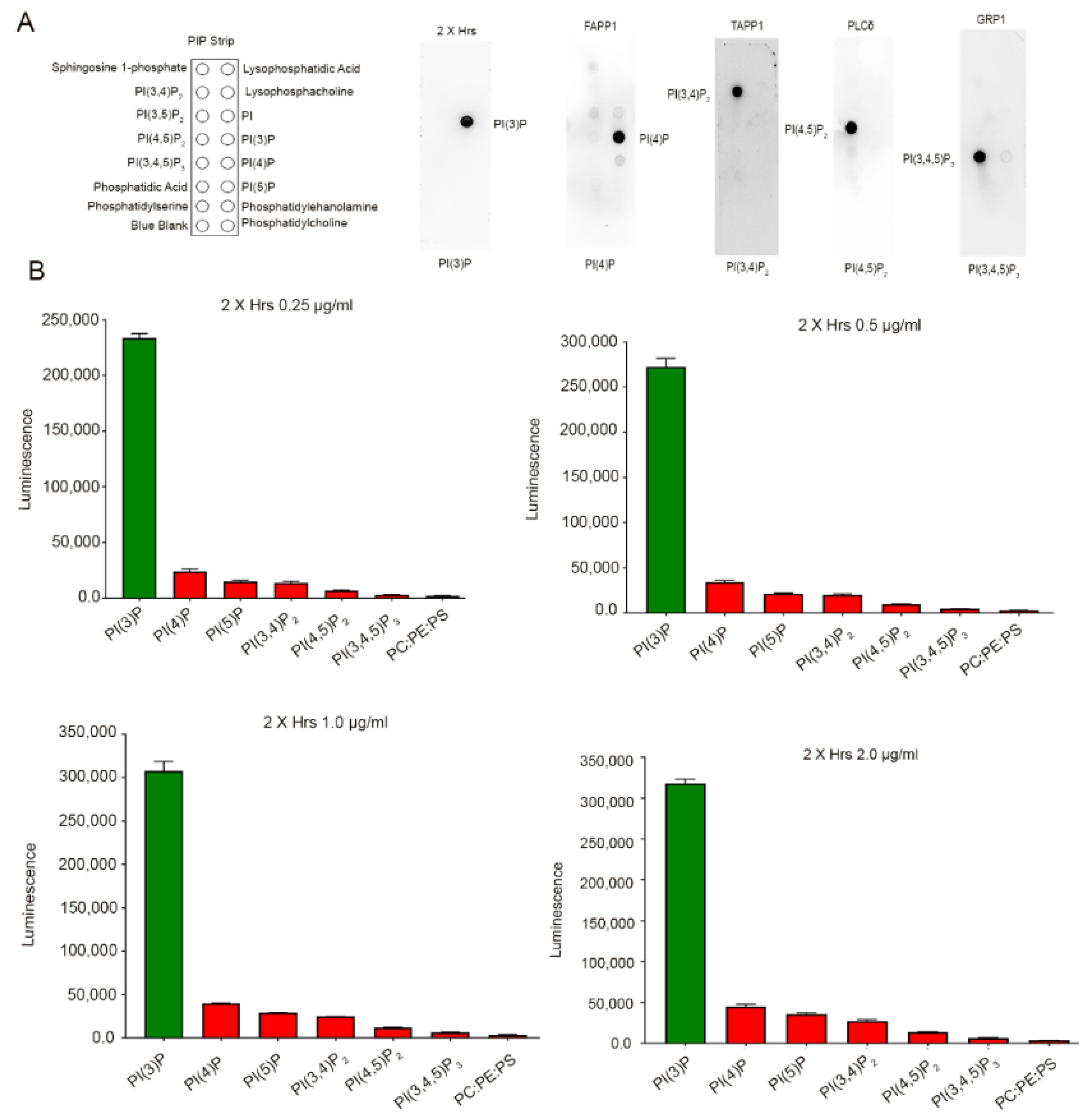
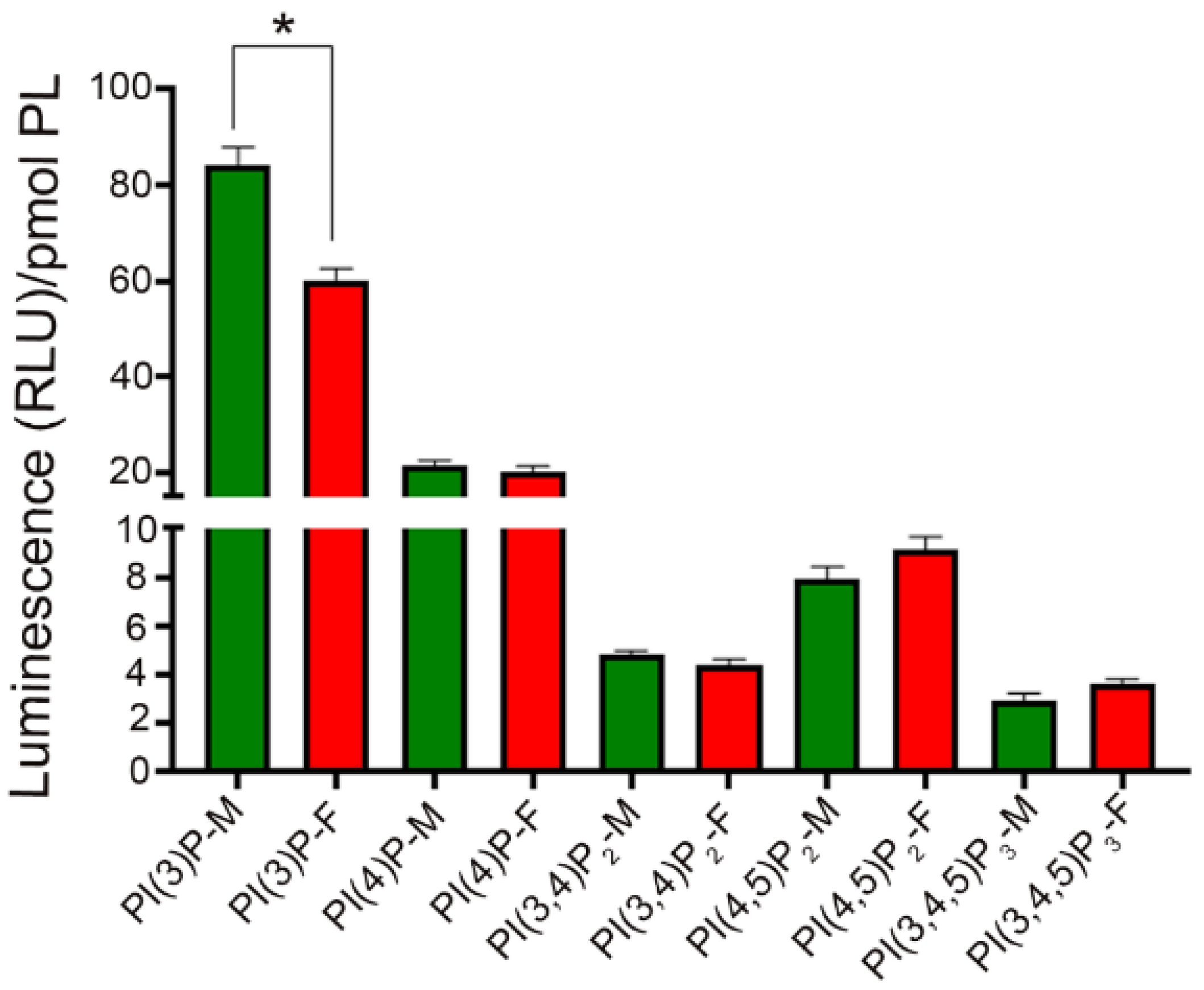
3.1. Binding Specificity of PI-Binding Protein to Specific Phosphoinositide Species
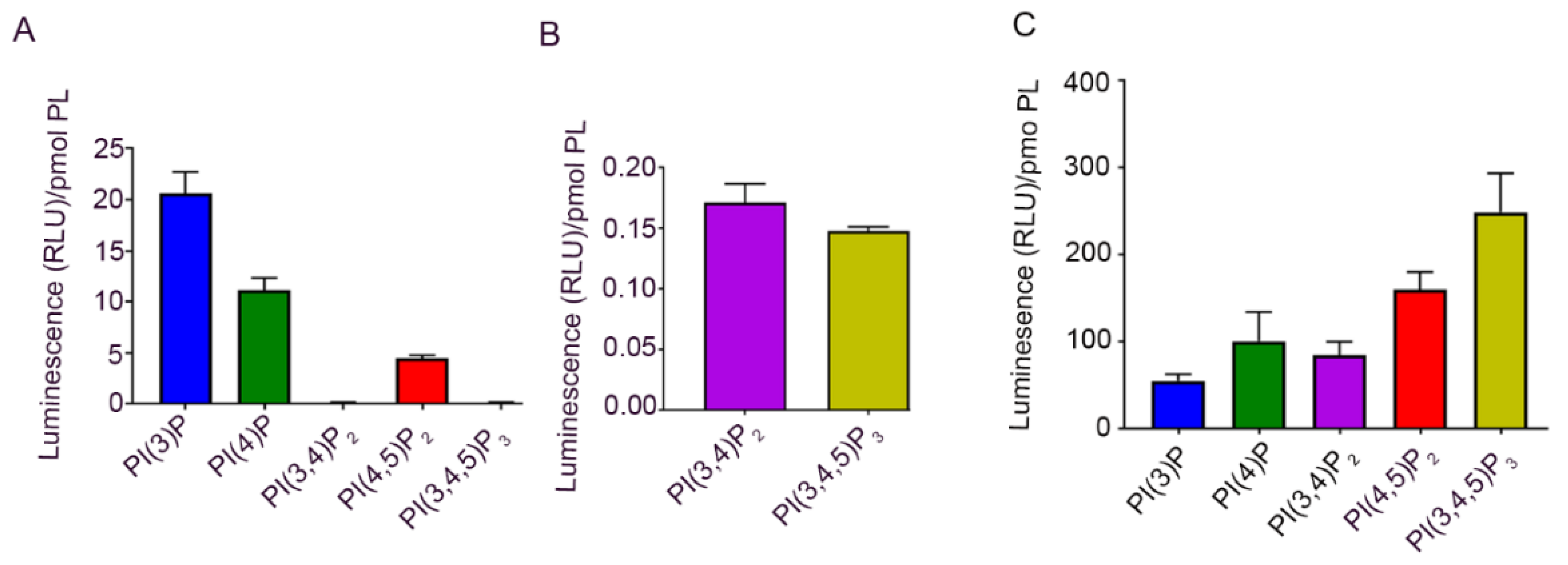
3.2. Phosphoinositide Lipids in the Cornea
3.3. Phosphoinositide Lipids in the RPE
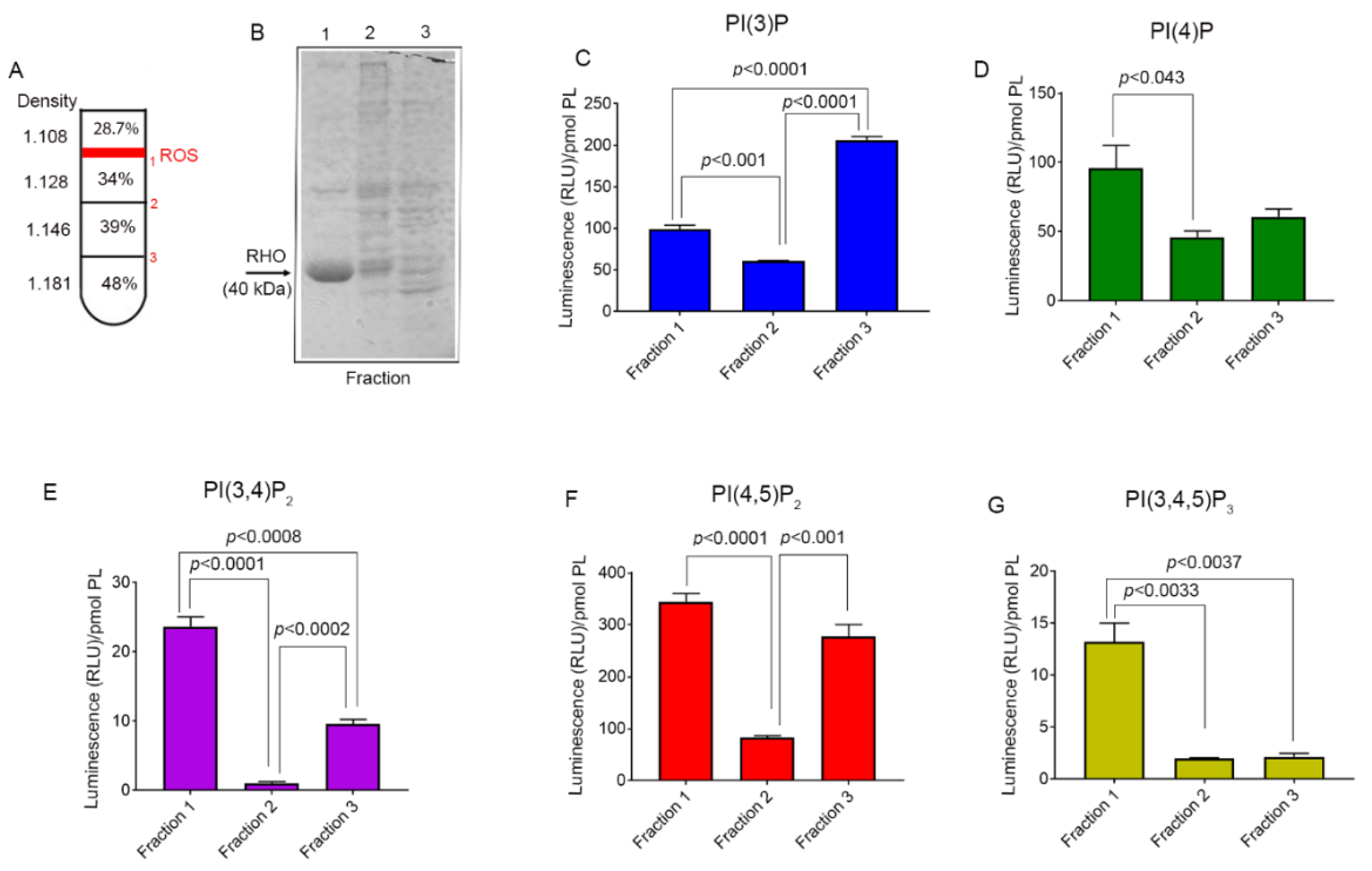
3.4. Phosphoinositide Lipids in the Retina
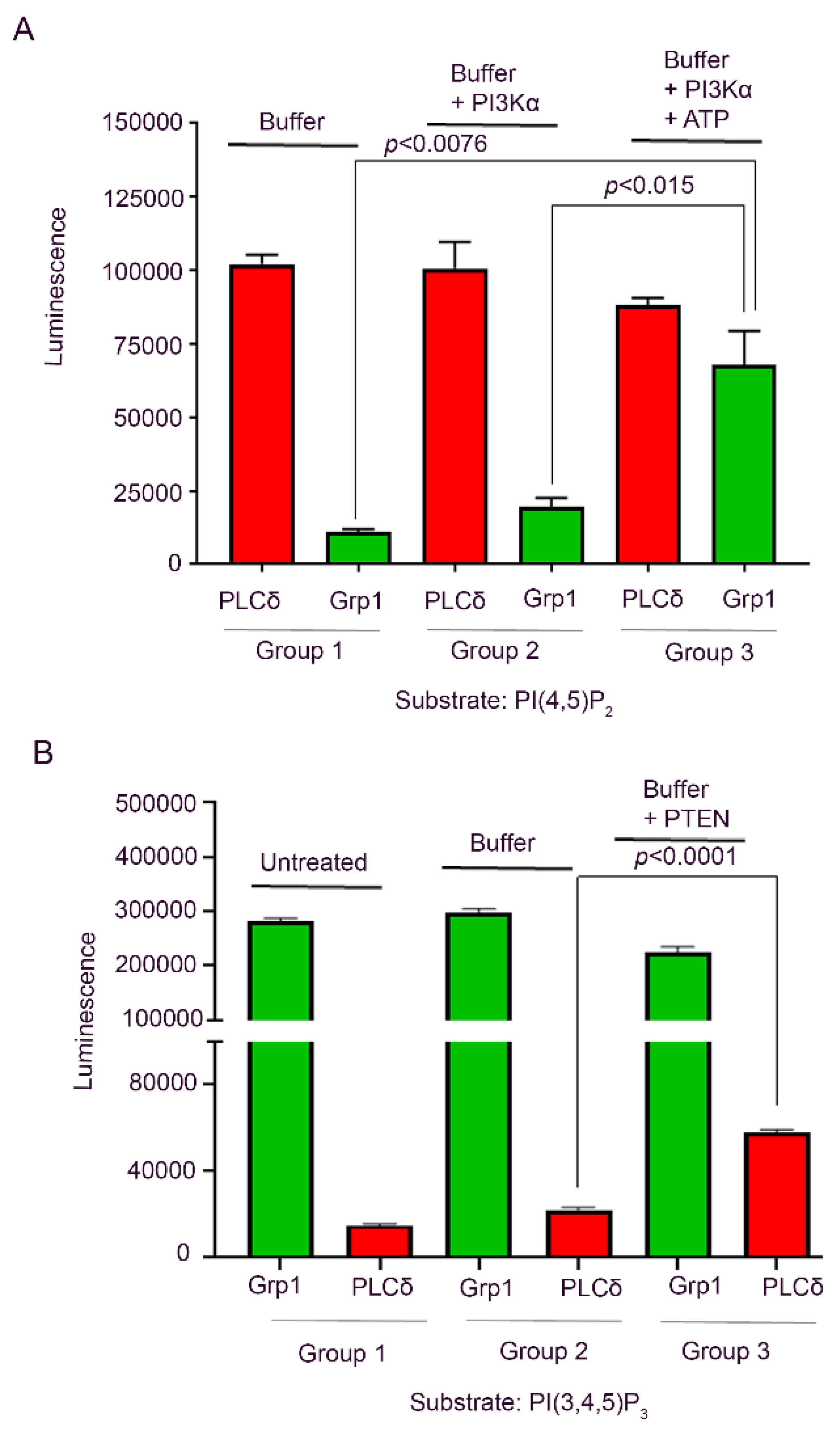
3.5. Recombinant PI3K and PTEN Enzyme ELISA
4. Discussion and Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Balla, T. Phosphoinositides: Tiny lipids with giant impact on cell regulation. Physiol. Rev. 2013, 93, 1019–1137. [Google Scholar] [CrossRef] [PubMed]
- Rusten, T.E.; Stenmark, H. Analyzing phosphoinositides and their interacting proteins. Nat. Methods 2006, 3, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Rajala, R.V. Phosphoinositide 3-kinase signaling in the vertebrate retina. J. Lipid Res. 2010, 51, 4–22. [Google Scholar] [CrossRef] [PubMed]
- Di Paolo, G.; De Camilli, P. Phosphoinositides in cell regulation and membrane dynamics. Nature 2006, 443, 651–657. [Google Scholar] [CrossRef] [PubMed]
- Bazan, H.E.; King, W.D.; Rossowska, M. Metabolism of phosphoinositides and inositol polyphosphates in rabbit corneal epithelium. Curr. Eye Res. 1985, 4, 793–801. [Google Scholar] [CrossRef]
- He, F.; Agosto, M.A.; Anastassov, I.A.; Tse, D.Y.; Wu, S.M.; Wensel, T.G. Phosphatidylinositol-3-phosphate is light-regulated and essential for survival in retinal rods. Sci. Rep. 2016, 6, 26978. [Google Scholar] [CrossRef]
