The Role of the Bacterial Flagellum in Adhesion and Virulence

Abstract
:1. Introduction

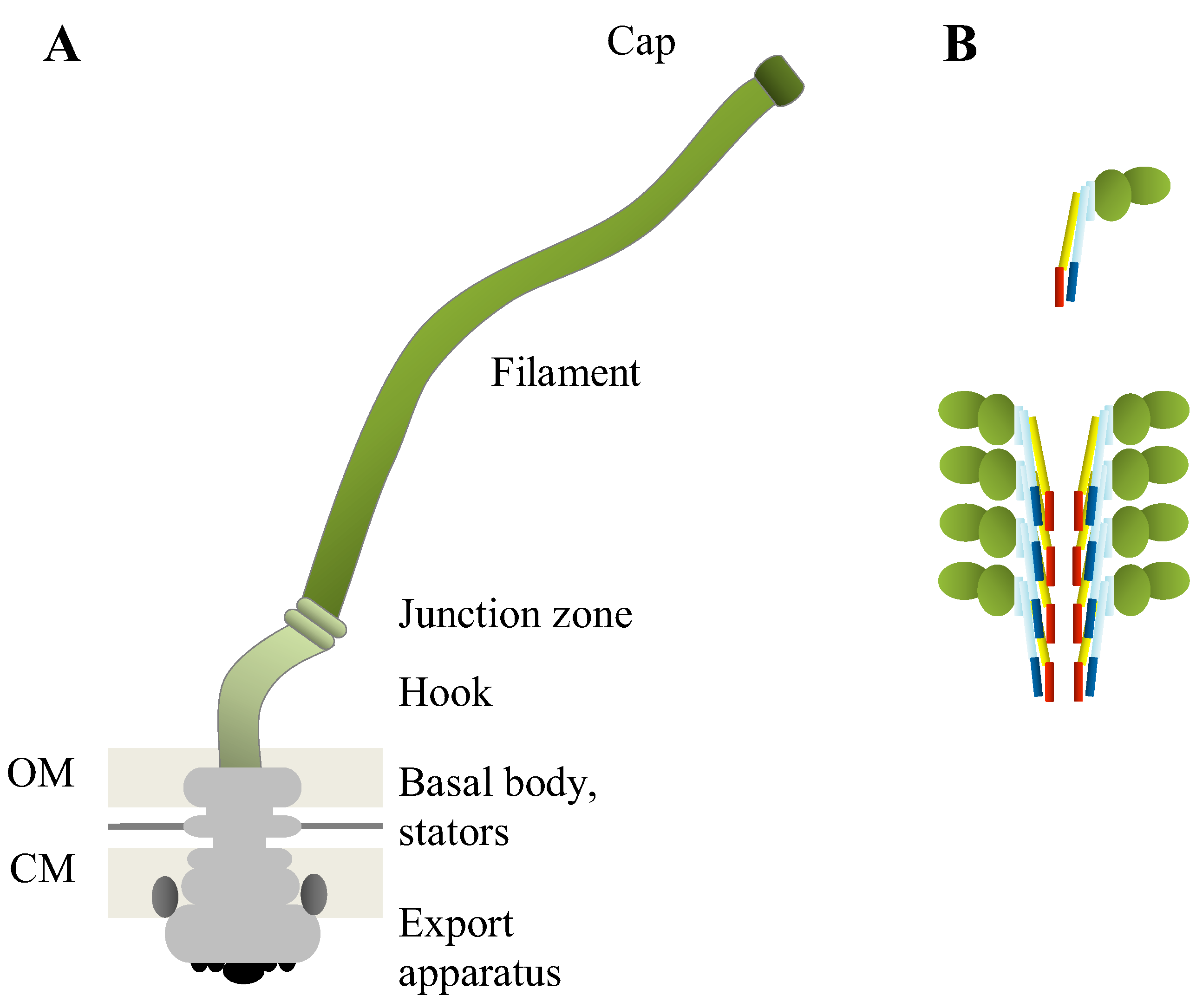
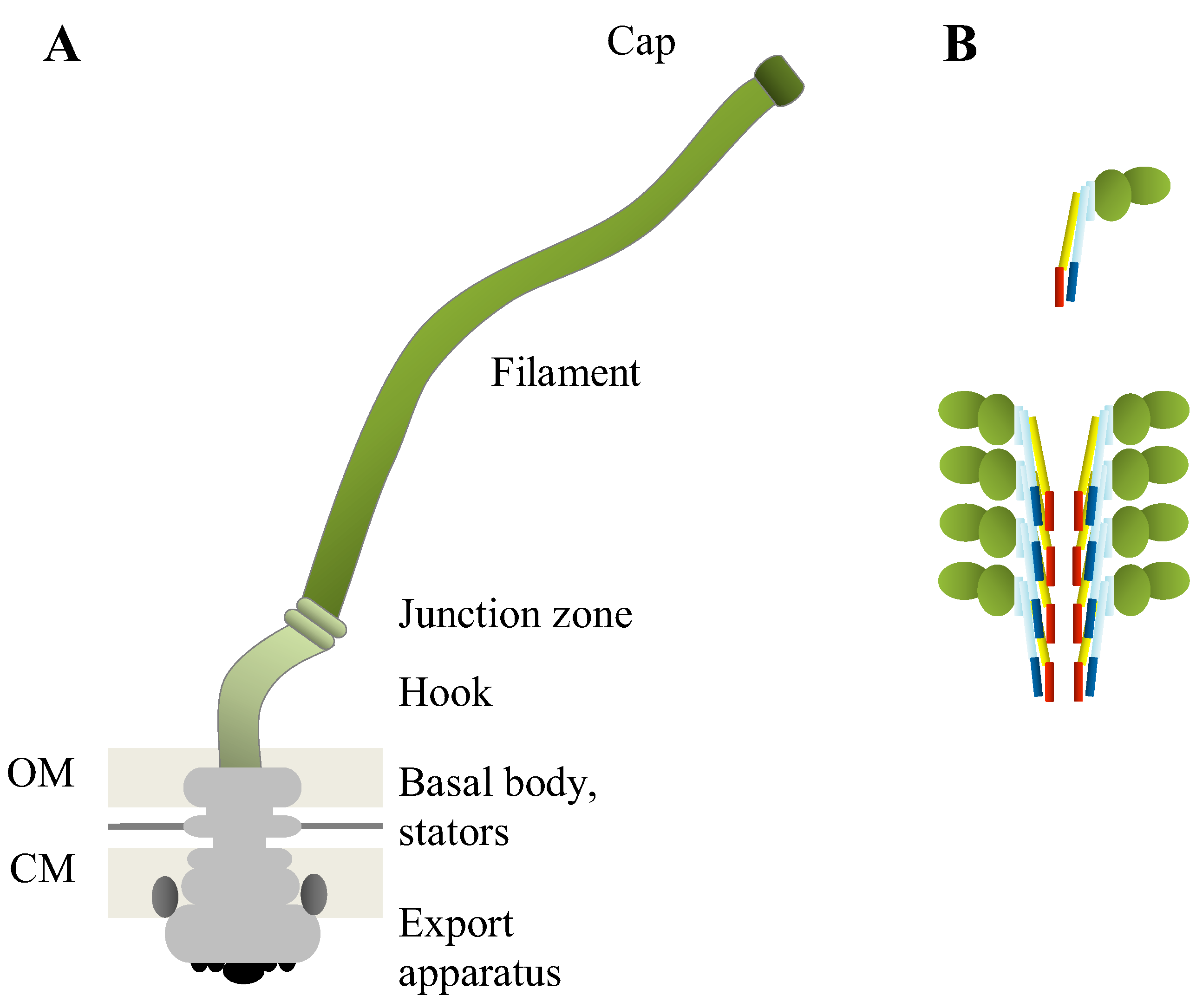{kind=link}
{kind=link}
| Role in/Part of flagellum | Protein name/Number of proteins | Function |
|---|---|---|
| Regulation of flagellar biosynthesis | FlhC, FlhD, FlbA | Regulators |
| FliI | Export-related ATPase | |
| FliA | Flagellar sigma factor | |
| Motility/chemotaxis | CheA | Smooth swimming |
| CheB | Tumbling | |
| CheW, CheV | Chemotaxis | |
| MotA, MotB | Rotation of flagellum | |
| Export machinery | 9 different proteins | Protein export |
| FlhE | Chaperone or plug | |
| Basal body | 20 different proteins | Cell-wall anchor, rotor, holds export apparatus |
| FlgJ | Muramidase | |
| Hook | FlgE | Hook |
| Hook-filament junction | FlgK, FlgL | Connect filament to hook |
| Filament | FliC (alternative names FljB, FlaA, FlaB) | Main structural subunit |
| FliD | Filament cap |
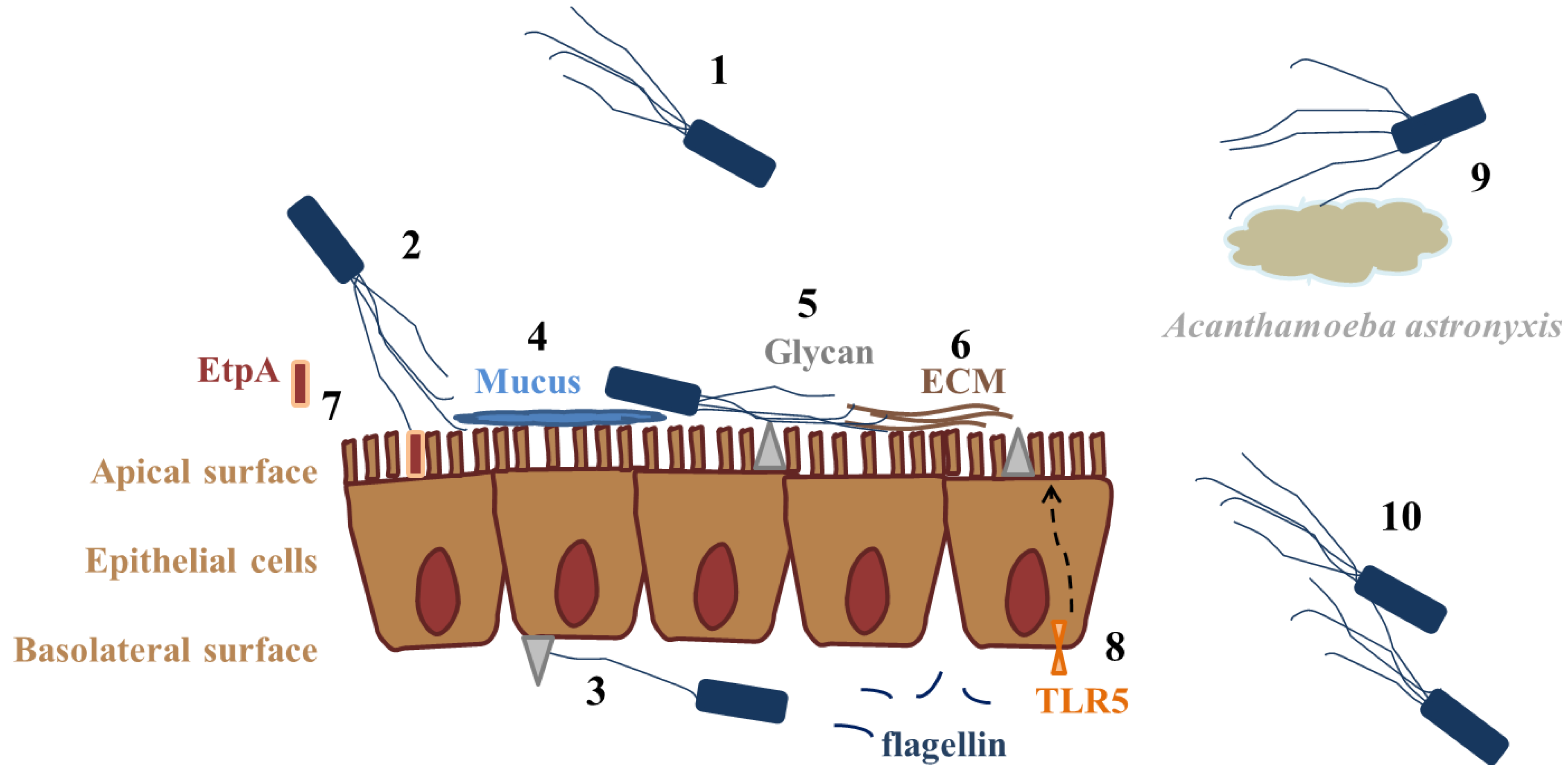
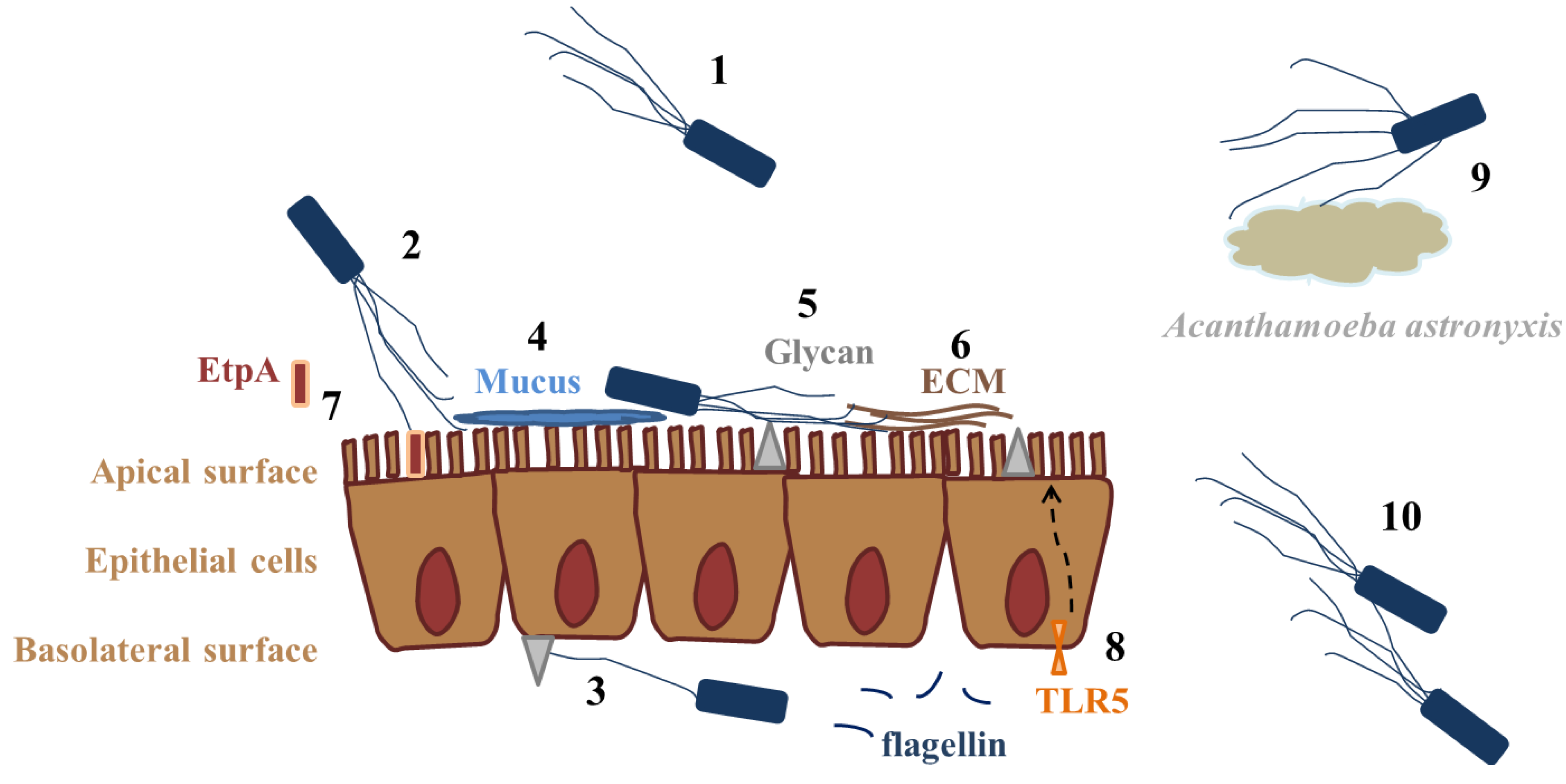
| Role of flagella/Bacterial species | Flagellum/Protein/Gene | Effect/Role in virulence | Target/Administration route | Receptor | Reference |
|---|---|---|---|---|---|
| Adhesion | |||||
| Bordetella bronchiseptica, Bordetella pertussis | Flagellum | Adhesion | HeLa cells | ND | [18] |
| Burkholderia pseudomallei | Flagellum | Adhesion; invasion | Acanthamoeba astronyxis | ND | [19] |
| Campylobacter jejuni | Flagellum | Adhesion | Intestine-407 | ND | [20] |
| Clostridium difficile | FliC, FliD; flagellum | Binding; no effect | Mouse cecal mucus, hamster | ND | [21,22] |
| Escherichia coli | Flagellum, FliC | Adhesion; Microcolony formation | HeLa cells | ND | [23] |
| Flagellum, FliC | Binding | Mucins (a), bovine intestinal mucus, laminin, collagen | ND | [24] | |
| Flagellum | Adhesion; invasion | Polarized Caco-2BBe, T-84; BREC; Caco-2; T-84; IPEC-J2; IPEC-1 | ND | [25,26,27,28,29,30] | |
| Flagellum | Association | HBMEC | ND | [31] | |
| Flagellum, FliC | Adhesion | Caco-2, mice intestine | EtpA | [32] | |
| FliC | Adhesion | Human intestinal cryosections, LS174T | gluconate | [33] | |
| Pseudomonas aeruginosa | FliC | Adhesion; virulence | 1HAEo; mice | GM1, GD1a, asialo-GM1 | [34] |
| FliC; FliD | Adhesion | mucin, MUC1 | ND | [35,36,37] | |
| FliD | Adhesion | human respiratory mucin | Lewis x glycotype | [38] | |
| Flagellum | Adhesion | Calu-3 basolateral surface | HSPGs (b) | [39] | |
| Salmonella enterica serovar Dublin | Flagellum | Adhesion; virulence | Intestine-407; orally in mice | ND | [40] |
| Salmonella enterica serovar Typhimurium | Flagellum | Adhesion | Intestine-407 | ND | [40] |
| Stenotrophomonas maltophilia | Flagellin | Adhesion | Mouse tracheal mucus | ND | [41] |
| Motility | |||||
| Cronobacter sakazakii | Flagellum | Adhesion; biofilm formation | Caco-2 | ND | [42] |
| Escherichia coli | Flagellum | Mucus penetration; persistence; invasion | Hep-2, HT2916E, HT2919A; orally in chicks; HCT-8 | asialo-GM1, lipid rafts | [43,44,45,46,47,48] |
| FliC | Virulence | Orally in Sm-treated mice | ND | [49] | |
| Helicobacter pylori | Flagellum | Colonization | Gastric epithelium, orally in mice | ND | [49] |
| Proteus mirabilis | Flagellum | Invasion; colonization | Human renal proximal tubular epithelial cells, EJ/28 | ND | [50,51] |
| Salmonella enterica serovar Dublin | Flagellum | Increased invasion | Intestine-407 | ND | [40] |
| Salmonella enterica serovar Enteritidis | Flagellum | Increased invasion | Caco-2, Hep-2, Div-1 | ND | [52,53,54] |
| Salmonella enterica serovar Typhimurium | Flagellum | Increased adhesion; invasion | Intestine-407 | ND | [40] |
| Regulation | |||||
| Escherichia coli | flhDC | Adhesion, invasion | Intestine-407 | ND | [55] |
| flhC | Colonization | Orally in cattle | ND | [56] | |
| FliA | Adhesion, invasion | Intestine-407 | ND | [57] | |
| Helicobacter pylori | flbA | Adhesion | Kato III, primary human gastric epithelial cells | ND | [58] |
2. Motility and Virulence
2.1. Flagellum Affects Virulence Mainly by Facilitating Motility
