Micro-Eukaryotic Diversity in Hypolithons from Miers Valley, Antarctica

Abstract
:1. Introduction
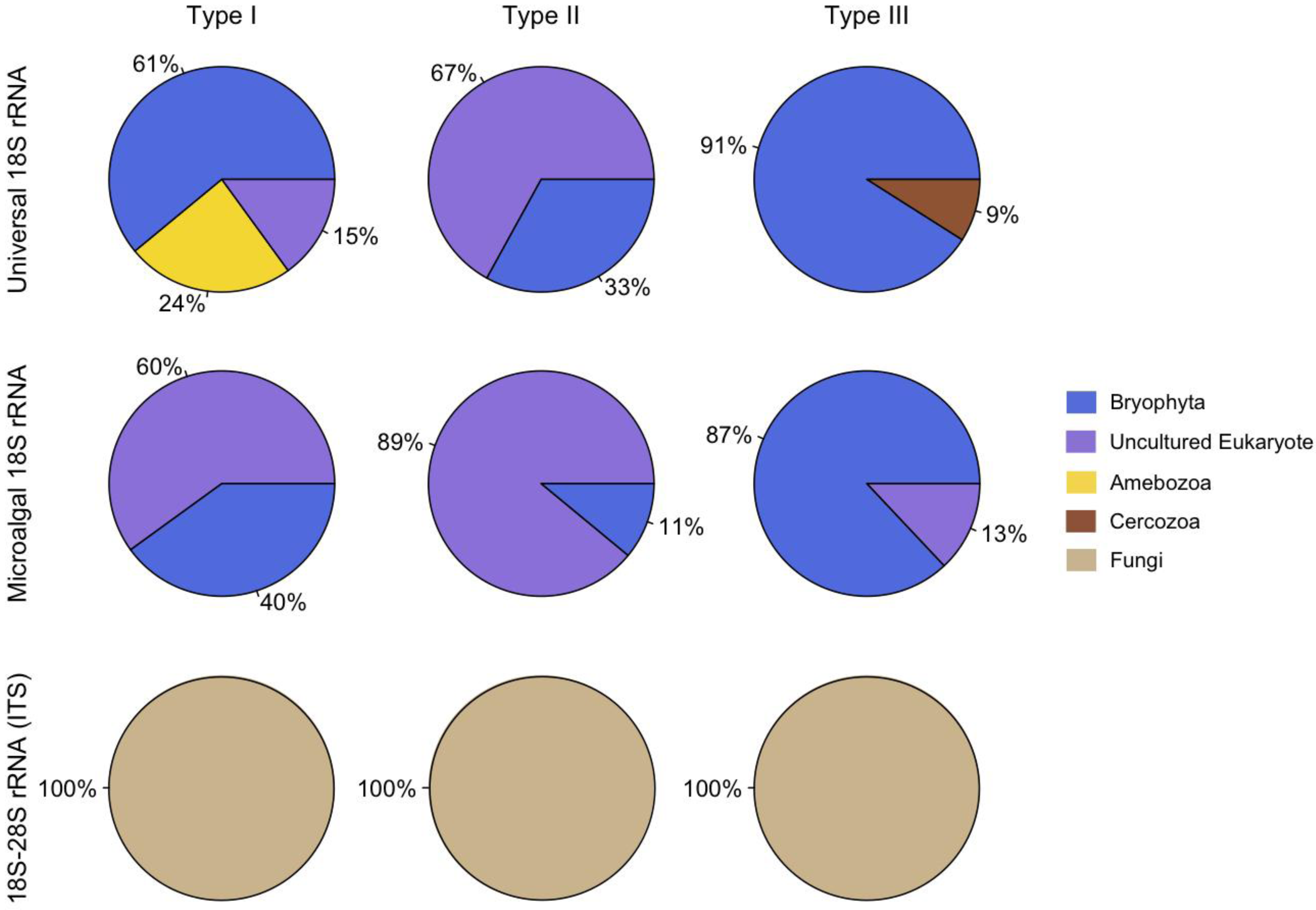
2. Results and Discussion

{kind=link}
| Primer Set | Sequence (5’- 3’) | Region of Amplification | PCR Parameters | Reference |
|---|---|---|---|---|
| EukA | AACCTGGTTGATCCTGCCAGT | 18S rRNA gene | 94 °C for 3 min; 30 cycles: 94 °C for 45 s, 50 °C for 1 min, 72 °C for 3 min; 72 °C for 20 min | [20] |
| EukB | TGATCCTTCTGCAGGTTCACCTAC | |||
| ITS1F | CTTGGTCATTTAGAGGAAGTAATC | ITS1-ITS2 | 94 °C for 5 min; 35 cycles: 94 °C for 1 min, 50 °C for 1 min, 72 °C for 1 min; 72 °C for 20 min | [21,22] |
| ITS4 | CTCCGCTTATTGATATGC | |||
| P45 | ACCTGGTTGATCCTGCCAGT | Microalgal 18S | 94 °C for 1 min; 37 cycles: 92 °C for 50 s, 57 °C for 50 s, 72°C for 50 s; 72°C for 10 min | [23] |
| P47 | TCTCAGGCTCCCTCTCCGGA | rRNA gene |
| Representative Clone | Accession No. | Closest Sequence Match | Accession No. | Identity | Typea |
|---|---|---|---|---|---|
| Eukaryote 18S rRNA | |||||
| Euk75-A1 | KC352912 | Uncultured Eucalypta (Bryophyta) | Y17871 | 81% | 2/I |
| Euk75-A7 | KC352913 | Uncultured Tortula ruralis (Bryophyta) | AF023682 | 78% | 2/I |
| Euk75-A8 | KC352914 | Uncultured Tortula ruralis (Bryophyta) | AF023682 | 86% | 2/I |
| Euk75-A12 | KC352915 | Uncultured Tortula ruralis (Bryophyta) | AF023682 | 86% | 2/I |
| Euk75-B2 | KC352916 | Uncultured Tortula ruralis (Bryophyta) | AF023682 | 92% | 2/I |
| Euk75-B9 | KC352917 | Saccamoeba limax (Amoebozoa) | AF293902 | 98% | 2/I |
| Euk75-C4 | KC352918 | Platyamoeba contorta (Amoebozoa) | DQ229954 | 96% | 2/I |
| Euk134-C6 | KC352919 | Pottia truncata (Bryophyta) | X95935 | 99% | 4/II |
| Euk134-D11 | KC352920 | Uncultured eukaryote | HM490274 | 100% | 4/II |
| Euk50-B10 | KC352921 | Uncultured eukaryote | EF024087 | 91% | 5/III |
| Euk50-D10 | KC352922 | Cercomonas plasmodialis (Cercozoa) | AF411268 | 99% | 5/III |
| Microalgal 18S RNA | |||||
| P50-A4 | KC352936 | Mnium hornum (Bryophyta) | X80985 | 95% | 5/III |
| P50-B3 | KC352937 | Uncultured eukaryote | EF024845 | 99% | 5/III |
| P50-B6 | KC352938 | Mnium hornum (Bryophyta) | X80985 | 91% | 5/III |
| P50-E7 | KC352939 | Uncultured eukaryote | EF526889 | 99% | 5/III |