- Rajala, R.V.; McClellan, M.E.; Ash, J.D.; Anderson, R.E. In vivo regulation of phosphoinositide 3-kinase in retina through light-induced tyrosine phosphorylation of the insulin receptor beta-subunit. J. Biol. Chem. 2002, 277, 43319–43326. [Google Scholar] [CrossRef]
- Anderson, R.E.; Hollyfield, J.G. Light stimulates the incorporation of inositol into phosphatidylinositol in the retina. Biochim. Biophys. Acta 1981, 665, 619–622. [Google Scholar] [CrossRef]
- Anderson, R.E.; Maude, M.B.; Kelleher, P.A.; Rayborn, M.E.; Hollyfield, J.G. Phosphoinositide metabolism in the retina: Localization to horizontal cells and regulation by light and divalent cations. J. Neurochem. 1983, 41, 764–771. [Google Scholar] [CrossRef]
- He, F.; Nichols, R.M.; Kailasam, L.; Wensel, T.G.; Agosto, M.A. Critical Role for Phosphatidylinositol-3 Kinase Vps34/PIK3C3 in ON-Bipolar Cells. Investig. Ophthalmol. Vis. Sci. 2019, 60, 2861–2874. [Google Scholar] [CrossRef]
- He, F.; Agosto, M.A.; Nichols, R.M.; Wensel, T.G. Multiple phosphatidylinositol(3)phosphate roles in retinal pigment epithelium membrane recycling. bioRxiv 2020. [Google Scholar] [CrossRef]
- Giusto, N.M.; Pasquare, S.J.; Salvador, G.A.; Ilincheta de Boschero, M.G. Lipid second messengers and related enzymes in vertebrate rod outer segments. J. Lipid Res. 2010, 51, 685–700. [Google Scholar] [CrossRef] [PubMed]
- Wensel, T.G. Phosphoinositides in Retinal Function and Disease. Cells 2020, 9, 886. [Google Scholar] [CrossRef] [PubMed]
