How Have Leukocyte In Vitro Chemotaxis Assays Shaped Our Ideas about Macrophage Migration?


Abstract
Simple Summary
Abstract
1. Historical Overview
2. Directional Cell Migration
2.1. Types of Directional Cell Migration: Signal-Dependent
- Chemokinesis happens when the detection of a chemoattractant by cells causes morphological changes that lead to increased overall motility but no directionality of the movement. Chemokinesis can be induced either in the presence of a concentration gradient or in uniform concentration of a chemoattractant, and therefore constitutes an important control for chemotaxis studies [15,16];
- Haptotaxis occurs when the chemoattractant gradient is surface bound, for example to the extracellular matrix (ECM) or endothelium [14]. There is an increasing appreciation that leukocyte migration is most likely directed by the mixture of diffused and surface-bound chemoattractants, highlighting the importance of haptotaxis research [14,17];
- Fugetaxis (chemorepulsion) is defined as the active migration of cells away from the source of the chemokinetic agents, which are, in this case, called chemorepulsants [18];
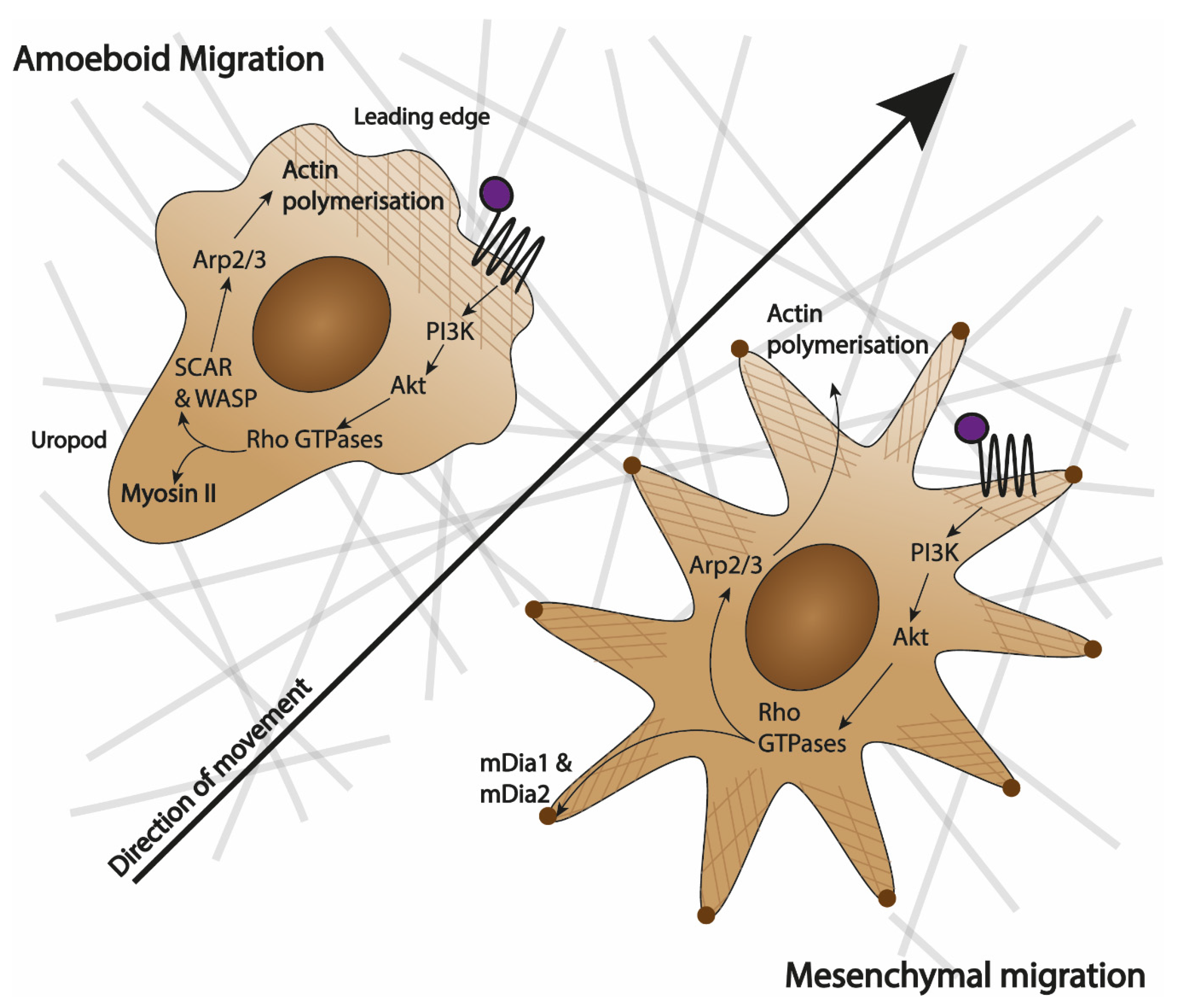
2.2. Types of Specialised Migration: Shape-Dependent
3. Macrophage Chemotaxis
4. Methods to Study Macrophage Chemotaxis In Vitro
4.1. The ‘Ideal Chemotaxis Assay’
- Have no passive movement of cells to ensure that any change in the position of the cells is because of its active motion;
- Ensure that if soluble factors are being tested, they are sufficiently localised in the experimental chamber;
- Be able to control the concentration gradients of chemoattractants from their source until they reach the cells, especially by preventing any convection currents;
- Enable cells to move both towards and away from the chemoattractant source in a homogenous environment;
- Ensure that if a test object is used, it is possible to distinguish between active cell migration and trapping of cells around the given object due to their random movement;
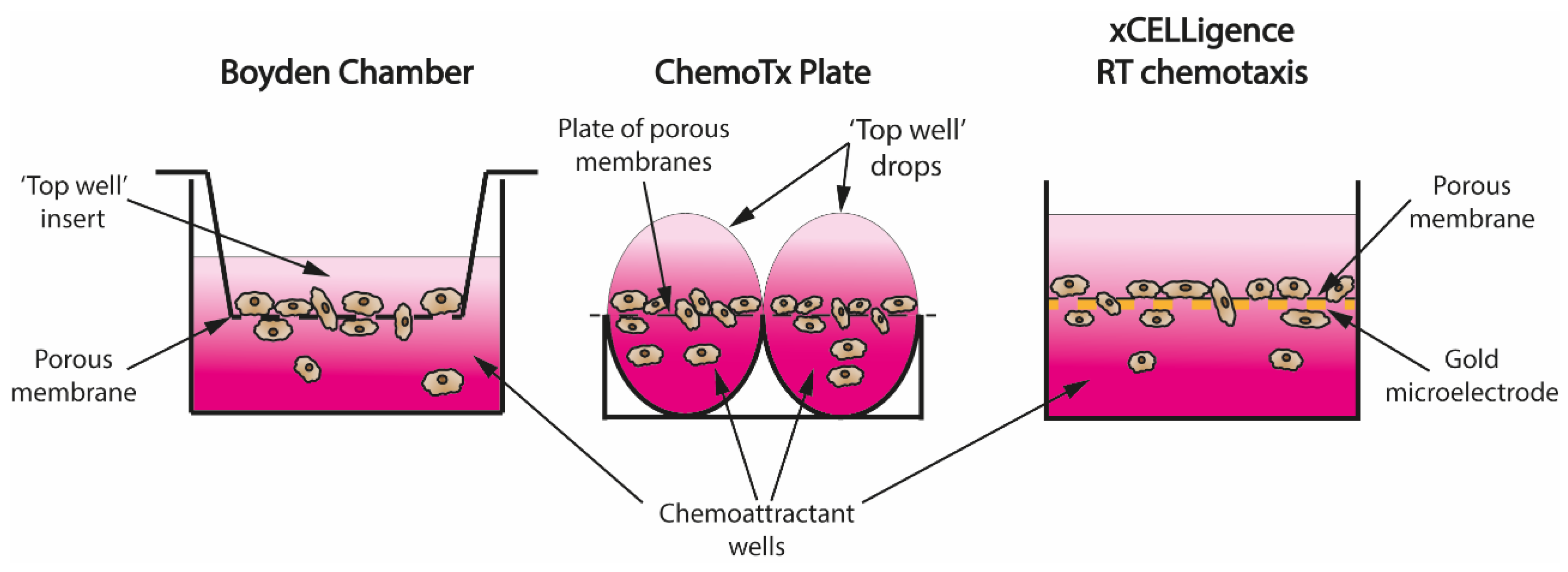
4.2. Transwell Assays—From Boyden Chamber to Real-Time Recordings
4.3. Direct Observation and Cell Tracking Chambers—Zigmond and Dunn Chambers
4.4. ‘Under Agarose’ Migration Assay
4.5. TAXIScan and Ibidi µ-Slides—Movement towards Microfluidics
4.6. Microfluidics Chemotaxis Assays
4.7. Gel Invasion—In Vitro 3D Migration Assay