| Abbreviation | Cell line | Origin |
|---|---|---|
| 1HAEo | airway epithelial cells | human |
| A549 | lung epithelial cells | human |
| BREC | primary rectal epithelial cells | bovine |
| Caco-2 | colorectal adenocarcinoma epithelial cells | human |
| Caco-2BBe | colorectal adenocarcinoma epithelial cells expressing enterocyte-like brush border | human |
| Calu-3 | lung adenocarcinoma epithelial cells | human |
| CHO Lec-2 | relatively ganglioside deficient CHO derivative | Chinese hamster |
| CHO-Muc1 | mucin 1-expressing ovary cells | Chinese hamster |
| Div-1 | gastrointestinal mucosal cells | avian |
| EJ/28 | urothelial cells | human |
| 16HBE | polarized airway epithelial cells | human |
| HBMEC | brain microvascular endothelial cells | human |
| HCT-8 | ileocecal colorectal adenocarcinoma cells | human |
| HeLa | cervical tumor cells | human |
| Hep-2 | epidermoid cancer cell line contaminated by HeLa cells | human |
| HT29 | colorectal adenocarcinoma epithelial cells | human |
| HT2916E | mucus-secreting subclone of HT29 | human |
| HT2919A | non-mucus-secreting subclone of HT29 | human |
| Int(estine)-407 | intestinal embryonic jejunum and ileum epithelial cells contaminated by HeLa cells | human |
| IPEC-1 | neonatal ileal and jejunal epithelial cells | piglet |
| IPEC-J2 | neonatal jejunal columnar epithelial cells | piglet |
| Kato III | gastric adenocarcinoma cells | human |
| LS174T | mucin-secreting colorectal adenocarcinoma epithelial cells | human |
| RAW264.7 | macrophage cells | murine |
| T24 | bladder epithelial cells | human |
| T-84 | colorectal adenocarcinoma cells | human |
2.2. Flagellum Affects Virulence by Regulating Other Virulence Factors
3. Flagellum-mediated Adhesion
3.1. Escherichia coli
3.1.1. E. coli Pathovars and Role of Flagella in Virulence
| Pathovar | Abbreviation | Disease/symptoms | Major site of colonization | Reference |
|---|---|---|---|---|
| IPEC | ||||
| Enterohaemorrhagic E. coli | EHEC | Gastroenteritis; hemorrhagic colitis; HUS (a) | Large bowel | [65,69] |
| Enteropathogenic E. coli | EPEC | Infant diarrhea in developing countries | Small bowel | [65,69] |
| Atypical enteropathogenic E. coli | aEPEC | Persistent diarrhea in children | Small bowel | [69] |
| Enterotoxigenic E. coli | ETEC | Infant diarrhea; travelers’ diarrhea | Small bowel | [65,69] |
| Enteroinvasive E. coli | EIEC | Dysentery | Large bowel | [65,69] |
| Enteroaggregative E. coli | EAEC | Variable (from non-symptomatic to HUS) | Large bowel, small bowel | [65,69] |
| Diffusely adherent E. coli | DAEC | Diarrhea in young children; nosocomial adult diarrhea | Small bowel | [65,69] |
| Shiga-toxin producing E. coli | STEC | Dependent on Shiga-toxin expressing pathovar | Dependent on pathovar | [69] |
| ExPEC | ||||
| Newborn-meningitis causing E. coli | NMEC | Newborn meningitis; septicaemia | Brain, blood | [65,68,71] |
| Uropathogenic E. coli | UPEC | Urinary tract infections; septicaemia | Bladder, kidney, blood | [65,68] |
| Other | ||||
| Adherent-invasive E. coli | AIEC | Crohn’s disease | Ileum of small bowel | [65,67,68,69] |
| Avian pathogenic E. coli | APEC | Avian colibacillosis | Systemic infection | [67,68] |
| Porcine pathogenic E. coli | Porcine STEC | Post-weaning diarrhea; edema disease | Intestine;vasculature | [28,29,48] |
3.1.2. Flagellum-mediated IPEC Adhesion
3.1.3. Flagellum-mediated ExPEC Adhesion
3.1.4. Role of Flagellum in AIEC Adhesion
3.1.5. Flagellum-mediated Adhesion of Porcine Pathogenic E. coli
3.1.6. Flagellum-mediated Adhesion of Probiotic E. coli
3.1.7. Flagellum-mediated Indirect E. coli Adhesion
3.2. Pseudomonas aeruginosa
3.3. Clostridium difficile
3.4. Flagellar Adhesion of Additional Bacterial Species
4. Additional Adhesion-related Mechanisms of Flagella
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Duan, Q.; Zhou, M.; Zhu, L.; Zhu, G. Flagella and bacterial pathogenicity. J. Basic Microb. 2013, 53, 1–8. [Google Scholar] [CrossRef]