- Womack, K.B.; Gordon, S.E.; He, F.; Wensel, T.G.; Lu, C.C.; Hilgemann, D.W. Do phosphatidylinositides modulate vertebrate phototransduction? J. Neurosci. 2000, 20, 2792–2799. [Google Scholar] [CrossRef]
- He, F.; Mao, M.; Wensel, T.G. Enhancement of phototransduction g protein-effector interactions by phosphoinositides. J. Biol. Chem. 2004, 279, 8986–8990. [Google Scholar] [CrossRef]
- Deretic, D.; Traverso, V.; Parkins, N.; Jackson, F.; Rodriguez de Turco, E.B.; Ransom, N. Phosphoinositides, ezrin/moesin, and rac1 regulate fusion of rhodopsin transport carriers in retinal photoreceptors. Mol. Biol. Cell 2004, 15, 359–370. [Google Scholar] [CrossRef]
- Lee, S.J.; Montell, C. Light-dependent translocation of visual arrestin regulated by the NINAC myosin III. Neuron 2004, 43, 95–103. [Google Scholar] [CrossRef]
- Bright, S.R.; Rich, E.D.; Varnum, M.D. Regulation of human cone cyclic nucleotide-gated channels by endogenous phospholipids and exogenously applied phosphatidylinositol 3,4,5-trisphosphate. Mol. Pharmacol. 2007, 71, 176–183. [Google Scholar] [CrossRef]
- Brady, J.D.; Rich, E.D.; Martens, J.R.; Karpen, J.W.; Varnum, M.D.; Brown, R.L. Interplay between PIP3 and calmodulin regulation of olfactory cyclic nucleotide-gated channels. Proc. Natl. Acad. Sci. USA 2006, 103, 15635–15640. [Google Scholar] [CrossRef]
- Pizarro-Cerda, J.; Kuhbacher, A.; Cossart, P. Phosphoinositides and host-pathogen interactions. Biochim. Biophys. Acta 2015, 1851, 911–918. [Google Scholar] [CrossRef]
- Payrastre, B.; Gaits-Iacovoni, F.; Sansonetti, P.; Tronchere, H. Phosphoinositides and cellular pathogens. Sub-Cell. Biochem. 2012, 59, 363–388. [Google Scholar] [CrossRef]
