
Quantitative Profiling of Hydroxy Lipid Metabolites in Mouse Organs Reveals Distinct Lipidomic Profiles and Modifications Due to Elevated n-3 Fatty Acid Levels

Abstract
:1. Introduction
2. Materials and Methods
2.1. Lipid Compounds
2.2. Animals and Sample Collection
2.3. Fatty Acid Analysis
2.3.1. Sample Preparation for the Analysis of Fatty Acids
2.3.2. Gas Chromatography (GC)
2.4. Lipid Metabolite Analysis
2.4.1. Sample Preparation for the Analysis of Lipid Metabolites
2.4.2. LC/ESI-MS/MS
2.5. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Calder, P.C. Functional roles of fatty acids and their effects on human health. JPEN J. Parenter. Enter. Nutr. 2015, 39, 18S–32S. [Google Scholar] [CrossRef] [PubMed]
- Hudert, C.A.; Weylandt, K.H.; Lu, Y.; Wang, J.; Hong, S.; Dignass, A.; Serhan, C.N.; Kang, J.X. Transgenic mice rich in endogenous omega-3 fatty acids are protected from colitis. Proc. Natl. Acad. Sci. USA 2006, 103, 11276–11281. [Google Scholar] [CrossRef] [PubMed]
- Schmocker, C.; Weylandt, K.H.; Kahlke, L.; Wang, J.; Lobeck, H.; Tiegs, G.; Berg, T.; Kang, J.X. Omega-3 fatty acids alleviate chemically induced acute hepatitis by suppression of cytokines. Hepatology 2007, 45, 864–869. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Periz, A.; Planaguma, A.; Gronert, K.; Miquel, R.; Lopez-Parra, M.; Titos, E.; Horrillo, R.; Ferre, N.; Deulofeu, R.; Arroyo, V.; et al. Docosahexaenoic acid (DHA) blunts liver injury by conversion to protective lipid mediators: Protectin d1 and 17s-hydroxy-DHA. FASEB J. 2006, 20, 2537–2539. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, F.R.; Wei, D.; Jia, W.; Kang, J.X.; Stefanovic-Racic, M.; Dai, Y.; Zhao, A.Z. Endogenous omega-3 polyunsaturated fatty acid production confers resistance to obesity, dyslipidemia, and diabetes in mice. Mol. Endocrinol. 2014, 28, 1316–1328. [Google Scholar] [CrossRef] [PubMed]
- Jia, Q.; Lupton, J.R.; Smith, R.; Weeks, B.R.; Callaway, E.; Davidson, L.A.; Kim, W.; Fan, Y.Y.; Yang, P.; Newman, R.A.; et al. Reduced colitis-associated colon cancer in fat-1 (n-3 fatty acid desaturase) transgenic mice. Cancer Res. 2008, 68, 3985–3991. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.; Lee, A.; Shim, S.C.; Lee, J.H.; Choe, J.Y.; Ahn, H.; Choi, C.B.; Sung, Y.K.; Bae, S.C. Effect of n-3 polyunsaturated fatty acid supplementation in patients with rheumatoid arthritis: A 16-week randomized, double-blind, placebo-controlled, parallel-design multicenter study in Korea. J. Nutr. Biochem. 2013, 24, 1367–1372. [Google Scholar] [CrossRef] [PubMed]