- Prüss, B.M.; Besemann, C.; Denton, A.; Wolfe, A.J. A complex transcription network controls the early stages of biofilm development by Escherichia coli. J. Bacteriol. 2006, 188, 3731–3739. [Google Scholar] [CrossRef]
- Conrad, J.C. Physics of bacterial near-surface motility using flagella and type IV pili: Implications for biofilm formation. Res. Microbiol. 2012, 163, 619–629. [Google Scholar] [CrossRef]
- Chevance, F.F.; Hughes, K.T. Coordinating assembly of a bacterial macromolecular machine. Nat. Rev. Microbiol. 2008, 6, 455–465. [Google Scholar] [CrossRef]
- Sourjik, V.; Wingreen, N.S. Responding to chemical gradients: Bacterial chemotaxis. Curr. Opin. Cell Biol. 2012, 24, 262–268. [Google Scholar] [CrossRef]
- Erhardt, M.; Namba, K.; Hughes, K.T. Bacterial nanomachines: The flagellum and type III injectisome. Cold Spring Harbor Perspect. Biol. 2010, 2, a000299. [Google Scholar] [CrossRef]
- Büttner, D. Protein export according to schedule: Architecture, assembly, and regulation of type III secretion systems from plant- and animal-pathogenic bacteria. Microbiol. Mol. Biol. Rev. 2012, 76, 262–310. [Google Scholar] [CrossRef]
- Smith, T.G.; Hoover, T.R. Deciphering bacterial flagellar gene regulatory networks in the genomic era. Adv. Appl. Microbiol. 2009, 67, 257–295. [Google Scholar] [CrossRef]
- Klemm, P.; Vejborg, R.M.; Hancock, V. Prevention of bacterial adhesion. Appl. Microbiol. Biotechnol. 2010, 88, 451–459. [Google Scholar] [CrossRef]
- Young, G.M.; Schmiel, D.H.; Miller, V.L. A new pathway for the secretion of virulence factors by bacteria: The flagellar export apparatus functions as a protein-secretion system. Proc. Natl. Acad. Sci. USA 1999, 96, 6456–6461. [Google Scholar] [CrossRef]
- Majander, K.; Anton, L.; Antikainen, J.; Lång, H.; Brummer, M.; Korhonen, T.K.; Westerlund-Wikström, B. Extracellular secretion of polypeptides using a modified Escherichia coli flagellar secretion apparatus. Nat. Biotechnol. 2005, 23, 475–481. [Google Scholar] [CrossRef]
- Hayashi, F.; Smith, K.D.; Ozinsky, A.; Hawn, T.R.; Yi, E.C.; Goodlett, D.R.; Eng, J.K.; Akira, S.; Underhill, D.M.; Aderem, A. The innate immune response to bacterial flagellin is mediated by Toll-like receptor 5. Nature 2001, 410, 1099–1103. [Google Scholar] [CrossRef]
- Gong, Y.N.; Shao, F. Sensing bacterial infections by NAIP receptors in NLRC4 inflammasome activation. Protein Cell 2012, 3, 98–105. [Google Scholar] [CrossRef]
- Halff, E.F.; Diebolder, C.A.; Versteeg, M.; Schouten, A.; Brondijk, T.H.; Huizinga, E.G. Formation and structure of a NAIP5-NLRC4 inflammasome induced by direct interactions with conserved N- and C-terminal regions of flagellin. J. Biol. Chem. 2012, 287, 38460–38472. [Google Scholar]
- Yoon, S.I.; Kurnasov, O.; Natarajan, V.; Hong, M.; Gudkov, A.V.; Osterman, A.L.; Wilson, I.A. Structural basis of TLR5-flagellin recognition and signaling. Science 2012, 335, 859–864. [Google Scholar]
- Yonekura, K.; Maki-Yonekura, S.; Namba, K. Complete atomic model of the bacterial flagellar filament by electron cryomicroscopy. Nature 2003, 424, 643–650. [Google Scholar] [CrossRef]
- Beatson, S.A.; Minamino, T.; Pallen, M.J. Variation in bacterial flagellins: From sequence to structure. Trends Microbiol. 2006, 14, 151–155. [Google Scholar] [CrossRef]
- Savelkoul, P.H.; de Kerf, D.P.; Willems, R.J.; Mooi, F.R.; van der Zeijst, B.A.; Gaastra, W. Characterization of the fim2 and fim3 fimbrial subunit genes of Bordetella bronchiseptica: Roles of Fim2 and Fim3 fimbriae and flagella in adhesion. Infect. Immun. 1996, 64, 5098–5105. [Google Scholar]
- Inglis, T.J.; Robertson, T.; Woods, D.E.; Dutton, N.; Chang, B.J. Flagellum-mediated adhesion by Burkholderia pseudomallei precedes invasion of Acanthamoeba astronyxis. Infect. Immun. 2003, 71, 2280–2282. [Google Scholar] [CrossRef]
- McSweegan, E.; Walker, R.I. Identification and characterization of two Campylobacter jejuni adhesins for cellular and mucous substrates. Infect. Immun. 1986, 53, 141–148. [Google Scholar]
- Tasteyre, A.; Barc, M.C.; Collignon, A.; Boureau, H.; Karjalainen, T. Role of FliC and FliD flagellar proteins of Clostridium difficile in adherence and gut colonization. Infect. Immun. 2001, 69, 7937–7940. [Google Scholar] [CrossRef]