| P75-E | KC352940 | Bryoxiphium norvegicum (Bryophyta) | AF223008 | 85% | 2/I |
| P134-A2 | KC352941 | Uncultured eukaryote | FN394778 | 100% | 4/II |
| P134-A11 | KC352942 | Uncultured eukaryote | HM490274 | 100% | 4/II |
| IGS 28S-18S rRNA | |||||
| ITS65-A1 | KC352923 | Uncultured Acremonium (Ascomycota) | HE977538 | 82% | 1/I |
| ITS65-A2 | KC352924 | Uncultured Acremonium (Ascomycota) | HE977544 | 85% | 1/I |
| ITS65-A5 | KC352925 | Uncultured Acremonium (Ascomycota) | HE977544 | 85% | 1/I |
| ITS65-A8 | KC352926 | Uncultured Acremonium (Ascomycota) | HE977544 | 78% | 1/I |
| ITS65-B6 | KC352927 | Uncultured Acremonium (Ascomycota) | HE977544 | 88% | 1/I |
| ITS65-C12 | KC352928 | Uncultured Acremonium (Ascomycota) | HE977544 | 88% | 1/I |
| ITS65-D2 | KC352929 | Uncultured Acremonium (Ascomycota) | HE977544 | 87% | 1/I |
| ITS65-D10 | KC352930 | Stromatonectria caraganae (Ascomycota) | HQ112288 | 80% | 1/I |
| ITS134-A1 | KC352931 | Verrucaria sp. (Ascomycota) | FJ664858 | 92% | 2/II |
| ITS50-B11 | KC352932 | Uncultured Acremonium (Ascomycota) | HE977544 | 84% | 5/III |
| ITS50-D2 | KC352933 | Uncultured Acremonium (Ascomycota) | HE977538 | 80% | 5/III |
| ITS50-E3 | KC352934 | Uncultured Acremonium (Ascomycota) | HE977538 | 86% | 5/III |
| ITS50-H7 | KC352935 | Uncultured Acremonium (Ascomycota) | HE977538 | 91% | 5/III |
3. Experimental Section
3.1. Samples Collection
3.2. DNA Extractions and PCR Amplifications
3.3. Clone Library Construction and Phylogenetic Analysis
4. Conclusions
Acknowledgments
References
- Cary, S.C.; McDonald, I.R.; Barrett, J.E.; Cowan, D.A. On the rocks: The microbiology of Antarctic Dry Valley soils. Nat. Rev. Microbiol. 2010, 8, 129–138. [Google Scholar] [CrossRef]
- Franzmann, P.D. Examination of Antarctic prokaryotic diversity through molecular comparisons. Biodivers. Conser. 1996, 5, 1295–1305. [Google Scholar] [CrossRef]
- Campanaro, S.; Williams, T.J.; Burg, D.W.; De Francisci, D.; Treu, L.; Lauro, F.M.; Cavicchioli, R. Temperature-dependent global gene expression in the Antarctic archaeon Methanococcoides burtonii. Environ. Microbiol. 2011, 13, 2018–2038. [Google Scholar] [CrossRef]