- Peracino, B.; Balest, A.; Bozzaro, S. Phosphoinositides differentially regulate bacterial uptake and Nramp1-induced resistance to Legionella infection in Dictyostelium. J. Cell Sci. 2010, 123, 4039–4051. [Google Scholar] [CrossRef] [PubMed]
- Rajala, M.S.; Rajala, R.V.; Astley, R.A.; Butt, A.L.; Chodosh, J. Corneal cell survival in adenovirus type 19 infection requires phosphoinositide 3-kinase/Akt activation. J. Virol. 2005, 79, 12332–12341. [Google Scholar] [CrossRef] [PubMed]
- Mazzon, M.; Mercer, J. Lipid interactions during virus entry and infection. Cell. Microbiol. 2014, 16, 1493–1502. [Google Scholar] [CrossRef] [PubMed]
- Sher, A.A.; Glover, K.K.M.; Coombs, K.M. Zika Virus Infection Disrupts Astrocytic Proteins Involved in Synapse Control and Axon Guidance. Front. Microbiol. 2019, 10, 596. [Google Scholar] [CrossRef]
- Santagata, S.; Boggon, T.J.; Baird, C.L.; Gomez, C.A.; Zhao, J.; Shan, W.S.; Myszka, D.G.; Shapiro, L. G-protein signaling through tubby proteins. Science 2001, 292, 2041–2050. [Google Scholar] [CrossRef]
- Paloma, E.; Hjelmqvist, L.; Bayes, M.; Garcia-Sandoval, B.; Ayuso, C.; Balcells, S.; Gonzalez-Duarte, R. Novel mutations in the TULP1 gene causing autosomal recessive retinitis pigmentosa. Investig. Ophthalmol. Vis. Sci. 2000, 41, 656–659. [Google Scholar]
- Xi, Q.; Pauer, G.J.; Traboulsi, E.I.; Hagstrom, S.A. Mutation screen of the TUB gene in patients with retinitis pigmentosa and Leber congenital amaurosis. Exp. Eye Res. 2006, 83, 569–573. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Vicinanza, M.; Korolchuk, V.I.; Ashkenazi, A.; Puri, C.; Menzies, F.M.; Clarke, J.H.; Rubinsztein, D.C. PI(5)P regulates autophagosome biogenesis. Mol. Cell 2015, 57, 219–234. [Google Scholar] [CrossRef]