- Mozaffarian, D.; Wu, J.H.; de Oliveira Otto, M.C.; Sandesara, C.M.; Metcalf, R.G.; Latini, R.; Libby, P.; Lombardi, F.; O’Gara, P.T.; Page, R.L.; et al. Fish oil and post-operative atrial fibrillation: A meta-analysis of randomized controlled trials. J. Am. Coll. Cardiol. 2013, 61, 2194–2196. [Google Scholar] [CrossRef] [PubMed]
- Feagan, B.G.; Sandborn, W.J.; Mittmann, U.; Bar-Meir, S.; D’Haens, G.; Bradette, M.; Cohen, A.; Dallaire, C.; Ponich, T.P.; McDonald, J.W.; et al. Omega-3 free fatty acids for the maintenance of remission in crohn disease: The epic randomized controlled trials. JAMA 2008, 299, 1690–1697. [Google Scholar] [CrossRef] [PubMed]
- Brasky, T.M.; Darke, A.K.; Song, X.; Tangen, C.M.; Goodman, P.J.; Thompson, I.M.; Meyskens, F.L., Jr.; Goodman, G.E.; Minasian, L.M.; Parnes, H.L.; et al. Plasma phospholipid fatty acids and prostate cancer risk in the select trial. J. Natl. Cancer Inst. 2013, 105, 1132–1141. [Google Scholar] [CrossRef] [PubMed]
- Mollace, V.; Gliozzi, M.; Carresi, C.; Musolino, V.; Oppedisano, F. Re-assessing the mechanism of action of n-3 pufas. Int. J. Cardiol. 2013, 170, S8–S11. [Google Scholar] [CrossRef] [PubMed]
- Bagga, D.; Wang, L.; Farias-Eisner, R.; Glaspy, J.A.; Reddy, S.T. Differential effects of prostaglandin derived from omega-6 and omega-3 polyunsaturated fatty acids on cox-2 expression and il-6 secretion. Proc. Natl. Acad. Sci. USA 2003, 100, 1751–1756. [Google Scholar] [CrossRef] [PubMed]
- Spite, M.; Claria, J.; Serhan, C.N. Resolvins, specialized proresolving lipid mediators, and their potential roles in metabolic diseases. Cell Metab. 2014, 19, 21–36. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Chiang, N. Resolution phase lipid mediators of inflammation: Agonists of resolution. Curr. Opin. Pharmacol. 2013, 13, 632–640. [Google Scholar] [CrossRef] [PubMed]
- Ostermann, A.I.; Willenberg, I.; Schebb, N.H. Comparison of sample preparation methods for the quantitative analysis of eicosanoids and other oxylipins in plasma by means of LC-MS/MS. Anal. Bioanal. Chem. 2015, 407, 1403–1414. [Google Scholar] [CrossRef] [PubMed]
- Gomolka, B.; Siegert, E.; Blossey, K.; Schunck, W.H.; Rothe, M.; Weylandt, K.H. Analysis of omega-3 and omega-6 fatty acid-derived lipid metabolite formation in human and mouse blood samples. Prostaglandins Other Lipid Mediat. 