HLA-DRB1*07:01 and *08:02 Alleles Confer a Protective Effect Against ACPA-Positive Rheumatoid Arthritis in a Latin American Admixed Population

 , ,
, , 

Abstract
Simple Summary
Abstract
1. Introduction
2. Patients and Methods
2.1. Study Population
2.2. Characterization of HLA-DRB1 Alleles and Levels of ACPA
2.3. SNP Genotyping
2.4. Statistical Analysis
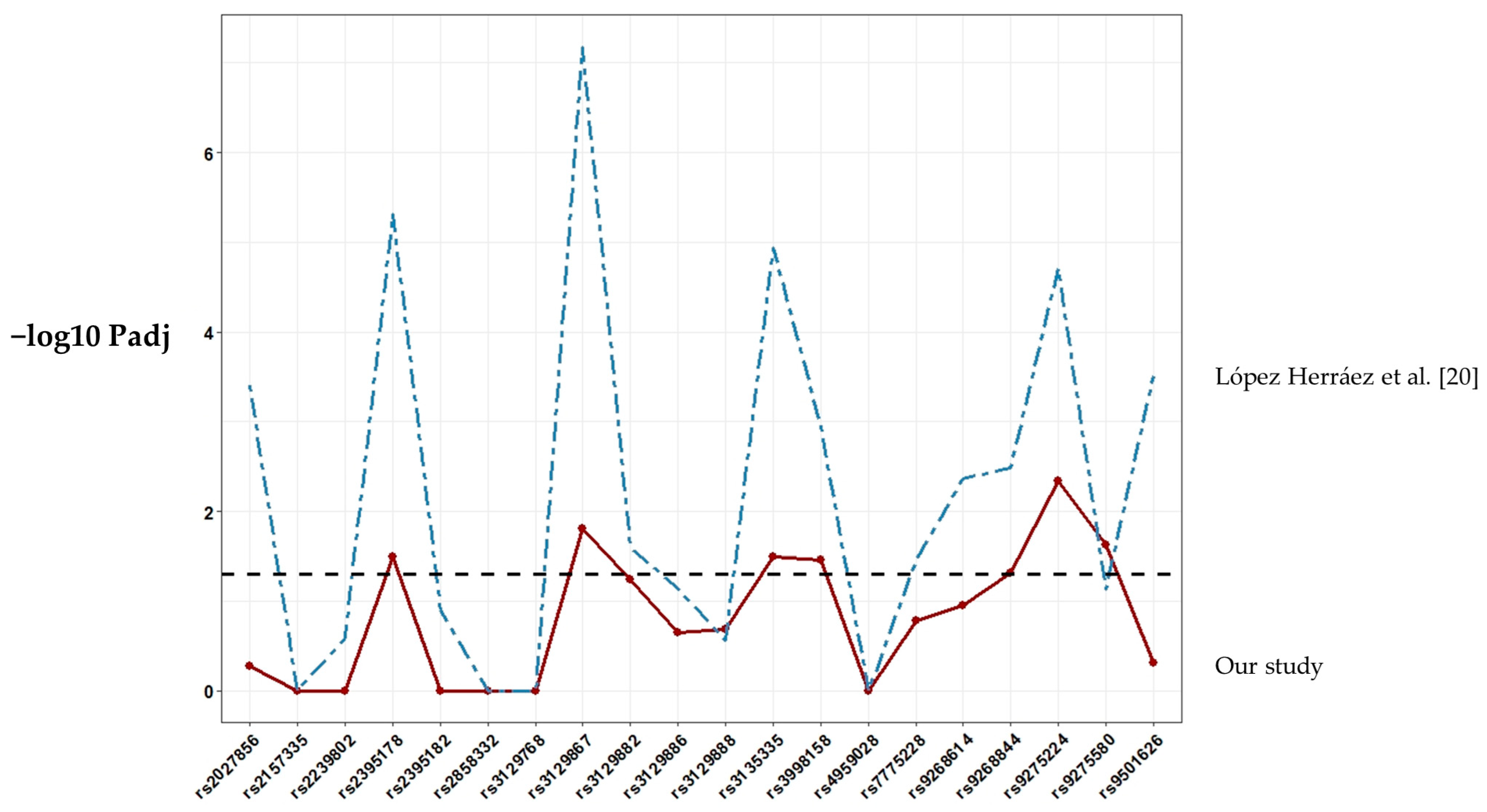
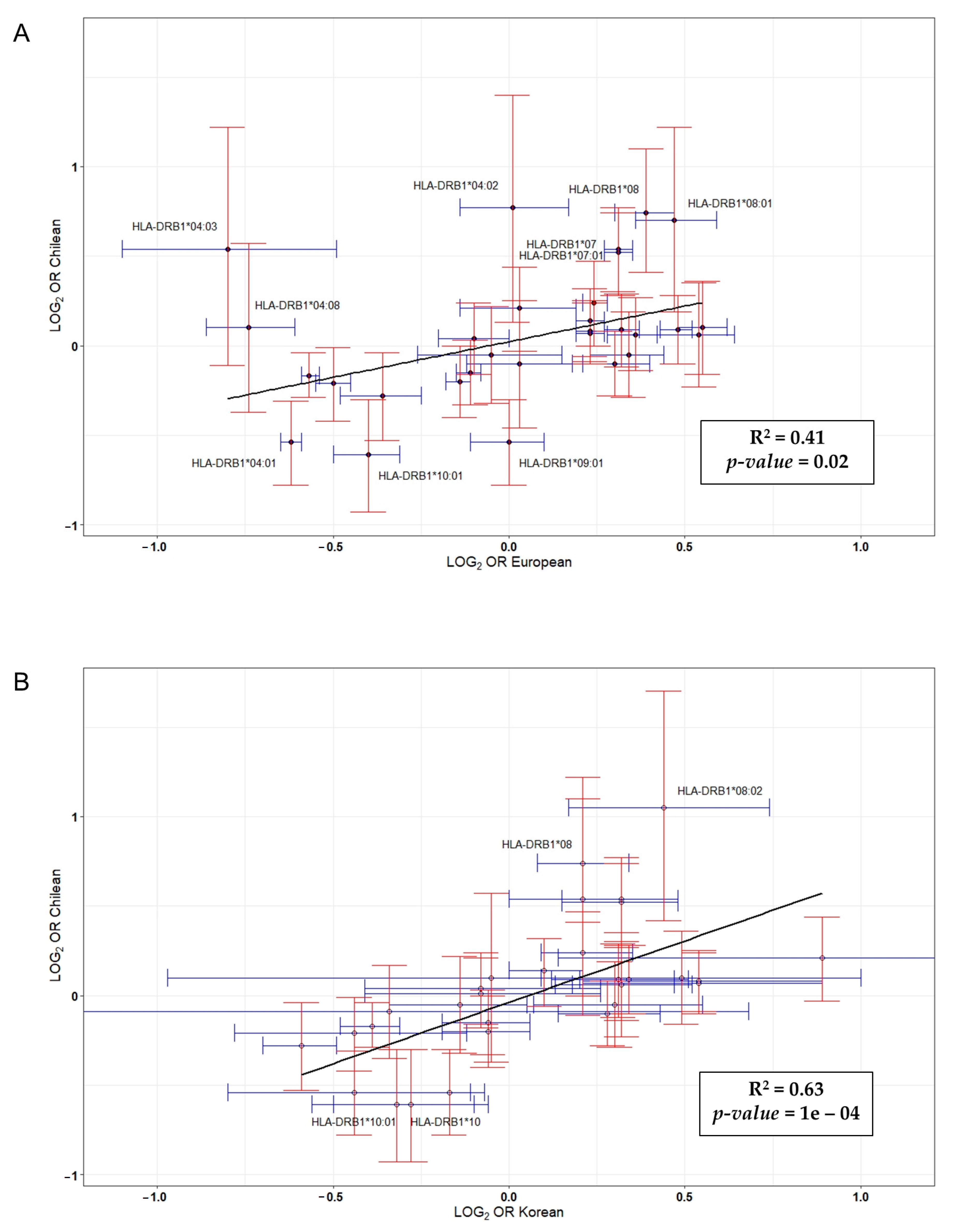
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Tobón, G.J.; Youinou, P.; Saraux, A. The environment, geo-epidemiology, and autoimmune disease: Rheumatoid arthritis. J. Autoimmun. 2010, 35, 10–14. [Google Scholar] [CrossRef] [PubMed]
- Sans, M. Admixture studies in Latin America: From the 20th to the 21st century. Hum. Biol. 2000, 72, 155–177. [Google Scholar] [PubMed]
- Price, A.L.; Patterson, N.; Yu, F.; Cox, D.R.; Waliszewska, A.; McDonald, G.J.; Tandon, A.; Schirmer, C.; Neubauer, J.; Bedoya, G.; et al. A genomewide admixture map for latino populations. Am. J. Hum. Genet. 2007, 80, 1024–1036. [Google Scholar] [CrossRef] [PubMed]
- Eyheramendy, S.; Martinez, F.I.; Manevy, F.; Vial, C.; Repetto, G.M. Genetic structure characterization of Chileans reflects historical immigration patterns. Nat. Commun. 2015, 17, 6472. [Google Scholar] [CrossRef] [PubMed]
- Spindler, A.; Bellomio, V.; Berman, A.; Lucero, E.; Baigorria, M.; Paz, S.; Garrone, N.; Torres, A.I.; Romano, O.; Carraccio, A.; et al. Prevalence of rheumatoid arthritis in Tucuman, Argentina. J. Rheumatol. 2002, 29, 1166–1170. [Google Scholar]
- Rodrigues Senna, É.; De Barros, A.L.P.; Silva, E.O.; Costa, I.F.; Pereira, L.V.B.; Ciconelli, R.M.; Ferraz, M.B. Prevalence of Rheumatic Diseases in Brazil: A Study Using the COPCORD Approach. J. Rheumatol. 2004, 31, 594–597. [Google Scholar]
- Gamboa, R.; Medina, M.; Acevedo, E.; Pastor, C.; Cucho, J.; Gutiérrez, C.; Ugarte, M.; Sánchez, C.; Perich, R.; Alfaro, J.; et al. Prevalencia de enfermedades reumatológicas y discapacidad en una comunidad urbano-marginal: Resultados del primer estudio COPCORD en el Perú. Rev. Peru. Reumatol. 2009, 15, 40–46. [Google Scholar]