- Boisset, G.; Polok, B.K.; Schorderet, D.F. Characterization of pip5k3 fleck corneal dystrophy-linked gene in zebrafish. Gene Expr. Patterns 2008, 8, 404–410. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Guo, X.X.; Chen, S.X.; Alvarez, K.M.; Bell, M.W.; Anderson, R.E. Regulation of type II phosphatidylinositol phosphate kinase by tyrosine phosphorylation in bovine rod outer segments. Biochemistry 2001, 40, 4550–4559. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Ma, Z.; Jiao, X.; Fariss, R.; Kantorow, W.L.; Kantorow, M.; Pras, E.; Frydman, M.; Pras, E.; Riazuddin, S.; et al. Mutations in FYCO1 cause autosomal-recessive congenital cataracts. Am. J. Hum. Genet. 2011, 88, 827–838. [Google Scholar] [CrossRef] [PubMed]
- Hong, Z.; Pedersen, N.M.; Wang, L.; Torgersen, M.L.; Stenmark, H.; Raiborg, C. PtdIns3P controls mTORC1 signaling through lysosomal positioning. J. Cell Biol. 2017, 216, 4217–4233. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Gonzalo, F.R.; Phua, S.C.; Roberson, E.C.; Garcia, G., 3rd; Abedin, M.; Schurmans, S.; Inoue, T.; Reiter, J.F. Phosphoinositides Regulate Ciliary Protein Trafficking to Modulate Hedgehog Signaling. Dev. Cell 2015, 34, 400–409. [Google Scholar] [CrossRef] [PubMed]
- Nakatsu, F. A Phosphoinositide Code for Primary Cilia. Dev. Cell 2015, 34, 379–380. [Google Scholar] [CrossRef] [PubMed]