- Dingle, T.C.; Mulvey, G.L.; Armstrong, G.D. Mutagenic analysis of the Clostridium difficile flagellar proteins, FliC and FliD, and their contribution to virulence in hamsters. Infect. Immun. 2011, 79, 4061–4067. [Google Scholar] [CrossRef]
- Girón, J.A.; Torres, A.G.; Freer, E.; Kaper, J.B. The flagella of enteropathogenic Escherichia coli mediate adherence to epithelial cells. Mol. Microbiol. 2002, 44, 361–379. [Google Scholar] [CrossRef]
- Erdem, A.L.; Avelino, F.; Xicohtencatl-Cortes, J.; Girón, J.A. Host protein binding and adhesive properties of H6 and H7 flagella of attaching and effacing Escherichia coli. J. Bacteriol. 2007, 189, 7426–7435. [Google Scholar] [CrossRef]
- Eaves-Pyles, T.; Allen, C.A.; Taormina, J.; Swidsinski, A.; Tutt, C.B.; Jezek, G.E.; Islas-Islas, M.; Torres, A.G. Escherichia coli isolated from a Crohn’s disease patient adheres, invades, and induces inflammatory responses in polarized intestinal epithelial cells. Int. J. Med. Microbiol. 2008, 298, 397–409. [Google Scholar] [CrossRef]
- Mahajan, A.; Currie, C.G.; Mackie, S.; Tree, J.; McAteer, S.; McKendrick, I.; McNeilly, T.N.; Roe, A.; La Ragione, R.M.; Woodward, M.J.; et al. An investigation of the expression and adhesin function of H7 flagella in the interaction of Escherichia coli O157:H7 with bovine intestinal epithelium. Cell. Microbiol. 2009, 11, 121–137. [Google Scholar] [CrossRef]
- Sampaio, S.C.; Gomes, T.A.; Pichon, C.; du Merle, L.; Guadagnini, S.; Abe, C.M.; Sampaio, J.L.; Le Bouguénec, C. The flagella of an atypical enteropathogenic Escherichia coli strain are required for efficient interaction with and stimulation of interleukin-8 production by enterocytes in vitro. Infect. Immun. 2009, 77, 4406–4413. [Google Scholar] [CrossRef]
- Duan, Q.; Zhou, M.; Zhu, X.; Bao, W.; Wu, S.; Ruan, X.; Zhang, W.; Yang, Y.; Zhu, J.; Zhu, G. The flagella of F18ab Escherichia coli is a virulence factor that contributes to infection in a IPEC-J2 cell model in vitro. Vet. Microbiol. 2012, 160, 132–140. [Google Scholar] [CrossRef]
- Duan, Q.; Zhou, M.; Zhu, X.; Yang, Y.; Zhu, J.; Bao, W.; Wu, S.; Ruan, X.; Zhang, W.; Zhu, G. Flagella from F18+ Escherichia coli play a role in adhesion to pig epithelial cell lines. Microb. Pathog. 2013, 55, 32–38. [Google Scholar] [CrossRef]
- Zhou, M.; Duan, Q.; Zhu, X.; Guo, Z.; Li, Y.; Hardwidge, P.R.; Zhu, G. Both flagella and F4 fimbriae from F4ac+ enterotoxigenic Escherichia coli contribute to attachment to IPEC-J2 cells in vitro. Vet. Res. 2013, 44, 30. [Google Scholar] [CrossRef]
- Parthasarathy, G.; Yao, Y.; Kim, K.S. Flagella promote Escherichia coli K1 association with and invasion of human brain microvascular endothelial cells. Infect. Immun. 2007, 75, 2937–2945. [Google Scholar] [CrossRef]
- Roy, K.; Hilliard, G.M.; Hamilton, D.J.; Luo, J.; Ostmann, M.M.; Fleckenstein, J.M. Enterotoxigenic Escherichia coli EtpA mediates adhesion between flagella and host cells. Nature 2009, 457, 594–598. [Google Scholar] [CrossRef]
- Troge, A.; Scheppach, W.; Schroeder, B.O.; Rund, S.A.; Heuner, K.; Wehkamp, J.; Stange, E.F.; Oelschlaeger, T.A. More than a marine propeller—The flagellum of the probiotic Escherichia coli strain Nissle 1917 is the major adhesin mediating binding to human mucus. Int. J. Med. Microbiol. 2012, 302, 304–314. [Google Scholar] [CrossRef]
- Feldman, M.; Bryan, R.; Rajan, S.; Scheffler, L.; Brunnert, S.; Tang, H.; Prince, A. Role of flagella in pathogenesis of Pseudomonas aeruginosa pulmonary infection. Infect. Immun. 1998, 66, 43–51. [Google Scholar]
- Arora, S.K.; Ritchings, B.W.; Almira, E.C.; Lory, S.; Ramphal, R. Cloning and characterization of Pseudomonas aeruginosa fliF, necessary for flagellar assembly and bacterial adherence to mucin. Infect. Immun. 1996, 64, 2130–2136. [Google Scholar]
- Arora, S.K.; Ritchings, B.W.; Almira, E.C.; Lory, S.; Ramphal, R. The Pseudomonas aeruginosa flagellar cap protein, FliD, is responsible for mucin adhesion. Infect. Immun. 1998, 66, 1000–1007. [Google Scholar]