- Scublinsky, D.; Venarotti, H.; Citera, G.; Messina, O.D.; Scheines, E.; Rillo, O.; Arturi, A.; Hofman, J.; Somma, L.F.; Casado, G.; et al. The prevalence of rheumatoid arthritis in argentina: A capture-recapture study in a city of Buenos Aires Province. J. Clin. Rheumatol. 2010, 16, 317–321. [Google Scholar] [CrossRef]
- Peláez-Ballestas, I.; Sanin, L.H.; Moreno-Montoya, J.; Alvarez-Nemegyei, J.; Burgos-Vargas, R.; Garza-Elizondo, M.; Rodríguez-Amado, J.; Goycochea-Robles, M.V.; Madariaga, M.; Zamudio, J.; et al. Epidemiology of the rheumatic diseases in Mexico. A study of 5 regions based on the COPCORD methodology. J. Rheumatol. 2011, 86, 3–8. [Google Scholar] [CrossRef]
- Guevara-Pacheco, S.; Feicán-Alvarado, A.; Sanín, L.H.; Vintimilla-Ugalde, J.; Vintimilla-Moscoso, F.; Delgado-Pauta, J.; Lliguisaca-Segarra, A.; Dután-Erráez, H.; Guevara-Mosquera, D.; Ochoa-Robles, V.; et al. Prevalence of musculoskeletal disorders and rheumatic diseases in Cuenca, Ecuador: A WHO-ILAR COPCORD study. Rheumatol. Int. 2016, 36, 1195–1204. [Google Scholar] [CrossRef]
- Granados, Y.; Cedeño, L.; Rosillo, C.; Berbin, S.; Azocar, M.; Molina, M.E.; Lara, O.; Sanchez, G.; Peláez-Ballestas, I. Prevalence of musculoskeletal disorders and rheumatic diseases in an urban community in Monagas State, Venezuela: A COPCORD study. Clin. Rheumatol. 2015, 34, 871–877. [Google Scholar] [CrossRef] [PubMed]
- Durán, J.; Massardo, L.; Llanos, C.; Iacobelli, S.; Burgos, P.I.; Cisternas, M.; Iruretagoyena, M.; Armstrong, M.; Aguilera, R.; Radrigán, F.; et al. The Prevalence of Rheumatoid Arthritis in Chile: A Nationwide Study Performed as Part of the National Health Survey. J. Rheumatol. 2020, 47, 951–958. [Google Scholar] [CrossRef] [PubMed]
- Stastny, P. Association of the B-Cell Alloantigen DRw4 with Rheumatoid Arthritis. N. Engl. J. Med. 1978, 298, 869–871. [Google Scholar] [CrossRef]
- Gregersen, P.K.; Silver, J.; Winchester, R.J. The shared epitope hypothesis. An approach to understanding the molecular genetics of susceptibility to rheumatoid arthritis. Arthritis Rheum. 1987, 30, 1205–1213. [Google Scholar] [CrossRef]
- Aletaha, D.; Neogi, T.; Silman, A.J.; Funovits, J.; Felson, D.T.; Bingham, C.O.; Birnbaum, N.S.; Burmester, G.R.; Bykerk, V.P.; Cohen, M.D.; et al. 2010 Rheumatoid arthritis classification criteria: An American College of Rheumatology/European League Against Rheumatism collaborative initiative. Arthritis Rheum. 2010, 62, 2569–2581. [Google Scholar] [CrossRef]
