Centrifugation Force and Time Alter CASA Parameters and Oxidative Status of Cryopreserved Stallion Sperm


 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Sperm Samples and Experimental Design
2.3. Sperm Motility Assessment by Computer-Assisted Sperm Analysis (CASA)
2.4. Mitochondria Respiration Study
2.5. Sperm Staining with MitoTracker Green FM and Laser Scanning Confocal Microscopy (LSCM)-based Assessment of Mitochondrial Activity
2.6. Carbonylated Protein Assessment by Western Blot
2.7. Terminal Deoxynucleotidyl Transferase-mediated dUTP Nick-End Labeling (TUNEL) Assay for DNA Fragmentation Assessment
2.8. Statistical Analysis
3. Results
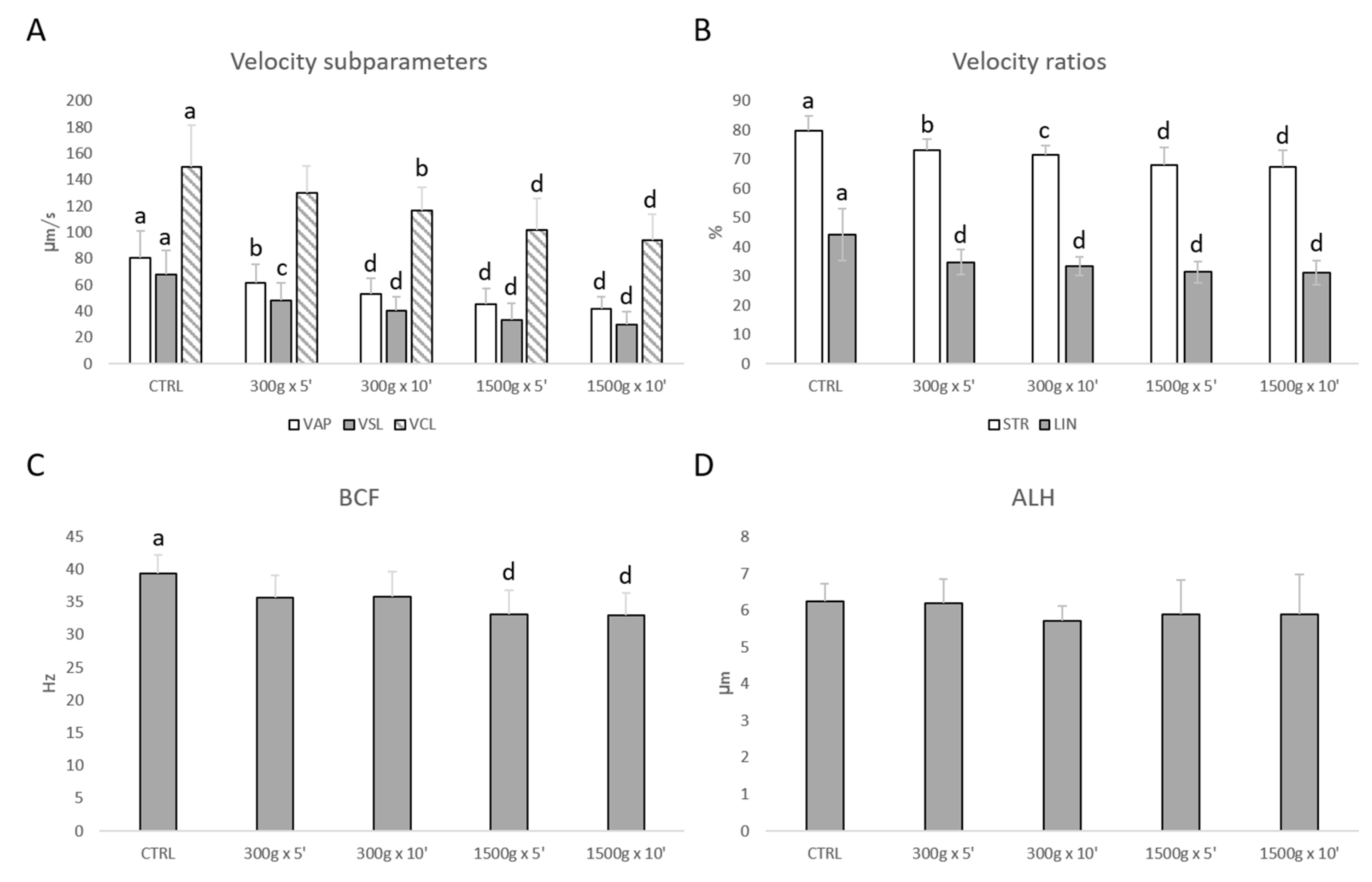