- Lillehoj, E.P.; Kim, B.T.; Kim, K.C. Identification of Pseudomonas aeruginosa flagellin as an adhesin for Muc1 mucin. Am. J. Physiol. Lung C. 2002, 282, L751–L756. [Google Scholar]
- Scharfman, A.; Arora, S.K.; Delmotte, P.; van Brussel, E.; Mazurier, J.; Ramphal, R.; Roussel, P. Recognition of Lewis x derivatives present on mucins by flagellar components of Pseudomonas aeruginosa. Infect. Immun. 2001, 69, 5243–5248. [Google Scholar]
- Bucior, I.; Pielage, J.F.; Engel, J.N. Pseudomonas aeruginosa pili and flagella mediate distinct binding and signaling events at the apical and basolateral surface of airway epithelium. PLoS Pathog. 2012, 8, e1002616. [Google Scholar] [CrossRef]
- Olsen, J.E.; Hoegh-Andersen, K.H.; Casadesus, J.; Rosenkranzt, J.; Chadfield, M.S.; Thomsen, L.E. The role of flagella and chemotaxis genes in host pathogen interaction of the host adapted Salmonella enterica serovar Dublin compared to the broad host range serovar S. Typhimurium. BMC Microbiol. 2013, 13, 67. [Google Scholar] [CrossRef]
- Zgair, A.K.; Chhibber, S. Adhesion of Stenotrophomonas maltophilia to mouse tracheal mucus is mediated through flagella. J. Med. Microbiol. 2011, 60, 1032–1037. [Google Scholar] [CrossRef]
- Hartmann, I.; Carranza, P.; Lehner, A.; Stephan, R.; Eberl, L.; Riedel, K. Genes involved in Cronobacter sakazakii biofilm formation. Appl. Environ. Microbiol. 2010, 76, 2251–2261. [Google Scholar] [CrossRef]
- La Ragione, R.M.; Cooley, W.A.; Woodward, M.J. The role of fimbriae and flagella in the adherence of avian strains of Escherichia coli O78:K80 to tissue culture cells and tracheal and gut explants. J. Med. Microbiol. 2000, 49, 327–338. [Google Scholar]
- La Ragione, R.M.; Sayers, A.R.; Woodward, M.J. The role of fimbriae and flagella in the colonization, invasion and persistence of Escherichia coli O78:K80 in the day-old-chick model. Epidemiol. Infect. 2000, 124, 351–363. [Google Scholar] [CrossRef]
- Best, A.; La Ragione, R.M.; Sayers, A.R.; Woodward, M.J. Role for flagella but not intimin in the persistent infection of the gastrointestinal tissues of specific-pathogen-free chicks by shiga toxin-negative Escherichia coli O157:H7. Infect. Immun. 2005, 73, 1836–1846. [Google Scholar] [CrossRef]
- Luck, S.N.; Badea, L.; Bennett-Wood, V.; Robins-Browne, R.; Hartland, E.L. Contribution of FliC to epithelial cell invasion by enterohemorrhagic Escherichia coli O113:H21. Infect. Immun. 2006, 74, 6999–7004. [Google Scholar] [CrossRef]
- Rogers, T.J.; Thorpe, C.M.; Paton, A.W.; Paton, J.C. Role of lipid rafts and flagellin in invasion of colonic epithelial cells by Shiga-toxigenic Escherichia coli O113:H21. Infect. Immun. 2012, 80, 2858–2867. [Google Scholar] [CrossRef]
- Duan, Q.; Zhou, M.; Liang, H.; Zhu, X.; Guo, Z.; Li, Y.; Hardwidge, P.R.; Zhu, G. Contribution of flagellin subunit FliC to piglet epithelial cells invasion by F18ab E. coli. Vet. Microbiol. 2013. [Google Scholar] [CrossRef]
- Rogers, T.J.; Paton, J.C.; Wang, H.; Talbot, U.M.; Paton, A.W. Reduced virulence of an fliC mutant of Shiga-toxigenic Escherichia coli O113:H21. Infect. Immun. 2006, 74, 1962–1966. [Google Scholar] [CrossRef]
- Allison, C.; Coleman, N.; Jones, P.L.; Hughes, C. Ability of Proteus mirabilis to invade human urothelial cells is coupled to motility and swarming differentiation. Infect. Immun. 1992, 60, 4740–4746. [Google Scholar]
- Mobley, H.L.; Belas, R.; Lockatell, V.; Chippendale, G.; Trifillis, A.L.; Johnson, D.E.; Warren, J.W. Construction of a flagellum-negative mutant of Proteus mirabilis: Effect on internalization by human renal epithelial cells and virulence in a mouse model of ascending urinary tract infection. Infect. Immun. 1996, 64, 5332–5340. [Google Scholar]
- Dibb-Fuller, M.P.; Allen-Vercoe, E.; Thorns, C.J.; Woodward, M.J. Fimbriae- and flagella-mediated association with and invasion of cultured epithelial cells by Salmonella enteritidis. Microbiology 1999, 145, 1023–1031. [Google Scholar] [CrossRef]