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ryan, G.B.; Majno, G. Inflammation; The Upjohn Company: Kalamazoo, MI, USA, 1977. [Google Scholar]
- Wilkinson, P.C. Chemotaxis. In Encyclopedia of Immunology; Delves, P.J., Ed.; Elsevier: Amsterdam, The Netherlands, 1998; pp. 533–537. ISBN 9780122267659. [Google Scholar]
- Harris, H. Role of chemotaxis in inflammation. Physiol. Rev. 1954, 34, 529–562. [Google Scholar] [CrossRef]
- Snyderman, R.; Mergenhagen, S.E. Chemotaxis of Macrophages. In Immunobiology of the Macrophage; Nelson, D.S., Ed.; Academic Press, Inc.: New York, NY, USA, 1976; pp. 323–350. [Google Scholar]
- Harris, H. Chemotaxis of granulocytes. J. Pathol. Bacteriol. 1953, 66, 135–146. [Google Scholar] [CrossRef]
- Boyden, S. The chemotactic effect of mixtures of antibody and antigen on polymorphonuclear leucocytes. J. Exp. Med. 1962, 115, 453–466. [Google Scholar] [CrossRef]
- Yang, X.; Dormann, D.; Münsterberg, A.E.; Weijer, C.J. Cell movement patterns during gastrulation in the chick are controlled by positive and negative chemotaxis mediated by FGF4 and FGF8. Dev. Cell 2002, 3, 425–437. [Google Scholar] [CrossRef]
- Chernyavsky, A.I.; Arrendondo, J.; Marubio, L.M.; Grando, S.A. Differential regulation of keratinocyte chemokinesis and chemotaxis through distinct nicotinic receptor subtypes. J. Cell Sci. 2004, 117, 5665–5679. [Google Scholar] [CrossRef]
- Quinn, M.T.; Parthasarathy, S.; Steinberg, D. Lysophosphatidylcholine: A chemotactic factor for human monocytes and its potential role in atherogenesis. Proc. Natl. Acad. Sci. USA 1988, 85, 2805–2809. [Google Scholar] [CrossRef]
- Schneider, L.; Cammer, M.; Lehman, J.; Nielsen, S.K.; Guerra, C.F.; Veland, I.R.; Stock, C.; Hoffmann, E.K.; Yoder, B.K.; Schwab, A.; et al. Directional cell migration and chemotaxis in wound healing response to PDGF-AA are coordinated by the primary cilium in fibroblasts. Cell. Physiol. Biochem. 2010, 25, 279–292. [Google Scholar] [CrossRef]
- Martini, F.J.; Valiente, M.; Bendito, G.L.; Szabó, G.; Moya, F.; Valdeolmillos, M.; Marín, O. Biased selection of leading process branches mediates chemotaxis during tangential neuronal migration. Development 2009, 136, 41–50. [Google Scholar] [CrossRef]
- Vitorino, P.; Meyer, T. Modular control of endothelial sheet migration. Genes Dev. 2008, 22, 3268–3281. [Google Scholar] [CrossRef]