3.1. Effects of Different Centrifugation Protocols on Sperm
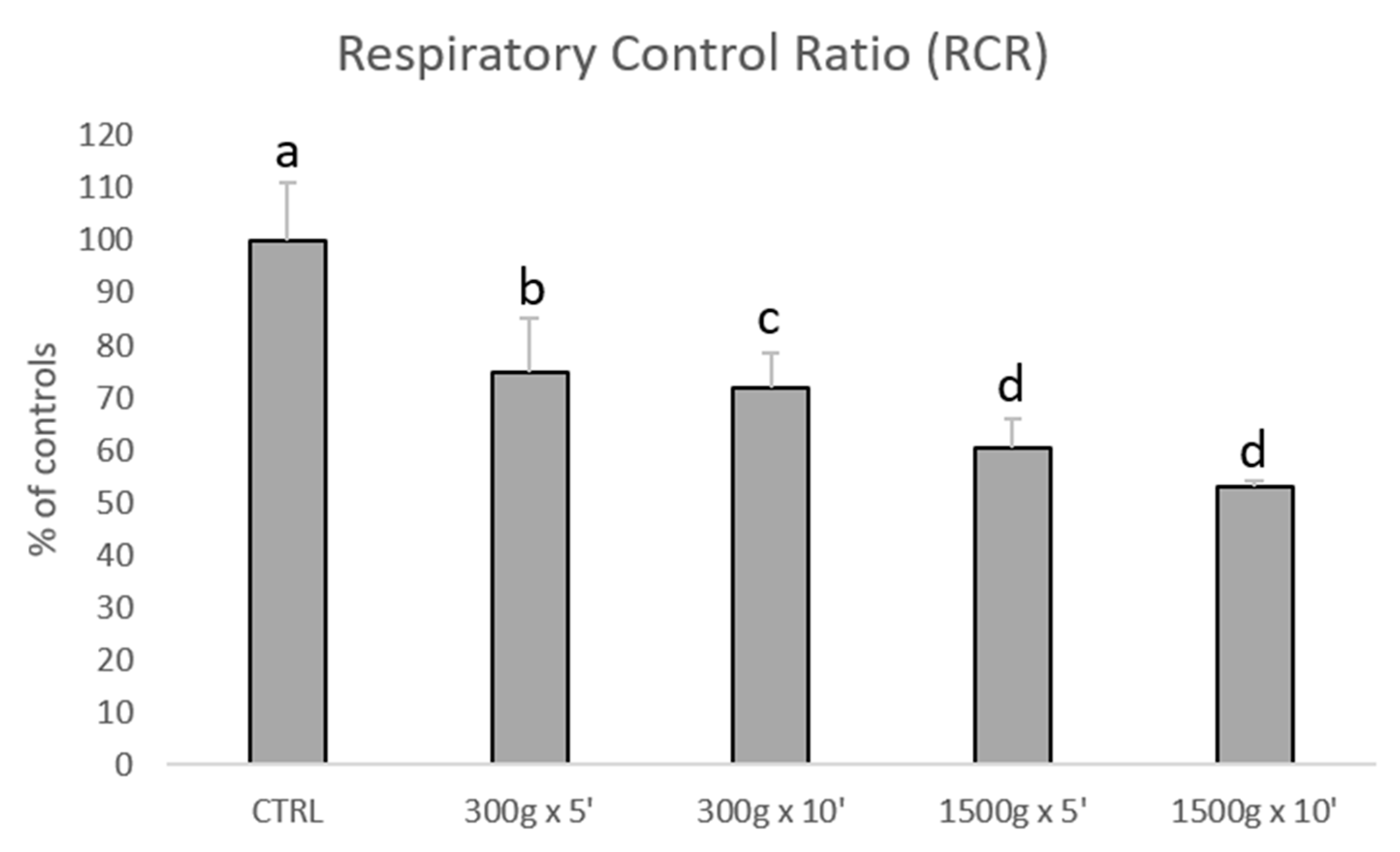
3.2. Effects of Different Centrifugation Protocols on Sperm Mitochondrial Respiration
3.3. Effects of Different Centrifugation Protocols on Sperm Mitochondrial Membrane Potential
3.4. Effect of Different Centrifugation Protocols on Sperm Oxidative Damages
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Henkel, R.R.; Schill, W.B. Sperm Preparation for Art. Reprod. Biol. Endocrinol. 2003, 1, 108. [Google Scholar] [CrossRef] [PubMed]
- WHO. Laboratory Manual Examination and Processing of Human Semen. 2010. Available online: https://www.who.int/reproductivehealth/publications/infertility/9789241547789/en/ (accessed on 10 October 2019).