- Van Asten, F.J.; Hendriks, H.G.; Koninkx, J.F.; Van Dijk, J.E. Flagella-mediated bacterial motility accelerates but is not required for Salmonella serotype Enteritidis invasion of differentiated Caco-2 cells. Int. J. Med. Microbiol. 2004, 294, 395–399. [Google Scholar] [CrossRef]
- La Ragione, R.M.; Cooley, W.A.; Velge, P.; Jepson, M.A.; Woodward, M.J. Membrane ruffling and invasion of human and avian cell lines is reduced for aflagellate mutants of Salmonella enterica serotype Enteritidis. Int. J. Med. Microbiol. 2003, 293, 261–272. [Google Scholar] [CrossRef]
- Barnich, N.; Boudeau, J.; Claret, L.; Darfeuille-Michaud, A. Regulatory and functional co-operation of flagella and type 1 pili in adhesive and invasive abilities of AIEC strain LF82 isolated from a patient with Crohn’s disease. Mol. Microbiol. 2003, 48, 781–794. [Google Scholar] [CrossRef]
- Dobbin, H.S.; Hovde, C.J.; Williams, C.J.; Minnich, S.A. The Escherichia coli O157 flagellar regulatory gene flhC and not the flagellin gene fliC impacts colonization of cattle. Infect. Immun. 2006, 74, 2894–2905. [Google Scholar] [CrossRef]
- Claret, L.; Miquel, S.; Vieille, N.; Ryjenkov, D.A.; Gomelsky, M.; Darfeuille-Michaud, A. The flagellar sigma factor FliA regulates adhesion and invasion of Crohn disease-associated Escherichia coli via a cyclic dimeric GMP-dependent pathway. J. Biol. Chem. 2007, 282, 33275–33283. [Google Scholar]
- Clyne, M.; Ocroinin, T.; Suerbaum, S.; Josenhans, C.; Drumm, B. Adherence of isogenic flagellum-negative mutants of Helicobacter pylori and Helicobacter mustelae to human and ferret gastric epithelial cells. Infect. Immun. 2000, 68, 4335–4339. [Google Scholar] [CrossRef]
- Guentzel, M.N.; Berry, L.J. Motility as a virulence factor for Vibrio cholerae. Infect. Immun. 1975, 11, 890–897. [Google Scholar]
- Zunino, P.; Piccini, C.; Legnani-Fajardo, C. Flagellate and non-flagellate Proteus mirabilis in the development of experimental urinary tract infection. Microb. Pathog. 1994, 16, 379–385. [Google Scholar]
- Legnani-Fajardo, C.; Zunino, P.; Piccini, C.; Allen, A.; Maskell, D. Defined mutants of Proteus mirabilis lacking flagella cause ascending urinary tract infection in mice. Microb. Pathogenesis 1996, 21, 395–405. [Google Scholar] [CrossRef]
- Armbruster, C.E.; Mobley, H.L. Merging mythology and morphology: The multifaceted lifestyle of Proteus mirabilis. Nat. Rev. Microbiol. 2012, 10, 743–754. [Google Scholar] [CrossRef]
- Sheu, B.S.; Yang, H.B.; Yeh, Y.C.; Wu, J.J. Helicobacter pylori colonization of the human gastric epithelium: A bug’s first step is a novel target for us. J. Gastroen. Hepatol. 2010, 25, 26–32. [Google Scholar] [CrossRef]
- Kim, J.S.; Chang, J.H.; Chung, S.I.; Yum, J.S. Molecular cloning and characterization of the Helicobacter pylori fliD gene, an essential factor in flagellar structure and motility. J. Bacteriol. 1999, 181, 6969–6976. [Google Scholar]
- Croxen, M.A.; Finlay, B.B. Molecular mechanisms of Escherichia coli pathogenicity. Nat. Rev. Microbiol. 2010, 8, 26–38. [Google Scholar]
- Fairbrother, J.M.; Nadeau, E.; Gyles, C.L. Escherichia coli in postweaning diarrhea in pigs: An update on bacterial types, pathogenesis, and prevention strategies. Anim. Health Res. Rev. 2005, 6, 17–39. [Google Scholar] [CrossRef]
- Tivendale, K.A.; Logue, C.M.; Kariyawasam, S.; Jordan, D.; Hussein, A.; Li, G.; Wannemuehler, Y.; Nolan, L.K. Avian-pathogenic Escherichia coli strains are similar to neonatal meningitis E. coli strains and are able to cause meningitis in the rat model of human disease. Infect. Immun. 2010, 78, 3412–3419. [Google Scholar] [CrossRef]
- Köhler, C.D.; Dobrindt, U. What defines extraintestinal pathogenic Escherichia coli? Int. J. Med. Microbiol. 2011, 301, 642–647. [Google Scholar] [CrossRef]
- Clements, A.; Young, J.C.; Constantinou, N.; Frankel, G. Infection strategies of enteric pathogenic Escherichia coli. Gut Microbes 2012, 3, 71–87. [Google Scholar] [CrossRef]
- Wang, L.; Rothemund, D.; Curd, H.; Reeves, P.R. Species-wide variation in the Escherichia coli flagellin (H-antigen) gene. J. Bacteriol. 2003, 185, 2936–2943. [Google Scholar] [CrossRef]