- Luo, N.; Lu, J.; Sun, Y. Evidence of a role of inositol polyphosphate 5-phosphatase INPP5E in cilia formation in zebrafish. Vis. Res. 2012, 75, 98–107. [Google Scholar] [CrossRef]
- Lee, J.E.; Gleeson, J.G. Cilia in the nervous system: Linking cilia function and neurodevelopmental disorders. Curr. Opin. Neurol. 2011, 24, 98–105. [Google Scholar] [CrossRef]
- Berman, E.R.; Feeney, L. Letter: Clean start for the retinal pigment epithelium. Investig. Ophthalmol. 1976, 15, 238–240. [Google Scholar]
- Rothman, H.; Feeney, L.; Berman, E.R. The retinal pigment epithelium. Analytical subcellular fractionation with special reference to acid lipase. Exp. Eye Res. 1976, 22, 519–532. [Google Scholar] [CrossRef]
- Rouser, G.; Siakotos, A.N.; Fleischer, S. Quantitative analysis of phospholipids by thin-layer chromatography and phosphorus analysis of spots. Lipids 1966, 1, 85–86. [Google Scholar] [CrossRef]
- Gortner, W.A. An evaluation of micromethods for phospholipids. J. Biol. Chem. 1945, 159, 97–100. [Google Scholar]
- Furutani, M.; Tsujita, K.; Itoh, T.; Ijuin, T.; Takenawa, T. Application of phosphoinositide-binding domains for the detection and quantification of specific phosphoinositides. Anal. Biochem. 2006, 355, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Oikawa, T.; Yamaguchi, H.; Itoh, T.; Kato, M.; Ijuin, T.; Yamazaki, D.; Suetsugu, S.; Takenawa, T. PtdIns(3,4,5)P3 binding is necessary for WAVE2-induced formation of lamellipodia. Nat. Cell Biol. 2004, 6, 420–426. [Google Scholar] [CrossRef]
- Lebendiker, M.; Danieli, T. Purification of proteins fused to maltose-binding protein. Methods Mol. Biol. 2011, 681, 281–293. [Google Scholar] [CrossRef]