- Robinson, J.; Halliwell, J.A.; Hayhurst, J.D.; Flicek, P.; Parham, P.; Marsh, S.G.E. The IPD and IMGT/HLA database: Allele variant databases. Nucleic Acids Res. 2015, D423–D431. [Google Scholar] [CrossRef] [PubMed]
- Wysocki, T.; Olesińska, M.; Paradowska-Gorycka, A. Current Understanding of an Emerging Role of HLA-DRB1 Gene in Rheumatoid Arthritis-From Research to Clinical Practice. Cells 2020, 9, 1127. [Google Scholar] [CrossRef]
- Castro-Santos, P.; Díaz-Peña, R. Genetics of rheumatoid arthritis: A new boost is needed in Latin American populations. Rev. Bras. Reumatol. 2016, 56, 171–177. [Google Scholar] [CrossRef]
- Delgado-Vega, A.M.; Anaya, J.-M. Meta-analysis of HLA-DRB1 polymorphism in Latin American patients with rheumatoid arthritis. Autoimmun. Rev. 2007, 6, 402–408. [Google Scholar] [CrossRef]
- Herráez, D.L.; Martínez-Bueno, M.; Riba, L.; De La Torre, I.G.; Sacnún, M.; Goñi, M.; Berbotto, G.A.; Paira, S.; Musuruana, J.L.; Graf, C.E.; et al. Rheumatoid arthritis in latin americans enriched for amerindian ancestry is associated with loci in chromosomes 1, 12, and 13, and the HLA Class II region. Arthritis Rheum. 2013, 65, 1457–1467. [Google Scholar] [CrossRef]
- Okada, Y.; Wu, D.; Trynka, G.; Raj, T.; Terao, C.; Ikari, K.; Kochi, Y.; Ohmura, K.; Suzuki, A.; Yoshida, S.; et al. Genetics of rheumatoid arthritis contributes to biology and drug discovery. Nature 2013, 506, 376–381. [Google Scholar] [CrossRef] [PubMed]
- Terao, C.; Raychaudhuri, S.; Gregersen, P.K. Recent Advances in Defining the Genetic Basis of Rheumatoid Arthritis. Annu. Rev. Genom. Hum. Genet. 2016, 17, 273–301. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Peña, R.; Quiñones, L.A.; Castro-Santos, P.; Durán, J.; Lucia, A. Latin American Genes: The Great Forgotten in Rheumatoid Arthritis. J. Pers. Med. 2020, 10, 196. [Google Scholar] [CrossRef] [PubMed]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef]
- Massardo, L.; Pons-Estel, B.A.; Wojdyla, D.; Cardiel, M.H.; Galarza-Maldonado, C.M.; Sacnun, M.P.; Soriano, E.R.; Laurindo, I.M.; Acevedo-Vásquez, E.M.; Caballero-Uribe, C.V.; et al. Early rheumatoid arthritis in Latin America: Low socioeconomic status related to high disease activity at baseline. Arthritis Care Res. 2012, 64, 1135–1143. [Google Scholar] [CrossRef]