- Bonacorsi, S.; Bingen, E. Molecular epidemiology of Escherichia coli causing neonatal meningitis. Int. J. Med. Microbiol. 2005, 295, 373–381. [Google Scholar] [CrossRef]
- Xie, Y.; Parthasarathy, G.; Di Cello, F.; Teng, C.H.; Paul-Satyaseela, M.; Kim, K.S. Transcriptome of Escherichia coli K1 bound to human brain microvascular endothelial cells. Biochem. Biophys. Res. Commun. 2008, 365, 201–206. [Google Scholar] [CrossRef]
- Teng, C.H.; Xie, Y.; Shin, S.; di Cello, F.; Paul-Satyaseela, M.; Cai, M.; Kim, K.S. Effects of ompA deletion on expression of type 1 fimbriae in Escherichia coli K1 strain RS218 and on the association of E. coli with human brain microvascular endothelial cells. Infect. Immun. 2006, 74, 5609–5616. [Google Scholar] [CrossRef]
- Human Microbiome Project Consortium. Structure, function and diversity of the healthy human microbiome. Nature 2012, 486, 207–214. [Google Scholar] [CrossRef] [Green Version]
- Arora, S.K.; Dasgupta, N.; Lory, S.; Ramphal, R. Identification of two distinct types of flagellar cap proteins, FliD, in Pseudomonas aeruginosa. Infect. Immun. 2000, 68, 1474–1479. [Google Scholar] [CrossRef]
- Ternan, N.G.; Jain, S.; Srivastava, M.; McMullan, G. Comparative transcriptional analysis of clinically relevant heat stress response in Clostridium difficile strain 630. PLoS One 2012, 7, e42410. [Google Scholar]
- Jones, A.L.; Beveridge, T.J.; Woods, D.E. Intracellular survival of Burkholderia pseudomallei. Infect. Immun. 1996, 64, 782–790. [Google Scholar]
- Inglis, T.J.; Rigby, P.; Robertson, T.A.; Dutton, N.S.; Henderson, M.; Chang, B.J. Interaction between Burkholderia pseudomallei and Acanthamoeba species results in coiling phagocytosis, endamebic bacterial survival, and escape. Infect. Immun. 2000, 68, 1681–1686. [Google Scholar] [CrossRef]
- Chuaygud, T.; Tungpradabkul, S.; Sirisinha, S.; Chua, K.L.; Utaisincharoen, P. A role of Burkholderia pseudomallei flagella as a virulent factor. T. Roy. Soc. Trop. Med. H. 2008, 102, S140–S144. [Google Scholar] [CrossRef]
- Wajanarogana, S.; Sonthayanon, P.; Wuthiekanun, V.; Panyim, S.; Simpson, A.J.; Tungpradabkul, S. Stable marker on flagellin gene sequences related to arabinose non-assimilating pathogenic Burkholderia pseudomallei. Microbiol. Immunol. 1999, 43, 995–1001. [Google Scholar]
- DeShazer, D.; Brett, P.J.; Carlyon, R.; Woods, D.E. Mutagenesis of Burkholderia pseudomallei with Tn5-OT182: Isolation of motility mutants and molecular characterization of the flagellin structural gene. J. Bacteriol. 1997, 179, 2116–2125. [Google Scholar]
- Tomich, M.; Herfst, C.A.; Golden, J.W.; Mohr, C.D. Role of flagella in host cell invasion by Burkholderia cepacia. Infect. Immun. 2002, 70, 1799–1806. [Google Scholar] [CrossRef]
- Lee, J.H.; Rho, J.B.; Park, K.J.; Kim, C.B.; Han, Y.S.; Choi, S.H.; Lee, K.H.; Park, S.J. Role of flagellum and motility in pathogenesis of Vibrio vulnificus. Infect. Immun. 2004, 72, 4905–4910. [Google Scholar] [CrossRef]
- Wassenaar, T.M.; Bleumink-Pluym, N.M.; van der Zeijst, B.A. Inactivation of Campylobacter jejuni flagellin genes by homologous recombination demonstrates that flaA but not flaB is required for invasion. EMBO J. 1991, 10, 2055–2061. [Google Scholar]
- Grant, C.C.; Konkel, M.E.; Cieplak, W., Jr.; Tompkins, L.S. Role of flagella in adherence, internalization, and translocation of Campylobacter jejuni in nonpolarized and polarized epithelial cell cultures. Infect. Immun. 1993, 61, 1764–1771. [Google Scholar]
- Song, Y.C.; Jin, S.; Louie, H.; Ng, D.; Lau, R.; Zhang, Y.; Weerasekera, R.; Al Rashid, S.; Ward, L.A.; Der, S.D.; Chan, V.L. FlaC, a protein of Campylobacter jejuni TGH9011 (ATCC43431) secreted through the flagellar apparatus, binds epithelial cells and influences cell invasion. Mol. Microbiol. 2004, 53, 541–553. [Google Scholar] [CrossRef]
- Friedlander, R.S.; Vlamakis, H.; Kim, P.; Khan, M.; Kolter, R.; Aizenberg, J. Bacterial flagella explore microscale hummocks and hollows to increase adhesion. Proc. Natl. Acad. Sci. USA 2013, 110, 5624–5629. [Google Scholar]