- Massana, R.; Taylor, L.J.; Murray, A.E.; Wu, K.Y.; Jeffrey, W.H.; DeLong, E.F. Vertical distribution and temporal variation of marine planktonic archaea in the Gerlache Strait, Antarctica, during early spring. Limnol. Oceanogr. 1998, 43, 607–617. [Google Scholar] [CrossRef]
- Gardner, H.; Kerry, K.; Riddle, M.; Brouwer, S.; Gleeson, L. Poultry virus infection in Antarctic penguins. Nature 1997, 387, 245–245. [Google Scholar]
- Yau, S.; Lauro, F.M.; DeMaere, M.Z.; Brown, M.V.; Thomas, T.; Raftery, M.J.; Andrews-Pfannkoch, C.; Lewis, M.; Hoffman, J.M.; Gibson, J.A.; Cavicchioli, R. Virophage control of antarctic algal host-virus dynamics. Proc. Natl. Acad. Sci. USA 2011, 108, 6163–6168. [Google Scholar]
- Fell, J.W.; Scorzetti, G.; Connell, L.; Craig, S. Biodiversity of micro-eukaryotes in Antarctic Dry Valley soils with <5% soil moisture. Soil Biol. Biochem. 2006, 38, 3107–3119. [Google Scholar] [CrossRef]
- Cowan, D.A. Cryptic microbial communities in Antarctic deserts. Proc. Natl. Acad. Sci. USA 2009, 106, 19749–19750. [Google Scholar]
- Cowan, D.A.; Khan, N.; Pointing, S.B.; Cary, S.C. Diverse hypolithic refuge communities in the McMurdo Dry Valleys. Antarc. Sci. 2010, 22, 714–720. [Google Scholar] [CrossRef]
- Cowan, D.A.; Pointing, S.B.; Stevens, M.I.; Cary, S.C.; Stomeo, F.; Tuffin, I.M. Distribution and abiotic influences on hypolithic microbial communities in an Antarctic Dry Valley. Polar Biol. 2011, 34, 307–311. [Google Scholar] [CrossRef]
- Khan, N.; Tuffin, M.; Stafford, W.; Cary, C.; Lacap, D.C.; Pointing, S.B.; Cowan, D. Hypolithic microbial communities of quartz rocks from Miers Valley, McMurdo Dry Valleys, Antarctica. Polar Biol. 2011, 34, 1657–1668. [Google Scholar] [CrossRef]
- Chan, Y.; Lacap, D.C.; Lau, M.C.Y.; Ha, K.Y.; Warren-Rhodes, K.A.; Cockell, C.S.; Cowan, D.A.; McKay, C.P.; Pointing, S.B. Hypolithic microbial communities: Between a rock and a hard place. Environ. Microbiol. 2012. [Google Scholar]
- Pointing, S.B.; Chan, Y.; Lacap, D.C.; Lau, M.C.Y.; Jurgens, J.A.; Farrell, R.L. Highly specialized microbial diversity in hyper-arid polar desert. Proc. Natl. Acad. Sci. USA 2009, 106, 19964–19969. [Google Scholar]
- Cockell, C.S.; Stokes, M.D. Widespread colonization by polar hypoliths. Nature 2004, 431, 414. [Google Scholar] [CrossRef]