- Verma, O.P.; Kumar, R.; Kumar, A.; Chand, S. Assisted Repproductive Techniques in Farm Animal - from Artificial Insemination to Nanobiotechnology. Vet. World 2012, 5, 301–310. [Google Scholar] [CrossRef]
- Li, Z.; Zhou, Y.; Liu, R.; Lin, H.; Lin, W.; Xiao, W.; Lin, Q. Effects of Semen Processing on the Generation of Reactive Oxygen Species and Mitochondrial Membrane Potential of Human Spermatozoa. Andrologia 2012, 44, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, J.G.; Lasso, J.L.; Blasco, L.; Nunez, R.C.; Heyner, S.; Caballero, P.P.; Storey, B.T. Centrifugation of Human Spermatozoa Induces Sublethal Damage; Separation of Human Spermatozoa from Seminal Plasma by a Dextran Swim-up Procedure without Centrifugation Extends Their Motile Lifetime. Hum. Reprod. 1993, 8, 1087–1092. [Google Scholar] [CrossRef]
- Zhu, W.J. Preparation and Observation Methods Can Produce Misleading Artefacts in Human Sperm Ultrastructural Morphology. Andrologia 2018, 50. [Google Scholar] [CrossRef]
- Ghaleno, L.R.; Valojerdi, M.R.; Janzamin, E.; Chehrazi, M.; Sharbatoghli, M.; Yazdi, R.S. Evaluation of Conventional Semen Parameters, Intracellular Reactive Oxygen Species, DNA Fragmentation and Dysfunction of Mitochondrial Membrane Potential after Semen Preparation Techniques: A Flow Cytometric Study. Arch. Gynecol. Obs. 2014, 289, 173–180. [Google Scholar] [CrossRef]
- Muratori, M.; Tarozzi, N.; Cambi, M.; Boni, L.; Iorio, A.L.; Passaro, C.; Luppino, B.; Nadalini, M.; Marchiani, S.; Tamburrino, L.; et al. Variation of DNA Fragmentation Levels During Density Gradient Sperm Selection for Assisted Reproduction Techniques: A Possible New Male Predictive Parameter of Pregnancy? Medicine (Baltim.) 2016, 95, e3624. [Google Scholar] [CrossRef]
- Oliveira, L.Z.; Arruda, R.P.; Celeghini, E.C.C.; de Andrade, A.F.C.; Perini, A.P.; Resende, M.V.; Miguel, M.C.V.; Lucio, A.C.; Hossepian de Lima, V.F.M. Effects of Discontinuous Percoll Gradient Centrifugation on the Quality of Bovine Spermatozoa Evaluated with Computer-Assisted Semen Analysis and Fluorescent Probes Association. Andrologia 2012, 44, 9–15. [Google Scholar] [CrossRef]
- Varisli, O.; Uguz, C.; Agca, C.; Agca, Y. Various Physical Stress Factors on Rat Sperm Motility, Integrity of Acrosome, and Plasma Membrane. J. Androl. 2009, 30, 75–86. [Google Scholar] [CrossRef]
- Kim, S.; Agca, C.; Agca, Y. Effects of Various Physical Stress Factors on Mitochondrial Function and Reactive Oxygen Species in Rat Spermatozoa. Reprod. Fertil. Dev. 2013, 25, 1051–1064. [Google Scholar] [CrossRef]
- Ambruosi, B.; Lacalandra, G.M.; Iorga, A.I.; De Santis, T.; Mugnier, S.; Matarrese, R.; Goudet, G.; Dell’aquila, M.E. Cytoplasmic Lipid Droplets and Mitochondrial Distribution in Equine Oocytes: Implications on Oocyte Maturation, Fertilization and Developmental Competence after Icsi. Theriogenology 2009, 71, 1093–1104. [Google Scholar] [CrossRef] [PubMed]
- Hinrichs, K. Assisted Reproductive Techniques in Mares. Reprod. Domest. Anim. 2018, 53, 4–13. [Google Scholar] [CrossRef] [PubMed]