- Schäfer, C.; Sauter, J.; Riethmüller, T.; Kashi, Z.M.; Schmidt, A.H.; Barriga, F.J. HLA-A, -B, -DRB1 allele and haplotype frequencies of 920 cord blood units from Central Chile. Hum. Immunol. 2016, 77, 622–623. [Google Scholar] [CrossRef]
- Rey, D.; Parga-Lozano, C.; Moscoso, J.; Areces, C.; Enriquez-De-Salamanca, M.; Fernández-Honrado, M.; Abd-El-Fatah-Khalil, S.; Alonso-Rubio, J.; Arnaiz-Villena, A. HLA genetic profile of Mapuche (Araucanian) Amerindians from Chile. Mol. Biol. Rep. 2013, 40, 4257–4267. [Google Scholar] [CrossRef]
- Thorsby, E.; Flm, S.T.; Woldseth, B.; Dupuy, B.M.; Sanchez-Mazas, A.; Fernandez-Vina, M.A. Further evidence of an Amerindian contribution to the Polynesian gene pool on Easter Island. Tissue Antigens 2009, 73, 582–585. [Google Scholar] [CrossRef]
- Raychaudhuri, S.; Sandor, C.; Stahl, E.A.; Freudenberg, J.; Lee, H.S.; Jia, X.; Alfredsson, L.; Padyukov, L.; Klareskog, L.; Worthington, J.; et al. Five amino acids in three HLA proteins explain most of the association between MHC and seropositive rheumatoid arthritis. Nat. Genet. 2012, 44, 291–296. [Google Scholar] [CrossRef]
- Okada, Y.; Kim, K.; Han, B.; Pillai, N.E.; Ong, R.T.H.; Saw, W.Y.; Luo, M.; Jiang, L.; Yin, J.; Bang, S.Y.; et al. Risk for ACPA-positive rheumatoid arthritis is driven by shared HLA amino acid polymorphisms in Asian and European populations. Hum. Mol. Genet. 2014, 23, 6916–6926. [Google Scholar] [CrossRef]
- Karami, J.; Aslani, S.; Jamshidi, A.; Garshasbi, M.; Mahmoudi, M. Genetic implications in the pathogenesis of rheumatoid arthritis; an updated review. Gene 2019, 702, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Castro-Santos, P.; Verdugo, R.A.; Alonso-Arias, R.; Gutiérrez, M.A.; Suazo, J.; Aguillón, J.C.; Olloquequi, J.; Pinochet, C.; Lucia, A.; Quiñones, L.A.; et al. Association analysis in a Latin American population revealed ethnic differences in rheumatoid arthritis-associated SNPs in Caucasian and Asian populations. Sci. Rep. 2020, 10, 7879. [Google Scholar] [CrossRef] [PubMed]
- Sinha, A.A. The genetics of pemphigus. Dermatol. Clin. 2011, 29, 381–391. [Google Scholar] [CrossRef]