- Dowler, S.; Kular, G.; Alessi, D.R. Protein lipid overlay assay. Sci. STKE: Signal Transduct. Knowl. Environ. 2002, 2002, pl6. [Google Scholar] [CrossRef] [PubMed]
- Rameh, L.E.; Cantley, L.C. The role of phosphoinositide 3-kinase lipid products in cell function. J. Biol. Chem. 1999, 274, 8347–8350. [Google Scholar] [CrossRef]
- Ghalayini, A.J.; Anderson, R.E. Light adaptation of bovine retinas in situ stimulates phosphatidylinositol synthesis in rod outer segments in vitro. Curr. Eye Res. 1995, 14, 1025–1029. [Google Scholar] [CrossRef]
- Huang, Z.; Ghalayini, A.; Guo, X.X.; Alvarez, K.M.; Anderson, R.E. Light-mediated activation of diacylglycerol kinase in rat and bovine rod outer segments. J. Neurochem. 2000, 75, 355–362. [Google Scholar] [CrossRef]
- Guo, X.; Ghalayini, A.J.; Chen, H.; Anderson, R.E. Phosphatidylinositol 3-kinase in bovine photoreceptor rod outer segments. Investig. Ophthalmol. Vis. Sci. 1997, 38, 1873–1882. [Google Scholar]
- Guo, X.X.; Huang, Z.; Bell, M.W.; Chen, H.; Anderson, R.E. Tyrosine phosphorylation is involved in phosphatidylinositol 3-kinase activation in bovine rod outer segments. Mol. Vis. 2000, 6, 216–221. [Google Scholar]
- Hazlett, L.; Suvas, S.; McClellan, S.; Ekanayaka, S. Challenges of corneal infections. Expert Rev. Ophthalmol. 2016, 11, 285–297. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Rajala, A.; Wiechmann, A.F.; Anderson, R.E.; Rajala, R.V. Activation and membrane binding of retinal protein kinase Balpha/Akt1 is regulated through light-dependent generation of phosphoinositides. J. Neurochem. 