- Bianco, A.; Poukkula, M.; Cliffe, A.; Mathieu, J.; Luque, C.M.; Fulga, T.A.; Rørth, P. Two distinct modes of guidance signalling during collective migration of border cells. Nature 2007, 448, 362–365. [Google Scholar] [CrossRef]
- Rink, I.; Rink, J.; Helmer, D.; Sachs, D.; Schmitz, K. A Haptotaxis Assay for Leukocytes Based on Surface-Bound Chemokine Gradients. J. Immunol. 2015, 194, 5549–5558. [Google Scholar] [CrossRef]
- Boneschansker, L.; Nakayama, H.; Eisenga, M.; Wedel, J.; Klagsbrun, M.; Irimia, D.; Briscoe, D.M. Netrin-1 Augments Chemokinesis in CD4 + T Cells In Vitro and Elicits a Proinflammatory Response In Vivo. J. Immunol. 2016, 197, 1389–1398. [Google Scholar] [CrossRef]
- Gervais, F.G.; Cruz, R.P.G.; Chateauneuf, A.; Gale, S.; Sawyer, N.; Nantel, F.; Metters, K.M.; O’Neill, G.P. Selective modulation of chemokinesis, degranulation, and apoptosis in eosinophils through the PGD2 receptors CRTH2 and DP. J. Allergy Clin. Immunol. 2001, 108, 982–988. [Google Scholar] [CrossRef]
- Schwarz, J.; Bierbaum, V.; Merrin, J.; Frank, T.; Hauschild, R.; Bollenbach, T.; Tay, S.; Sixt, M.; Mehling, M. A microfluidic device for measuring cell migration towards substrate-bound and soluble chemokine gradients. Sci. Rep. 2016, 6, 36440. [Google Scholar] [CrossRef] [PubMed]
- Vianello, F.; Olszak, I.T.; Poznansky, M.C. Fugetaxis: Active movement of leukocytes away from a chemokinetic agent. J. Mol. Med. 2005, 83, 752–763. [Google Scholar] [CrossRef]
- McDonald, B.; Pittman, K.; Menezes, G.B.; Hirota, S.A.; Slaba, I.; Waterhouse, C.C.M.; Beck, P.L.; Muruve, D.A.; Kubes, P. Intravascular danger signals guide neutrophils to sites of sterile inflammation. Science 2010, 330, 362–366. [Google Scholar] [CrossRef]
- Debru, C. A particular form of chemotaxis: Necrotaxis. An historical view. Blood Cells 1993, 19, 5–19, discussion 20–23. [Google Scholar]
- Horwitz, R.; Webb, D. Cell Migration. Curr. Biol. 2003, 13, 756–759. [Google Scholar] [CrossRef]
- Trepat, X.; Chen, Z.; Jacobson, K. Cell migration. Compr. Physiol. 2012, 2, 2369–2392. [Google Scholar] [CrossRef]
- Frank, D.E.; Carter, W.G. Laminin 5 deposition regulates keratinocyte polarization and persistent migration. J. Cell Sci. 2004, 117, 1351–1363. [Google Scholar] [CrossRef]
- Friedl, P.; Weigelin, B. Interstitial leukocyte migration and immune function. Nat. Immunol. 2008, 9, 960–969. [Google Scholar] [CrossRef]