- Leemans, B.; Stout, T.A.E.; De Schauwer, C.; Heras, S.; Nelis, H.; Hoogewijs, M.; Van Soom, A.; Gadella, B.M. Update on Mammalian Sperm Capacitation: How Much Does the Horse Differ from Other Species? Reproduction 2019, 157, R181. [Google Scholar] [CrossRef] [PubMed]
- England, G. Fertility and Obstetrics in the Horse; John Wiley Sons: Hoboken, NJ, USA, 2008; pp. 200–211. [Google Scholar]
- McKinnon, A.O.; Squires, E.L.; Vaala, W.E.; Varner, D.D. Equine Reproduction, 2nd ed.; Wiley-Blackwell: Oxford, UK, 2011; pp. 867–1571. [Google Scholar]
- Neuhauser, S.; Gosele, P.; Handler, J. Postthaw Addition of Autologous Seminal Plasma Improves Sperm Motion Characteristics in Fair and Poor Freezer Stallions. J. Equine Vet. Sci. 2019, 72, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Pena, F.J.; Garcia, B.M.; Samper, J.C.; Aparicio, I.M.; Tapia, J.A.; Ferrusola, C.O. Dissecting the Molecular Damage to Stallion Spermatozoa: The Way to Improve Current Cryopreservation Protocols? Theriogenology 2011, 76, 1177–1186. [Google Scholar] [CrossRef]
- Gibb, Z.; Aitken, R.J. Recent Developments in Stallion Semen Preservation. J. Equine Vet. Sci. 2016, 43, S29–S36. [Google Scholar] [CrossRef]
- Sieme, H.; Harrison, R.A.; Petrunkina, A.M. Cryobiological Determinants of Frozen Semen Quality, with Special Reference to Stallion. Anim. Reprod. Sci. 2008, 107, 276–292. [Google Scholar] [CrossRef]
- Loomis, P.R.; Graham, J.K. Commercial Semen Freezing: Individual Male Variation in Cryosurvival and the Response of Stallion Sperm to Customized Freezing Protocols. Anim. Reprod. Sci. 2008, 105, 119–128. [Google Scholar] [CrossRef]
- Papas, M.; Catalan, J.; Fernandez-Fuertes, B.; Arroyo, L.; Bassols, A.; Miro, J.; Yeste, M. Specific Activity of Superoxide Dismutase in Stallion Seminal Plasma Is Related to Sperm Cryotolerance. Antioxidants (Basel) 2019, 8, 539. [Google Scholar] [CrossRef]
- Yeste, M.; Estrada, E.; Rocha, L.G.; Marin, H.; Rodriguez-Gil, J.E.; Miro, J. Cryotolerance of Stallion Spermatozoa Is Related to Ros Production and Mitochondrial Membrane Potential Rather Than to the Integrity of Sperm Nucleus. Andrology 2015, 3, 395–407. [Google Scholar] [CrossRef]
- Pickett, B.W.; Sullivan, J.J.; Byers, W.W.; Pace, M.M.; Remmenga, E.E. Effect of Centrifugation and Seminal Plasma on Motility and Fertility of Stallion and Bull Spermatozoa. Fertil. Steril. 1975, 26, 167–174. [Google Scholar] [CrossRef]
- Jasko, D.J.; Moran, D.M.; Farlin, M.E.; Squires, E.L. Effect of Seminal Plasma Dilution or Removal on Spermatozoal Motion Characteristics of Cooled Stallion Semen. Theriogenology 1991, 35, 1059–1067. [Google Scholar] [CrossRef]
- Brinsko, S.P.; Crockett, E.C.; Squires, E.L. Effect of Centrifugation and Partial Removal of Seminal Plasma on Equine Spermatozoal Motility after Cooling and Storage. Theriogenology 2000, 54, 129–136. [Google Scholar] [CrossRef]