2008, 107, 1382–1397. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.W.; Kang, K.H.; Burrola, P.; Mak, T.W.; Lemke, G. Retinal degeneration triggered by inactivation of PTEN in the retinal pigment epithelium. Genes Dev. 2008, 22, 3147–3157. [Google Scholar] [CrossRef] [PubMed]
- Venkatesh, A.; Ma, S.; Le, Y.Z.; Hall, M.N.; Ruegg, M.A.; Punzo, C. Activated mTORC1 promotes long-term cone survival in retinitis pigmentosa mice. J. Clin. Investig. 2015, 125, 1446–1458. [Google Scholar] [CrossRef]
- Rajala, R.V.; Anderson, R.E. Interaction of the insulin receptor beta-subunit with phosphatidylinositol 3-kinase in bovine ROS. Investig. Ophthalmol. Vis. Sci. 2001, 42, 3110–3117. [Google Scholar]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rajala, A.; McCauley, A.; Brush, R.S.; Nguyen, K.; Rajala, R.V.S. Phosphoinositide Lipids in Ocular Tissues. Biology 2020, 9, 125. https://doi.org/10.3390/biology9060125
Rajala A, McCauley A, Brush RS, Nguyen K, Rajala RVS. Phosphoinositide Lipids in Ocular Tissues. Biology. 2020; 9(6):125. https://doi.org/10.3390/biology9060125
Chicago/Turabian StyleRajala, Ammaji, Austin McCauley, Richard S. Brush, Khuong Nguyen, and Raju V.S. Rajala. 2020. "Phosphoinositide Lipids in Ocular Tissues" Biology 9, no. 6: 125. https://doi.org/10.3390/biology9060125
APA StyleRajala, A., McCauley, A., Brush, R. S., Nguyen, K., & Rajala, R. V. S. (2020). Phosphoinositide Lipids in Ocular Tissues. Biology, 9(6), 125. https://doi.org/10.3390/biology9060125