- Wolf, K.; Müller, R.; Borgmann, S.; Bröcker, E.B.; Friedl, P. Amoeboid shape change and contact guidance: T-lymphocyte crawling through fibrillar collagen is independent of matrix remodeling by MMPs and other proteases. Blood 2003, 102, 3262–3269. [Google Scholar] [CrossRef]
- Fritz-Laylin, L.K.; Lord, S.J.; Mullins, R.D. WASP and SCAR are evolutionarily conserved in actin-filled pseudopod-based motility. J. Cell Biol. 2017, 216, 1673–1688. [Google Scholar] [CrossRef]
- Epelman, S.; Lavine, K.J.; Randolph, G.J. Origin and Functions of Tissue Macrophages. Immunity 2014, 41, 21–35. [Google Scholar] [CrossRef]
- Taylor, L.; Brodermann, M.H.; McCaffary, D.; Iqbal, A.J.; Greaves, D.R. Netrin-1 Reduces Monocyte and Macrophage Chemotaxis towards the Complement Component C5a. PLoS ONE 2016, 11, e0160685. [Google Scholar] [CrossRef]
- Cui, K.; Ardell, C.L.; Podolnikova, N.P.; Yakubenko, V.P. Distinct migratory properties of M1, M2, and resident macrophages are regulated by αdβ2 and αmβ2 integrin-mediated adhesion. Front. Immunol. 2018, 9, 2650. [Google Scholar] [CrossRef]
- Barros-Becker, F.; Lam, P.Y.; Fisher, R.; Huttenlocher, A. Live imaging reveals distinct modes of neutrophil and macrophage migration within interstitial tissues. J. Cell Sci. 2017, 130, 3801–3808. [Google Scholar] [CrossRef]
- Grabher, C.; Cliffe, A.; Miura, K.; Hayflick, J.; Pepperkok, R.; Rørth, P.; Wittbrodt, J. Birth and life of tissue macrophages and their migration in embryogenesis and inflammation in medaka. J. Leukoc. Biol. 2007, 81, 263–271. [Google Scholar] [CrossRef]
- Van Goethem, E.; Poincloux, R.; Gauffre, F.; Maridonneau-Parini, I.; Le Cabec, V. Matrix Architecture Dictates Three-Dimensional Migration Modes of Human Macrophages: Differential Involvement of Proteases and Podosome-Like Structures. J. Immunol. 2010, 184, 1049–1061. [Google Scholar] [CrossRef]
- Bignold, L.P. Measurement of chemotaxis of polymorphonuclear leukocytes in vitro. The problems of the control of gradients of chemotactic factors, of the control of the cells and of the separation of chemotaxis from chemokinesis. J. Immunol. Methods 1988, 108, 1–18. [Google Scholar] [CrossRef]
- Frow, E.K.; Reckless, J.; Grainger, D.J. Tools for anti-inflammatory drug design: In vitro models of leukocyte migration. Med. Res. Rev. 2004, 24, 276–298. [Google Scholar] [CrossRef]