2011, 94, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Masoodi, M.; Mir, A.A.; Petasis, N.A.; Serhan, C.N.; Nicolaou, A. Simultaneous lipidomic analysis of three families of bioactive lipid mediators leukotrienes, resolvins, protectins and related hydroxy-fatty acids by liquid chromatography/electrospray ionisation tandem mass spectrometry. Rapid. Commun. Mass Spectrom 2008, 22, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Dolegowska, B.; Blogowski, W.; Safranow, K.; Domanski, L.; Jakubowska, K.; Olszewska, M. Lipoxygenase-derived hydroxyeicosatetraenoic acids—Novel perioperative markers of early post-transplant allograft function? Nephrol. Dial. Transpl. 2010, 25, 4061–4067. [Google Scholar] [CrossRef] [PubMed]
- Masoodi, M.; Pearl, D.S.; Eiden, M.; Shute, J.K.; Brown, J.F.; Calder, P.C.; Trebble, T.M. Altered colonic mucosal polyunsaturated fatty acid (PUFA) derived lipid mediators in ulcerative colitis: New insight into relationship with disease activity and pathophysiology. PLoS ONE 2013, 8, e76532. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.X.; Wang, J.; Wu, L.; Kang, Z.B. Transgenic mice: Fat-1 mice convert n-6 to n-3 fatty acids. Nature 2004. [Google Scholar] [CrossRef] [PubMed]
- Weylandt, K.H.; Chiu, C.Y.; Gomolka, B.; Waechter, S.F.; Wiedenmann, B. Omega-3 fatty acids and their lipid mediators: Towards an understanding of resolvin and protectin formation. Prostaglandins Other Lipid Mediat. 2012, 97, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Weylandt, K.H.; Krause, L.F.; Gomolka, B.; Chiu, C.Y.; Bilal, S.; Nadolny, A.; Waechter, S.F.; Fischer, A.; Rothe, M.; Kang, J.X. Suppressed liver tumorigenesis in fat-1 mice with elevated omega-3 fatty acids is associated with increased omega-3 derived lipid mediators and reduced tnf-alpha. Carcinogenesis 2011, 32, 897–903. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.X.; Wang, J. A simplified method for analysis of polyunsaturated fatty acids. BMC Biochem. 2005, 6, 5. [Google Scholar] [CrossRef] [PubMed]
- Arnold, C.; Markovic, M.; Blossey, K.; Wallukat, G.; Fischer, R.; Dechend, R.; Konkel, A.; von Schacky, C.; Luft, F.C.; Muller, D.N.; et al. Arachidonic acid-metabolizing cytochrome P450 enzymes are targets of ω-3 fatty acids. J. Biol. Chem. 2010, 285, 32720–32733. [Google Scholar] [CrossRef] [PubMed]
- Fischer, R.; Konkel, A.; Mehling, H.; Blossey, K.; Gapelyuk, A.; Wessel, N.; von Schacky, C.; Dechend, R.; Muller, D.N.; Rothe, M.; et al. Dietary omega-3 fatty acids modulate the eicosanoid profile in man primarily via the cyp-epoxygenase pathway. J. Lipid Res. 2014, 55, 1150–1164. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, M.T.; Nara, T.Y. Essential fatty acid synthesis and its regulation in mammals. Prostaglandins Leukot. Essent. Fatty Acids 2003, 68, 145–150. [Google Scholar] [CrossRef]