- De Vries, N.; Tijssen, H.; Van Riel, P.L.C.M.; Van De Putte, L.B.A. Reshaping the shared epitope hypothesis: HLA-associated risk for rheumatoid arthritis is encoded by amino acid substitutions at positions 67–74 of the HLA-DRB1 molecule. Arthritis Rheum. 2002, 46, 921–928. [Google Scholar] [CrossRef] [PubMed]
- Morris, D.L.; Taylor, K.E.; Fernando, M.M.A.; Nititham, J.; Alarcón-Riquelme, M.E.; Barcellos, L.F.; Behrens, T.W.; Cotsapas, C.; Gaffney, P.M.; Graham, R.R.; et al. Unraveling multiple MHC gene associations with systemic lupus erythematosus: Model choice indicates a role for HLA alleles and non-HLA genes in europeans. Am. J. Hum. Genet. 2012, 91, 778–793. [Google Scholar] [CrossRef] [PubMed]
- Flåm, S.T.; Gunnarsson, R.; Garen, T.; Lie, B.A.; Molberg, O.; Lexberg, Å.S.; Time, K.; Dhainaut, A.S.S.; Bertelsen, L.T.; Palm, O.; et al. The HLA profiles of mixed connective tissue disease differ distinctly from the profiles of clinically related connective tissue diseases. Rheumatology (UK) 2015, 54, 528–535. [Google Scholar]
- Furukawa, H.; Oka, S.; Shimada, K.; Sugii, S.; Hashimoto, A.; Komiya, A.; Fukui, N.; Miyashita, T.; Migita, K.; Suda, A.; et al. HLA-DRB1*08:02 is associated with Bucillamine-induced proteinuria in Japanese rheumatoid arthritis patients. Biomark. Insights 2014, 9, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Dilthey, A.T.; Moutsianas, L.; Leslie, S.; McVean, G. HLA*IMP-an integrated framework for imputing classical HLA alleles from SNP genotypes. Bioinformatics 2011, 27, 968–972. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.; Han, B.; Onengut-Gumuscu, S.; Chen, W.M.; Concannon, P.J.; Rich, S.S.; Raychaudhuri, S.; de Bakker, P.I.W. Imputing Amino Acid Polymorphisms in Human Leukocyte Antigens. PLoS ONE 2013, 8, e64683. [Google Scholar] [CrossRef]
- Zheng, X.; Shen, J.; Cox, C.; Wakefield, J.C.; Ehm, M.G.; Nelson, M.R.; Weir, B.S. HIBAG—HLA genotype imputation with attribute bagging. Pharm. J. 2014, 14, 192–200. [Google Scholar] [CrossRef]
- Alarcón-Riquelme, M.E.; Ziegler, J.T.; Molineros, J.; Howard, T.D.; Moreno-Estrada, A.; Sánchez-Rodríguez, E.; Ainsworth, H.C.; Ortiz-Tello, P.; Comeau, M.E.; Rasmussen, A.; et al. Genome-Wide Association Study in an Amerindian Ancestry Population Reveals Novel Systemic Lupus Erythematosus Risk Loci and the Role of European Admixture. Arthritis Rheumatol. 2016, 68, 932–943. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Demographic Characteristics | RA Patients | Healthy Controls |