- Kareskoski, A.M.; Reilas, T.; Andersson, M.; Katila, T. Motility and Plasma Membrane Integrity of Spermatozoa in Fractionated Stallion Ejaculates after Storage. Reprod. Domest. Anim. 2006, 41, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Maarten, H.; Rijsselaere, T.; de Vliegher, S.; Vanhaesebrouck, E.; de Schauwer, C.; Jan Govaere Thys, M.; Hoflack, G.; van Soom, A.; de Kruif, A. Influence of Different Centrifugation Protocols on Equine Semen Preservation. Theriogenology 2010, 74, 118–126. [Google Scholar]
- Len, J.A.; Jenkins, J.A.; Eilts, B.E.; Paccamonti, D.L.; Lyle, S.K.; Hosgood, G. Immediate and Delayed (after Cooling) Effects of Centrifugation on Equine Sperm. Theriogenology 2010, 73, 225–231. [Google Scholar] [CrossRef]
- Len, J.A.; Beehan, D.P.; Lyle, S.K.; Eilts, B.E. Cushioned Versus Noncushioned Centrifugation: Sperm Recovery Rate and Integrity. Theriogenology 2013, 80, 648–653. [Google Scholar] [CrossRef]
- Piomboni, P.; Focarelli, R.; Stendardi, A.; Ferramosca, A.; Zara, V. The Role of Mitochondria in Energy Production for Human Sperm Motility. Int. J. Androl. 2012, 35, 109–124. [Google Scholar] [CrossRef]
- Ferramosca, A.; Zara, V. Bioenergetics of Mammalian Sperm Capacitation. Biomed. Res. Int. 2014, 2014. [Google Scholar] [CrossRef]
- Moscatelli, N.; Spagnolo, B.; Pisanello, M.; Lemma, E.D.; de Vittorio, M.; Zara, V.; Pisanello, F.; Ferramosca, A. Single-Cell-Based Evaluation of Sperm Progressive Motility Via Fluorescent Assessment of Mitochondria Membrane Potential. Sci. Rep. 2017, 7, 10. [Google Scholar] [CrossRef]
- Natalina, M.; Lunetti, P.; Braccia, C.; Armirotti, A.; Pisanello, F.; de Vittorio, M.; Zara, V.; Ferramosca, A. Comparative Proteomic Analysis of Proteins Involved in Bioenergetics Pathways Associated with Human Sperm Motility. Int. J. Mol. Sci. 2019, 20, 3000. [Google Scholar]
- Saraste, M. Oxidative Phosphorylation at the Fin De Siecle. Science 1999, 283, 1488–1493. [Google Scholar] [CrossRef]
- Gibb, Z.; Sarah, R.L.; Robert, J.A. The Paradoxical Relationship between Stallion Fertility and Oxidative Stress1. Biol. Reprod. 2014, 91. [Google Scholar] [CrossRef]
- Moraes, C.R.; Meyers, S. The Sperm Mitochondrion: Organelle of Many Functions. Anim. Reprod. Sci. 2018, 194, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Pena, F.J.; O’Flaherty, C.; Rodriguez, J.M.O.; Cano, F.E.M.; Gaitskell-Phillips, G.L.; Gil, M.C.; Ferrusola, C.O. Redox Regulation and Oxidative Stress: The Particular Case of the Stallion Spermatozoa. Antioxidants (Basel) 2019, 8, 567. [Google Scholar] [CrossRef] [PubMed]
- Alessandra, F.; Provenzano, S.P.; Montagna, D.D.; Coppola, L.; Zara, V. Oxidative Stress Negatively Affects Human Sperm Mitochondrial Respiration. Urology 2013, 82, 78–83. [Google Scholar]
- Aitken, R.J.; Gibb, Z.; Baker, M.A.; Drevet, J.; Gharagozloo, P. Causes and Consequences of Oxidative Stress in Spermatozoa. Reprod. Fertil. Dev. 2016, 28, 1–10. [Google Scholar] [CrossRef]
- Lone, S.A.; Mohanty, T.K.; Baithalu, R.K.; Yadav, H.P. Sperm Protein Carbonylation. Andrologia 2019, 51, e13233. [Google Scholar] [CrossRef]
- Yves, M.; Dale, B.; Cohen, M. DNA Damage and Repair in Human Oocytes and Embryos: A Review. Zygote 2010, 18, 357–365. [Google Scholar]