- Ji, S.; Hardy, R.W.; Wood, P.A. Transgenic expression of n-3 fatty acid desaturase (fat-1) in C57/BL6 mice: Effects on glucose homeostasis and body weight. J. Cell. Biochem. 2009, 107, 809–817. [Google Scholar] [CrossRef] [PubMed]
- Xia, S.; Lu, Y.; Wang, J.; He, C.; Hong, S.; Serhan, C.N.; Kang, J.X. Melanoma growth is reduced in fat-1 transgenic mice: Impact of omega-6/omega-3 essential fatty acids. Proc. Natl. Acad. Sci. USA 2006, 103, 12499–12504. [Google Scholar] [CrossRef] [PubMed]
- Bilal, S.; Haworth, O.; Wu, L.; Weylandt, K.H.; Levy, B.D.; Kang, J.X. Fat-1 transgenic mice with elevated omega-3 fatty acids are protected from allergic airway responses. Biochim. Biophys. Acta 2011, 1812, 1164–1169. [Google Scholar] [CrossRef] [PubMed]
- Guillou, H.; Zadravec, D.; Martin, P.G.; Jacobsson, A. The key roles of elongases and desaturases in mammalian fatty acid metabolism: Insights from transgenic mice. Prog. Lipid Res. 2010, 49, 186–199. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Hong, S.; Gronert, K.; Colgan, S.P.; Devchand, P.R.; Mirick, G.; Moussignac, R.L. Resolvins: A family of bioactive products of omega-3 fatty acid transformation circuits initiated by aspirin treatment that counter proinflammation signals. J. Exp. Med. 2002, 196, 1025–1037. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Hamberg, M.; Samuelsson, B. Lipoxins: Novel series of biologically active compounds formed from arachidonic acid in human leukocytes. Proc. Natl. Acad. Sci. USA 1984, 81, 5335–5339. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Clish, C.B.; Brannon, J.; Colgan, S.P.; Chiang, N.; Gronert, K. Novel functional sets of lipid-derived mediators with antiinflammatory actions generated from omega-3 fatty acids via cyclooxygenase 2-nonsteroidal antiinflammatory drugs and transcellular processing. J. Exp. Med. 2000, 192, 1197–1204. [Google Scholar] [CrossRef] [PubMed]
- Astarita, G.; McKenzie, J.H.; Wang, B.; Strassburg, K.; Doneanu, A.; Johnson, J.; Baker, A.; Hankemeier, T.; Murphy, J.; Vreeken, R.J.; et al. A protective lipidomic biosignature associated with a balanced omega-6/omega-3 ratio in fat-1 transgenic mice. PLoS ONE 2014, 9, e96221. [Google Scholar] [CrossRef] [PubMed]