- Cowan, D.A.; Sohm, J.A.; Makhalanyane, T.P.; Capone, D.G.; Green, T.G.A.; Cary, S.C.; Tuffin, I.M. Hypolithic communities: important nitrogen sources in Antarctic desert soils. Environ. Microbiol. Rep. 2011, 3, 581–586. [Google Scholar] [CrossRef]
- Potvin, M.; Lovejoy, C. PCR-Based Diversity Estimates of Artificial and Environmental 18S rRNA Gene Libraries. J. Eukaryot. Microbiol. 2009, 56, 174–181. [Google Scholar] [CrossRef]
- Jones, C.G.; Lawton, J.H.; Shachak, M. Organisms as ecosystem engineers. Oikos 1994, 69, 373–386. [Google Scholar] [CrossRef]
- Pointing, S.B.; Belnap, J. Microbial colonization and controls in dryland systems. Nat. Rev. Microbiol. 2012, 10, 551–562. [Google Scholar] [CrossRef]
- Zakhia, F.; Jungblut, A.; Taton, A.; Vincent, W.; Wilmotte, A. Cyanobacteria in cold environments. In Psychrophiles: From Biodiversity to Biotechnology; Margesin, R.S.F., Marx, J.C., Gerday, C., Eds.; Springer-Verlag: Berlin, 2007; pp. 121–135. [Google Scholar]
- Diez, B.; Pedros-Alio, C.; Marsh, T.L.; Massana, R. Application of denaturing gradient gel electrophoresis (DGGE) to study the diversity of marine picoeukaryotic assemblages and comparison of DGGE with other molecular techniques. Appl. Environ. Microbiol. 2001, 67, 2942–2951. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T.D. Its primers with enhanced specificity for basidiomycetes - application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990. [Google Scholar]
- Dorigo, U.; Berard, A.; Humbert, J.F. Comparison of eukaryotic phytobenthic community composition in a polluted river by partial 18S rRNA gene cloning and sequencing. Microb. Ecol. 2002, 44, 372–380. [Google Scholar] [CrossRef]
- Arenz, B.E.; Held, B.W.; Jurgens, J.A.; Farrell, R.L.; Blanchette, R.A. Fungal diversity in soils and historic wood from the Ross Sea Region of Antarctica. Soil Biol. Biochem. 2006, 38, 3057–3064. [Google Scholar] [CrossRef]
- Bridge, P.D.; Spooner, B.M. Non-lichenized Antarctic fungi: transient visitors or members of a cryptic ecosystem? Fungal Ecol. 2012, 5, 381–394. [Google Scholar] [CrossRef]
- Bridge, P.; Spooner, B.; Roberts, P. List of Non-lichenized Fungi from the Antarctic Region. Available online: http://www.antarctica.ac.uk/bas_research/data/access/fungi/ (Accessed on 10 December 2012).