|---|---|---|
| Women, n (%) | 245 (84.8) | 399 (78.2) |
| Men, n (%) | 44 (15.2) | 111 (21.8) |
| Age, years (median, range) | 48 (20–75) | 42 (25–72) |
| Clinical Parameters | ||
| Disease duration, years (median, range) | 8 (1–42) | - |
| Age of onset, years (median, range) | 40 (15–67) | - |
| HLA-DRB1 SE Alleles | Frequency | |||
|---|---|---|---|---|
| Our cohort (n = 510) | Schäfer C et al. [26] | Rey D et al. [27] | Thorsby E et al. [28] | |
| 01:01 | 0.08 | 0.02 | 0.01 | 0.00 |
| 01:02 | 0.03 | 0.03 | 0.00 | 0.24 |
| 04:01 | 0.04 | 0.01 | 0.00 | 0.05 |
| 04:04 | 0.08 | 0.01 | 0.01 | 0.02 |
| 04:05 | 0.05 | 0.02 | 0.00 | 0.02 |
| 04:08 | 0.02 | 0.01 | 0.00 | 0.00 |
| 10:01 | 0.02 | 0.01 | 0.01 | 0.00 |
| 14:02 | 0.06 | 0.04 | 0.15 | 0.00 |
| HLA-DRB1 Allele | RA Cases (n = 289) | Controls (n = 510) | p-Value | BONF | OR (CI 95%) | ||
|---|---|---|---|---|---|---|---|
| n | % | n | % | ||||
| SE-positive† | 179 | 61.76 | 169 | 33.10 | 2.83 × 10−15 | 1.02 × 10−13 | 2.28 (2.43–4.44) |
| 01:01 | 36 | 12.35 | 42 | 8.16 | NS | NS | - |
| 01:02 | 10 | 3.53 | 14 | 2.82 | NS | NS | - |
| 01:03 | 0 | 0.00 | 4 | 0.73 | NS | NS | - |
| 03:01 | 43 | 14.71 | 87 | 16.97 | NS | NS | - |
| 03:05 | 0 | 0.00 | 2 | 0.36 | NS | NS | - |
| 04:01 | 41 | 14.12 | 23 | 4.44 | 2.18 × 10−6 | 7.85 × 10−5 | 3.5 (2.05–5.97) |
| 04:02 | 2 | 0.59 | 20 | 3.83 | 0.01 | NS | 0.17 (0.04–0.74) |
| 04:03 | 2 | 0.59 | 12 | 2.37 | NS | NS | - |
| 04:04 | 37 | 12.94 | 42 | 8.25 | NS | NS | - |
| 04:05 | 26 | 8.82 | 25 | 4.97 | NS | NS | - |
| 04:07 | 19 | 6.47 | 52 | 10.27 | NS | NS | - |
| 04:08 | 5 | 1.76 | 11 | 2.18 | NS | NS | - |
| 04:10 | 0 | 0.00 | 1 | 0.20 | NS | NS | - |
| 07:01 | 17 | 5.88 | 88 | 17.33 | 7.08 × 10−6 | 2.55 × 10−4 | 0.30 (0.18–0.52) |
| 07:11 | 0 | 0.00 | 2 | 0.36 | NS | NS | - |
| 08:01 | 3 | 1.18 | 26 | 5.14 | 5 × 10−3 | NS | 0.20 (0.06–0.65) |
| 08:02 | 2 | 0.59 | 36 | 7.05 | 4.2 × 10−5 | 1.51 × 10−3 | 0.09 (0.02–0.38) |
| 08:06 | 0 | 0.00 | 1 | 0.20 | NS | NS | - |
| 08:11 | 2 | 0.59 | 0 | 0.00 | NS | NS | - |