- Choi, Y.H.; Chung, Y.G.; Walker, S.C.; Westhusin, M.E.; Hinrichs, K. In Vitro Development of Equine Nuclear Transfer Embryos: Effects of Oocyte Maturation Media and Amino Acid Composition During Embryo Culture. Zygote 2003, 11, 77–86. [Google Scholar] [CrossRef]
- Minervini, F.; Lacalandra, G.M.; Filannino, A.; Garbetta, A.; Nicassio, M.; Dell’Aquila, M.E.; Visconti, A. Toxic Effects Induced by Mycotoxin Fumonisin B1 on Equine Spermatozoa: Assessment of Viability, Sperm Chromatin Structure Stability, Ros Production and Motility. Toxicology 2010, 24, 2072–2078. [Google Scholar] [CrossRef] [PubMed]
- Ferramosca, A.; Focarelli, R.; Piomboni, P.; Coppola, L.; Zara, V. Oxygen Uptake by Mitochondria in Demembranated Human Spermatozoa: A Reliable Tool for the Evaluation of Sperm Respiratory Efficiency. Int. J. Androl. 2008, 31, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Amaral, A.; Ramalho-Santos, J. Assessment of Mitochondrial Potential: Implications for the Correct Monitoring of Human Sperm Function. Int. J. Androl. 2010, 33, E180–E186. [Google Scholar] [CrossRef] [PubMed]
- De Riccardis, L.; Rizzello, A.; Ferramosca, A.; Urso, E.; de Robertis, F.; Danieli, A.; Giudetti, A.M.; Trianni, G.; Zara, V.; Maffia, M. Bioenergetics Profile of Cd4(+) T Cells in Relapsing Remitting Multiple Sclerosis Subjects. J. Biotechnol. 2015, 202, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Giuseppina, M.; Chiriacò, M.S.; Primiceri, E.; Dell’Aquila, M.E.; Ramalho-Santos, J.; Zara, V.; Ferramosca, A.; Maruccio, G. Sperm Selection in Assisted Reproduction: A Review of Established Methods and Cutting-Edge Possibilities. Biotechnol. Adv. 2019. [Google Scholar] [CrossRef]
- Marzano, G.; Mastrorocco, A.; Zianni, R.; Mangiacotti, M.; Chiaravalle, A.E.; Lacalandra, G.M.; Minervini, F.; Cardinali, A.; Macciocca, M.; Vicenti, R.; et al. Altered Morphokinetics in Equine Embryos from Oocytes Exposed to Dehp During Ivm. Mol. Reprod. Dev. 2019, 86, 1388–1404. [Google Scholar] [CrossRef]
- Hammerstedt, R.H.; Graham, J.K.; Nolan, J.P. Cryopreservation of Mammalian Sperm: What We Ask Them to Survive. J. Androl. 1990, 11, 73–88. [Google Scholar]
- Darr, C.R.; Cortopassi, G.A.; Datta, S.; Varner, D.D.; Meyers, S.A. Mitochondrial Oxygen Consumption Is a Unique Indicator of Stallion Spermatozoal Health and Varies with Cryopreservation Media. Theriogenology 2016, 86, 1382–1392. [Google Scholar] [CrossRef]
- Darr, C.R.; Moraes, L.E.; Scanlan, T.N.; Baumber-Skaife, J.; Loomis, P.R.; Cortopassi, G.A.; Meyers, S.A. Sperm Mitochondrial Function Is Affected by Stallion Age and Predicts Post-Thaw Motility. J. Equine Vet. Sci. 2017, 50, 52–61. [Google Scholar] [CrossRef]
- Magdanz, V.; Boryshpolets, S.; Ridzewski, C.; Eckel, B.; Reinhardt, K. The Motility-Based Swim-up Technique Separates Bull Sperm Based on Differences in Metabolic Rates and Tail Length. PLoS ONE 2019, 14. [Google Scholar] [CrossRef]
- Calvert, S.J.; Reynolds, S.; Paley, M.N.; Walters, S.J.; Pacey, A.A. Probing Human Sperm Metabolism Using 13c-Magnetic Resonance Spectroscopy. Mol. Hum. Reprod. 2019, 25, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Gravance, C.G.; Garner, D.L.; Baumber, J.; Ball, B.A. Assessment of Equine Sperm Mitochondrial Function Using Jc-1. Theriogenology 2000, 53, 1691–1703. [Google Scholar] [CrossRef]