- Arenz, B.E.; Held, B.W.; Jurgens, J.A.; Blanchette, R.A. Fungal colonization of exotic substrates in Antarctica. Fungal Divers. 2011, 49, 13–22. [Google Scholar] [CrossRef]
- Wynn-Wmilliams, D.D. Seasonal fluctuations in microbial activity in antarctic moss peat. Biol. J. Linn. Soc. 1980, 14, 11–28. [Google Scholar] [CrossRef]
- Onofri, S.; Zucconi, L.; Tosi, S. Continental Antarctic Fungi; IHW-Verlag: Munich, Germany, 2007; p. 247. [Google Scholar]
- Bamforth, S.S.; Wall, D.H.; Virginia, R.A. Distribution and diversity of soil protozoa in the McMurdo Dry Valleys of Antarctica. Polar Biol. 2005, 28, 756–762. [Google Scholar] [CrossRef]
- Nakai, R.; Abe, T.; Baba, T.; Imura, S.; Kagoshima, H.; Kanda, H.; Kohara, Y.; Koi, A.; Niki, H.; Yanagihara, K.; Naganuma, T. Eukaryotic phylotypes in aquatic moss pillars inhabiting a freshwater lake in East Antarctica, based on 18S rRNA gene analysis. Polar Biol. 2012, 35, 1495–1504. [Google Scholar] [CrossRef]
- Calbet, A.; Landry, M.R. Phytoplankton growth, microzooplankton grazing, and carbon cycling in marine systems. Limnol. Oceanogr. 2004, 49, 51–57. [Google Scholar]
- Elliott, E.T.; Anderson, R.V.; Coleman, D.C.; Cole, C.V. Habitable pore-space and microbial trophic interactions. Oikos 1980, 35, 327–335. [Google Scholar] [CrossRef]
- Wardle, D. Communities and Ecosystems: Linking the Aboveground and Belowground Components; Princeton University Press: Princeton, USA, 2002; p. 400. [Google Scholar]
- Lange, O.L.; Kidron, G.J.; Budel, B.; Meyer, A.; Kilian, E.; Abeliovich, A. Taxonomic composition and photosynthetic characteristics of the biological soil crusts covering sand dunes in the western negev desert. Funct. Ecol. 1992, 6, 519–527. [Google Scholar] [CrossRef]
- Mues, R. Chemical constituents and biochemistry. In Bryophyte Biology; Shaw, A.J., Goffinet, B., Eds.; Cambridge University Press: Cambridge, UK, 2000; pp. 150–181. [Google Scholar]
- Liu, L.-C.; Li, S.-Z.; Duan, Z.-H.; Wang, T.; Zhang, Z.-S.; Li, X.-R. Effects of microbiotic crusts on dew deposition in the restored vegetation area at Shapotou, northwest China. J. Hydrol. 2006, 328, 331–337. [Google Scholar] [CrossRef]
- Proctor, M.C.F. The bryophyte paradox: Tolerance of desiccation, evasion of drought. Plant Ecol. 2000, 151, 41–49. [Google Scholar] [CrossRef]
- Von Sigler, W. DNA Extraction from soil, sediment and plant tissue: 50–50–50 buffer-chloroform/phenol method. Available online: http://www.eeescience.utoledo.edu/Faculty/Sigler/-Von_Sigler/LEPR_Protocols_files/DNA%20extraction%20-%20soil.pdf (Accessed on DD/ MM/ YY).
- Huber, T.; Faulkner, G.; Hugenholtz, P. Bellerophon: A program to detect chimeric sequences in multiple sequence alignments. Bioinformatics 2004, 20, 2317–2319. [Google Scholar]
- Huang, Y.; Niu, B.; Gao, Y.; Fu, L.; Li, W. CD-HIT Suite: A web server for clustering and comparing biological sequences. Bioinformatics 2010, 26, 680–682. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Gokul, J.K.; Valverde, A.; Tuffin, M.; Cary, S.C.; Cowan, D.A. Micro-Eukaryotic Diversity in Hypolithons from Miers Valley, Antarctica. Biology 2013, 2, 331-340. https://doi.org/10.3390/biology2010331
Gokul JK, Valverde A, Tuffin M, Cary SC, Cowan DA. Micro-Eukaryotic Diversity in Hypolithons from Miers Valley, Antarctica. Biology. 2013; 2(1):331-340. https://doi.org/10.3390/biology2010331
Chicago/Turabian StyleGokul, Jarishma K., Angel Valverde, Marla Tuffin, Stephen Craig Cary, and Don A. Cowan. 2013. "Micro-Eukaryotic Diversity in Hypolithons from Miers Valley, Antarctica" Biology 2, no. 1: 331-340. https://doi.org/10.3390/biology2010331
APA StyleGokul, J. K., Valverde, A., Tuffin, M., Cary, S. C., & Cowan, D. A. (2013). Micro-Eukaryotic Diversity in Hypolithons from Miers Valley, Antarctica. Biology, 2(1), 331-340. https://doi.org/10.3390/biology2010331