- Crawford, R.W.; Reeve, K.E.; Gunn, J.S. Flagellated but not hyperfimbriated Salmonella enterica serovar Typhimurium attaches to and forms biofilms on cholesterol-coated surfaces. J. Bacteriol. 2010, 192, 2981–2990. [Google Scholar] [CrossRef]
- Subramanian, N.; Qadri, A. Lysophospholipid sensing triggers secretion of flagellin from pathogenic salmonella. Nat. Immunol. 2006, 7, 583–589. [Google Scholar] [CrossRef]
- McNamara, N.; Khong, A.; McKemy, D.; Caterina, M.; Boyer, J.; Julius, D.; Basbaum, C. ATP transduces signals from ASGM1, a glycolipid that functions as a bacterial receptor. Proc. Natl. Acad. Sci. USA 2001, 98, 9086–9091. [Google Scholar]
- Adamo, R.; Sokol, S.; Soong, G.; Gomez, M.I.; Prince, A. Pseudomonas aeruginosa flagella activate airway epithelial cells through asialoGM1 and toll-like receptor 2 as well as toll-like receptor 5. Am. J. Respir. Cell Mol. 2004, 30, 627–634. [Google Scholar] [CrossRef]
- Ogushi, K.; Wada, A.; Niidome, T.; Okuda, T.; Llanes, R.; Nakayama, M.; Nishi, Y.; Kurazono, H.; Smith, K.D.; Aderem, A.; et al. Gangliosides act as co-receptors for Salmonella enteritidis FliC and promote FliC induction of human beta-defensin-2 expression in Caco-2 cells. J. Biol. Chem. 2004, 279, 12213–12219. [Google Scholar]
- McNamara, N.; Gallup, M.; Sucher, A.; Maltseva, I.; McKemy, D.; Basbaum, C. AsialoGM1 and TLR5 cooperate in flagellin-induced nucleotide signaling to activate Erk1/2. Am. J. Respir. Cell Mol. 2006, 34, 653–660. [Google Scholar] [CrossRef]
- Ishii, S.; Kosaka, T.; Hori, K.; Hotta, Y.; Watanabe, K. Coaggregation facilitates interspecies hydrogen transfer between Pelotomaculum thermopropionicum and Methanothermobacter thermautotrophicus. Appl. Environ. Microbiol. 2005, 71, 7838–7845. [Google Scholar] [CrossRef]
- Shimoyama, T.; Kato, S.; Ishii, S.; Watanabe, K. Flagellum mediates symbiosis. Science 2009, 323, 1574. [Google Scholar] [CrossRef]
- Döring, G.; Meisner, C.; Stern, M.; Flagella Vaccine Trial Study Group. A double-blind randomized placebo-controlled phase III study of a Pseudomonas aeruginosa flagella vaccine in cystic fibrosis patients. Proc. Natl. Acad. Sci. USA 2007, 104, 11020–11025. [Google Scholar]
- McNeilly, T.N.; Naylor, S.W.; Mahajan, A.; Mitchell, M.C.; McAteer, S.; Deane, D.; Smith, D.G.E.; Low, J.C.; Gally, D.L.; Huntley, J.F. Escherichia coli O157:H7 colonization in cattle following systemic and mucosal immunization with purified H7 flagellin. Infect. Immun. 2008, 76, 2594–2602. [Google Scholar] [CrossRef]
- Gat, O.; Galen, J.E.; Tennant, S.; Simon, R.; Blackewelder, W.C.; Silverman, D.J.; Pasetti, M.F.; Levine, M.M. Cell-associated flagella enhance the protection conferred by mucosally-administered attenuated Salmonella Paratyphi A vaccines. PLoS Neglect. Trop. D. 2011, 5, e1373. [Google Scholar] [CrossRef]
- Wajanarogana, S.; Nimnuch, P.; Thongmee, A.; Kritsiriwuthinan, K. Potential of recombinant flagellin fragment from Burkholderia thailandensis as an antigen for melioidosis antibody detection by indirect ELISA. Mol. Cell. Probes 2013, 27, 98–102. [Google Scholar] [CrossRef]
- Burdelya, L.G.; Krivokrysenko, V.I.; Tallant, T.C.; Strom, E.; Gleiberman, A.S.; Gupta, D.; Kurnasov, O.V.; Fort, F.L.; Osterman, A.L.; Didonato, J.A.; et al. An agonist of Toll Like Receptor 5 has radioprotective activity in mouse and primate models. Science 2008, 320, 226–230. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Haiko, J.; Westerlund-Wikström, B. The Role of the Bacterial Flagellum in Adhesion and Virulence. Biology 2013, 2, 1242-1267. https://doi.org/10.3390/biology2041242
Haiko J, Westerlund-Wikström B. The Role of the Bacterial Flagellum in Adhesion and Virulence. Biology. 2013; 2(4):1242-1267. https://doi.org/10.3390/biology2041242
Chicago/Turabian StyleHaiko, Johanna, and Benita Westerlund-Wikström. 2013. "The Role of the Bacterial Flagellum in Adhesion and Virulence" Biology 2, no. 4: 1242-1267. https://doi.org/10.3390/biology2041242
APA StyleHaiko, J., & Westerlund-Wikström, B. (2013). The Role of the Bacterial Flagellum in Adhesion and Virulence. Biology, 2(4), 1242-1267. https://doi.org/10.3390/biology2041242