- Morrell, J.M.; Lagerqvist, A.; Humblot, P.; Johannisson, A. Effect of Single Layer Centrifugation on Reactive Oxygen Species and Sperm Mitochondrial Membrane Potential in Cooled Stallion Semen. Reprod. Fertil. Dev. 2017, 29, 1039–1045. [Google Scholar] [CrossRef] [PubMed]
- Pena, F.J.; Ball, B.A.; Squires, E.L. A New Method for Evaluating Stallion Sperm Viability and Mitochondrial Membrane Potential in Fixed Semen Samples. Cytom. B Clin. Cytom. 2018, 94, 302–311. [Google Scholar] [CrossRef]
- Jezek, J.; Cooper, K.F.; Strich, R. Reactive Oxygen Species and Mitochondrial Dynamics: The Yin and Yang of Mitochondrial Dysfunction and Cancer Progression. Antioxidants 2018, 7, 13. [Google Scholar] [CrossRef]
- Morte, M.I.; Rodrigues, A.M.; Soares, D.; Rodrigues, A.S.; Gamboa, S.; Ramalho-Santos, J. The Quantification of Lipid and Protein Oxidation in Stallion Spermatozoa and Seminal Plasma: Seasonal Distinctions and Correlations with DNA Strand Breaks, Classical Seminal Parameters and Stallion Fertility. Anim. Reprod. Sci. 2008, 106, 36–47. [Google Scholar] [CrossRef][Green Version]
- Agarwal, A.; Virk, G.; Ong, C.; Plessis, S.S.D. Effect of Oxidative Stress on Male Reproduction. World J. Mens Health 2014, 32, 1–17. [Google Scholar] [CrossRef]
- Katkov, I.I.; Mazur, P. Influence of Centrifugation Regimes on Motility, Yield, and Cell Associations of Mouse Spermatozoa. J. Androl. 1998, 19, 232–241. [Google Scholar]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marzano, G.; Moscatelli, N.; Di Giacomo, M.; Martino, N.A.; Lacalandra, G.M.; Dell’Aquila, M.E.; Maruccio, G.; Primiceri, E.; Chiriacò, M.S.; Zara, V.; et al. Centrifugation Force and Time Alter CASA Parameters and Oxidative Status of Cryopreserved Stallion Sperm. Biology 2020, 9, 22. https://doi.org/10.3390/biology9020022
Marzano G, Moscatelli N, Di Giacomo M, Martino NA, Lacalandra GM, Dell’Aquila ME, Maruccio G, Primiceri E, Chiriacò MS, Zara V, et al. Centrifugation Force and Time Alter CASA Parameters and Oxidative Status of Cryopreserved Stallion Sperm. Biology. 2020; 9(2):22. https://doi.org/10.3390/biology9020022
Chicago/Turabian StyleMarzano, Giuseppina, Natalina Moscatelli, Mariangela Di Giacomo, Nicola Antonio Martino, Giovanni Michele Lacalandra, Maria Elena Dell’Aquila, Giuseppe Maruccio, Elisabetta Primiceri, Maria Serena Chiriacò, Vincenzo Zara, and et al. 2020. "Centrifugation Force and Time Alter CASA Parameters and Oxidative Status of Cryopreserved Stallion Sperm" Biology 9, no. 2: 22. https://doi.org/10.3390/biology9020022
APA StyleMarzano, G., Moscatelli, N., Di Giacomo, M., Martino, N. A., Lacalandra, G. M., Dell’Aquila, M. E., Maruccio, G., Primiceri, E., Chiriacò, M. S., Zara, V., & Ferramosca, A. (2020). Centrifugation Force and Time Alter CASA Parameters and Oxidative Status of Cryopreserved Stallion Sperm. Biology, 9(2), 22. https://doi.org/10.3390/biology9020022