- Gee, A.P. Advantages and limitations of methods for measuring cellular chemotaxis and chemokinesis. Mol. Cell. Biochem. 1984, 62, 5–11. [Google Scholar] [CrossRef]
- Falk, W.; Goodwin, R.H.; Leonard, E.J. A 48-well micro chemotaxis assembly for rapid and accurate measurement of leukocyte migration. J. Immunol. Methods 1980, 33, 239–247. [Google Scholar] [CrossRef]
- Iqbal, A.J.; Regan-Komito, D.; Christou, I.; White, G.E.; McNeill, E.; Kenyon, A.; Taylor, L.; Kapellos, T.S.; Fisher, E.A.; Channon, K.M.; et al. A real time chemotaxis assay unveils unique migratory profiles amongst different primary murine macrophages. PLoS ONE 2013, 8, e58744. [Google Scholar] [CrossRef]
- Gruen, M.L.; Hao, M.; Piston, D.W.; Hasty, A.H. Leptin requires canonical migratory signaling pathways for induction of monocyte and macrophage chemotaxis. Am. J. Physiol. Cell Physiol. 2007, 293, 1481–1488. [Google Scholar] [CrossRef]
- Penzo, M.; Habiel, D.M.; Ramadass, M.; Kew, R.R.; Marcu, K.B. Cell migration to CXCL12 requires simultaneous IKKα and IKKβ-dependent NF-κB signaling. Biochim. Biophys. Acta Mol. Cell Res. 2014, 1843, 1796–1804. [Google Scholar] [CrossRef]
- Zigmond, S.H. Ability of polymorphonuclear leukocytes to orient in gradients of chemotactic factors. J. Cell Biol. 1977, 75, 606–616. [Google Scholar] [CrossRef]
- Zicha, D.; Dunn, G.A.; Brown, A.F. A new direct-viewing chemotaxis chamber. J. Cell Sci. 1991, 99, 769–775. [Google Scholar]
- Wheeler, A.P.; Wells, C.M.; Smith, S.D.; Vega, F.M.; Henderson, R.B.; Tybulewicz, V.L.; Ridley, A.J. Rac1 and Rac2 regulate macrophage morphology but are not essential for migration. J. Cell Sci. 2006, 119, 2749–2757. [Google Scholar] [CrossRef]
- Lauffenburger, D.; Rothman, C.; Zigmond, S.H. Measurement of leukocyte motility and chemotaxis parameters with a linear under-agarose migration assay. J. Immunol. 1983, 131, 940–947. [Google Scholar]
- Vogel, D.Y.S.; Heijnen, P.D.A.M.; Breur, M.; de Vries, H.E.; Tool, A.T.J.; Amor, S.; Dijkstra, C.D. Macrophages migrate in an activation-dependent manner to chemokines involved in neuroinflammation. J. Neuroinflammation 2014, 11, 23. [Google Scholar] [CrossRef]
- Zengel, P.; Nguyen-Hoang, A.; Schildhammer, C.; Zantl, R.; Kahl, V.; Horn, E. μ-Slide Chemotaxis: A new chamber for long-term chemotaxis studies. BMC Cell Biol. 2011, 12, 21. [Google Scholar] [CrossRef]
- Berthier, E.; Young, E.W.K.; Beebe, D. Engineers are from PDMS-land, Biologists are from Polystyrenia. Lab Chip 2012, 12, 1224. [Google Scholar] [CrossRef]