- Endo, J.; Sano, M.; Isobe, Y.; Fukuda, K.; Kang, J.X.; Arai, H.; Arita, M. 18-hepe, an n-3 fatty acid metabolite released by macrophages, prevents pressure overload-induced maladaptive cardiac remodeling. J. Exp. Med. 2014, 211, 1673–1687. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.Y.; Gomolka, B.; Dierkes, C.; Huang, N.R.; Schroeder, M.; Purschke, M.; Manstein, D.; Dangi, B.; Weylandt, K.H. Omega-6 docosapentaenoic acid-derived resolvins and 17-hydroxydocosahexaenoic acid modulate macrophage function and alleviate experimental colitis. Inflamm. Res. 2012, 61, 967–976. [Google Scholar] [CrossRef] [PubMed]
- Bento, A.F.; Claudino, R.F.; Dutra, R.C.; Marcon, R.; Calixto, J.B. Omega-3 fatty acid-derived mediators 17(r)-hydroxy docosahexaenoic acid, aspirin-triggered resolvin d1 and resolvin d2 prevent experimental colitis in mice. J. Immunol. 2011, 187, 1957–1969. [Google Scholar] [CrossRef] [PubMed]
- Kohnke, T.; Gomolka, B.; Bilal, S.; Zhou, X.; Sun, Y.; Rothe, M.; Baumgart, D.C.; Weylandt, K.H. Acetylsalicylic acid reduces the severity of dextran sodium sulfate-induced colitis and increases the formation of anti-inflammatory lipid mediators. Biomed. Res. Int. 2013. [Google Scholar] [CrossRef] [PubMed]


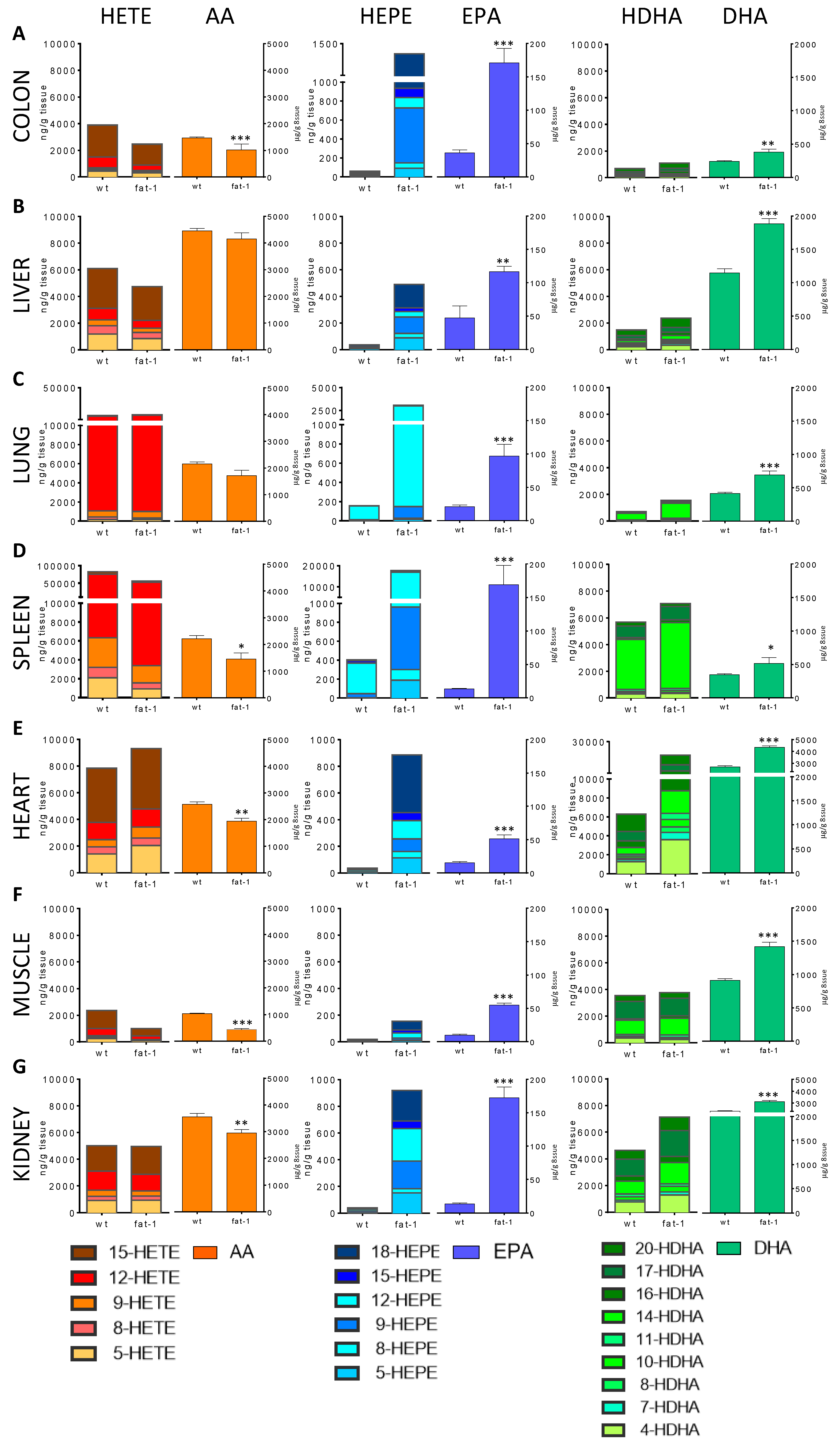{kind=link}
| Organ | Fatty Acid | Wildtype (μg/g) | Fat-1 (μg/g) | p-Value | AA Difference (-Fold) | EPA Difference (-Fold) | DHA Difference (-Fold) |
|---|---|---|---|---|---|---|---|
| Colon | AA | 1470 ± 41.0 | 1023 ± 108 | 0.001 | 0.70 | ||
| EPA | 36.0 ± 4.7 | 171.2 ± 21.7 | <0.001 | 4.8 | |||
| DHA | 239.7 ± 11.2 | 380.7 ± 42.1 | 0.003 | 1.6 | |||
| Liver | AA | 4448 ± 94.9 | 4152 ± 225 | 0.2 | 0.93 | ||
| EPA | 47.3 ± 17.9 | 116.5 ± 8.1 | 0.02 | 2.5 | |||
| DHA | 1147 ± 62.9 | 1883 ± 80.1 | <0.001 | 1.6 | |||
| Lung | AA | 2154 ± 71.2 | 1714 ± 199 | 0.06 | 0.80 | ||
| EPA | 20.8 ± 3.0 | 96.7 ± 17.6 | 0.001 | 4.6 | |||
| DHA | 415.7 ± 17.9 | 692.7 ± 59.2 | 0.001 | 1.7 | |||
| Spleen | AA | 2195 ± 173.7 | 1444 ± 234.5 | 0.03 | 0.66 | ||
| EPA | 13.4 ± 0.8 | 169.2 ± 29.0 | <0.001 | 12.6 | |||
| DHA | 359.8 ± 22.7 | 538.9 ± 88.6 | 0.03 | 1.5 | |||
| Heart | AA | 2574 ± 93 | 1942 ± 105 | 0.002 | 0.75 | ||
| EPA | 15.3 ± 1.9 | 51.37 ± 5.8 | <0.001 | 3.4 | |||
| DHA | 2689 ± 109 | 4340 ± 126 | <0.001 | 1.6 | |||
| Muscle | AA | 1039 ± 14.3 | 442.5 ± 32.5 | <0.001 | 0.43 | ||
| EPA | 9.9 ± 1.4 | 54.7 ± 3.1 | <0.001 | 5.5 | |||
| DHA | 913.9 ± 25.2 | 1420 ± 66.7 | <0.001 | 1.6 | |||
| Kidney | AA | 3556 ± 134 | 2947 ± 132 | 0.02 | 0.83 | ||
| EPA | 13.8 ± 1.4 | 172.3 ± 16.3 | <0.001 | 12.4 | |||
| DHA | 2304 ± 54.8 | 3110 ± 114 | <0.001 | 1.4 |
| Organ | Lipid Metabolites | Wildtype (ng/g) | Fat-1 (ng/g) | p-Value | HETE Difference (-Fold) | HEPE difference (-Fold) | HDHA Difference (-Fold) |