| 09:01 | 39 | 13.53 | 22 | 4.28 | 4.44 × 10−6 | 1.60 × 10−4 | 3.46 (2.01–5.96) |
| 10:01 | 24 | 8.24 | 11 | 2.07 | 7.15 × 10−5 | 2.57 × 10−3 | 4.11 (1.98–8.52) |
| 11:01 | 29 | 10.00 | 58 | 11.47 | NS | NS | - |
| 11:04 | 0 | 0.00 | 3 | 0.59 | NS | NS | - |
| 12:01 | 0 | 0.00 | 6 | 1.12 | NS | NS | - |
| 13:01 | 17 | 5.88 | 37 | 7.27 | NS | NS | - |
| 13:02 | 14 | 4.71 | 28 | 5.50 | NS | NS | - |
| 13:03 | 0 | 0.00 | 5 | 0.98 | NS | NS | - |
| 13:05 | 2 | 0.59 | 0 | 0.00 | NS | NS | - |
| 14:01 | 22 | 7.65 | 35 | 6.85 | NS | NS | - |
| 14:02 | 20 | 7.06 | 29 | 5.62 | NS | NS | - |
| 14:13 | 2 | 0.59 | 0 | 0.00 | NS | NS | - |
| 15:01 | 19 | 6.47 | 55 | 10.77 | NS | NS | - |
| 15:02 | 17 | 5.88 | 27 | 5.28 | NS | NS | - |
| 15:03 | 0 | 0.00 | 1 | 0.20 | NS | NS | - |
| 16:01 | 0 | 0.00 | 3 | 0.59 | NS | NS | - |
| 16:02 | 32 | 11.18 | 58 | 11.29 | NS | NS | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castro-Santos, P.; Olloquequi, J.; Verdugo, R.A.; Gutiérrez, M.A.; Pinochet, C.; Quiñones, L.A.; Díaz-Peña, R. HLA-DRB1*07:01 and *08:02 Alleles Confer a Protective Effect Against ACPA-Positive Rheumatoid Arthritis in a Latin American Admixed Population. Biology 2020, 9, 467. https://doi.org/10.3390/biology9120467
Castro-Santos P, Olloquequi J, Verdugo RA, Gutiérrez MA, Pinochet C, Quiñones LA, Díaz-Peña R. HLA-DRB1*07:01 and *08:02 Alleles Confer a Protective Effect Against ACPA-Positive Rheumatoid Arthritis in a Latin American Admixed Population. Biology. 2020; 9(12):467. https://doi.org/10.3390/biology9120467
Chicago/Turabian StyleCastro-Santos, Patricia, Jordi Olloquequi, Ricardo A. Verdugo, Miguel A. Gutiérrez, Carmen Pinochet, Luis A. Quiñones, and Roberto Díaz-Peña. 2020. "HLA-DRB1*07:01 and *08:02 Alleles Confer a Protective Effect Against ACPA-Positive Rheumatoid Arthritis in a Latin American Admixed Population" Biology 9, no. 12: 467. https://doi.org/10.3390/biology9120467
APA StyleCastro-Santos, P., Olloquequi, J., Verdugo, R. A., Gutiérrez, M. A., Pinochet, C., Quiñones, L. A., & Díaz-Peña, R. (2020). HLA-DRB1*07:01 and *08:02 Alleles Confer a Protective Effect Against ACPA-Positive Rheumatoid Arthritis in a Latin American Admixed Population. Biology, 9(12), 467. https://doi.org/10.3390/biology9120467