- Tweedy, L.; Thomason, P.A.; Paschke, P.I.; Martin, K.; Machesky, L.M.; Zagnoni, M.; Insall, R.H. Seeing around corners: Cells solve mazes and respond at a distance using attractant breakdown. Science 2020, 369. [Google Scholar] [CrossRef]
- Renkawitz, J.; Kopf, A.; Stopp, J.; de Vries, I.; Driscoll, M.K.; Merrin, J.; Hauschild, R.; Welf, E.S.; Danuser, G.; Fiolka, R.; et al. Nuclear positioning facilitates amoeboid migration along the path of least resistance. Nature 2019, 568, 546–550. [Google Scholar] [CrossRef]
- Pakshir, P.; Alizadehgiashi, M.; Wong, B.; Coelho, N.M.; Chen, X.; Gong, Z.; Shenoy, V.B.; McCulloch, C.; Hinz, B. Dynamic fibroblast contractions attract remote macrophages in fibrillar collagen matrix. Nat. Commun. 2019, 10, 1850. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Transwell Assays | Direct Observation and Cell Tracking Chambers | Under Agarose Migration Assay | TAXIScan and Iibidi µ-Slides | Microfluidics | Gel Invasion | |
|---|---|---|---|---|---|---|
| Controlled stable gradient? | + | +/− | − | + | + | − |
| Fugetaxis detection or reversibility? | − | + | + | − | +/− | +/− |
| Distinction between chemotaxis and chemokinesis? | + | + | − | +/− | + | + |
| Single cell tracking? | − | + | − | + | + | + |
| Parallel screening of multiple conditions? | − | − | +/− | + | + | − |
| High throughput? | +/− | − | − | − | +/− | − |
| Real-time recording? | +/− | + | − | + | + | +/− |
| Specialised equipment needed? | +/− | − | − | + | + | − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rumianek, A.N.; Greaves, D.R. How Have Leukocyte In Vitro Chemotaxis Assays Shaped Our Ideas about Macrophage Migration? Biology 2020, 9, 439. https://doi.org/10.3390/biology9120439
Rumianek AN, Greaves DR. How Have Leukocyte In Vitro Chemotaxis Assays Shaped Our Ideas about Macrophage Migration? Biology. 2020; 9(12):439. https://doi.org/10.3390/biology9120439
Chicago/Turabian StyleRumianek, Agata N., and David R. Greaves. 2020. "How Have Leukocyte In Vitro Chemotaxis Assays Shaped Our Ideas about Macrophage Migration?" Biology 9, no. 12: 439. https://doi.org/10.3390/biology9120439
APA StyleRumianek, A. N., & Greaves, D. R. (2020). How Have Leukocyte In Vitro Chemotaxis Assays Shaped Our Ideas about Macrophage Migration? Biology, 9(12), 439. https://doi.org/10.3390/biology9120439