|---|---|---|---|---|---|---|---|
| Colon | HETE | 4260 ± 282 | 2707 ± 249 | 0.005 | 0.64 | ||
| HEPE | 61.0 ± 7.8 | 1340 ± 94.2 | <0.001 | 21.9 | |||
| HDHA | 746.1 ± 42.6 | 1183 ± 103 | 0.001 | 1.6 | |||
| Liver | HETE | 6336 ± 698 | 4940 ± 298 | 0.18 | 0.78 | ||
| HEPE | 36.0 ± 4.4 | 489.0 ± 90.4 | <0.001 | 13.6 | |||
| HDHA | 1586 ± 186 | 2543 ± 168 | 0.008 | 1.6 | |||
| Lung | HETE | 16,362 ± 1641 | 17,290 ± 1926 | 0.73 | 1.06 | ||
| HEPE | 156.1 ± 24.5 | 3057 ± 530 | <0.001 | 19.6 | |||
| HDHA | 740.6 ± 102 | 1629 ± 152 | <0.001 | 2.2 | |||
| Spleen | HETE | 86,511 ± 3888 | 58,515 ± 2095 | <0.001 | 0.68 | ||
| HEPE | 403.2 ± 63.2 | 17,711 ± 2181 | <0.001 | 43.9 | |||
| HDHA | 5995 ± 791 | 7425 ± 485 | 0.24 | 1.2 | |||
| Heart | HETE | 8094 ± 845 | 9826 ± 3023 | 0.50 | 1.21 | ||
| HEPE | 36.6 ± 10 | 885.9 ± 163 | <0.001 | 24.2 | |||
| HDHA | 6642 ± 960 | 22,855 ± 8748 | 0.03 | 3.4 | |||
| Muscle | HETE | 2471 ± 234 | 1057 ± 129 | 0.002 | 0.43 | ||
| HEPE | 19.4 ± 2.4 | 156.9 ± 20.9 | <0.001 | 8.1 | |||
| HDHA | 3759 ± 347 | 4021 ± 459 | 0.66 | 1.1 | |||
| Kidney | HETE | 5487 ± 719 | 5339 ± 1541 | 0.92 | 0.97 | ||
| HEPE | 40.3 ± 6.6 | 919.2 ± 417 | 0.02 | 22.8 | |||
| HDHA | 5108 ± 832 | 7896 ± 2509 | 0.22 | 1.6 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiu, C.-Y.; Smyl, C.; Dogan, I.; Rothe, M.; Weylandt, K.-H. Quantitative Profiling of Hydroxy Lipid Metabolites in Mouse Organs Reveals Distinct Lipidomic Profiles and Modifications Due to Elevated n-3 Fatty Acid Levels. Biology 2017, 6, 9. https://doi.org/10.3390/biology6010009
Chiu C-Y, Smyl C, Dogan I, Rothe M, Weylandt K-H. Quantitative Profiling of Hydroxy Lipid Metabolites in Mouse Organs Reveals Distinct Lipidomic Profiles and Modifications Due to Elevated n-3 Fatty Acid Levels. Biology. 2017; 6(1):9. https://doi.org/10.3390/biology6010009
Chicago/Turabian StyleChiu, Cheng-Ying, Christopher Smyl, Inci Dogan, Michael Rothe, and Karsten-H. Weylandt. 2017. "Quantitative Profiling of Hydroxy Lipid Metabolites in Mouse Organs Reveals Distinct Lipidomic Profiles and Modifications Due to Elevated n-3 Fatty Acid Levels" Biology 6, no. 1: 9. https://doi.org/10.3390/biology6010009
APA StyleChiu, C.-Y., Smyl, C., Dogan, I., Rothe, M., & Weylandt, K.-H. (2017). Quantitative Profiling of Hydroxy Lipid Metabolites in Mouse Organs Reveals Distinct Lipidomic Profiles and Modifications Due to Elevated n-3 Fatty Acid Levels. Biology, 6(1), 9. https://doi.org/10.3390/biology6010009