2,4-Epibrassinolide Mitigates Cd Stress by Enhancing Chloroplast Structural Remodeling and Chlorophyll Metabolism in Vigna angularis Leaves

 and
and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials, Growth, and Treatment Conditions
2.2. Biomass and Morphological Measurements
2.3. Cadmium (Cd) Content Measurement
2.4. Photosynthetic Pigment Content and Chlorophyll Fluorescence Measurement
2.5. Determination of Leaf Microstructure
2.6. ROS, MDA, and Antioxidant Contents and the Activity of Antioxidant Enzymes
2.7. Osmoregulatory Substance Content
2.8. RNA Extraction, First-Strand cDNA Synthesis, and qRT-PCR Analysis
2.9. RNA-seq Analysis
2.10. Statistical Analysis
3. Results
3.1. BR Treatment Alleviates the Inhibition of V. angularis growth Under Cd Stress
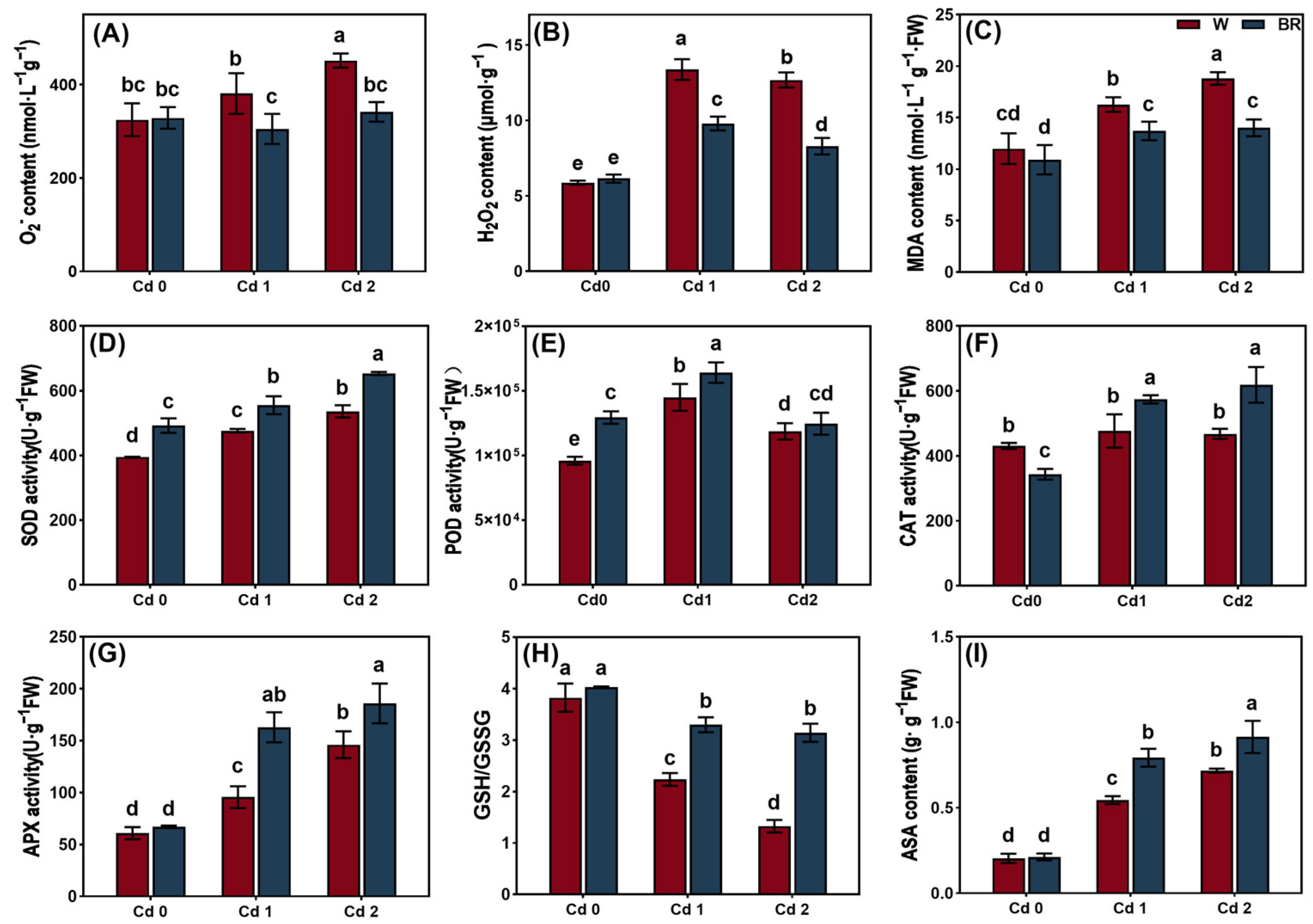
3.2. BR Treatment Reduced the Oxidative Damage Induced by Cd in the V. angularis Leaves
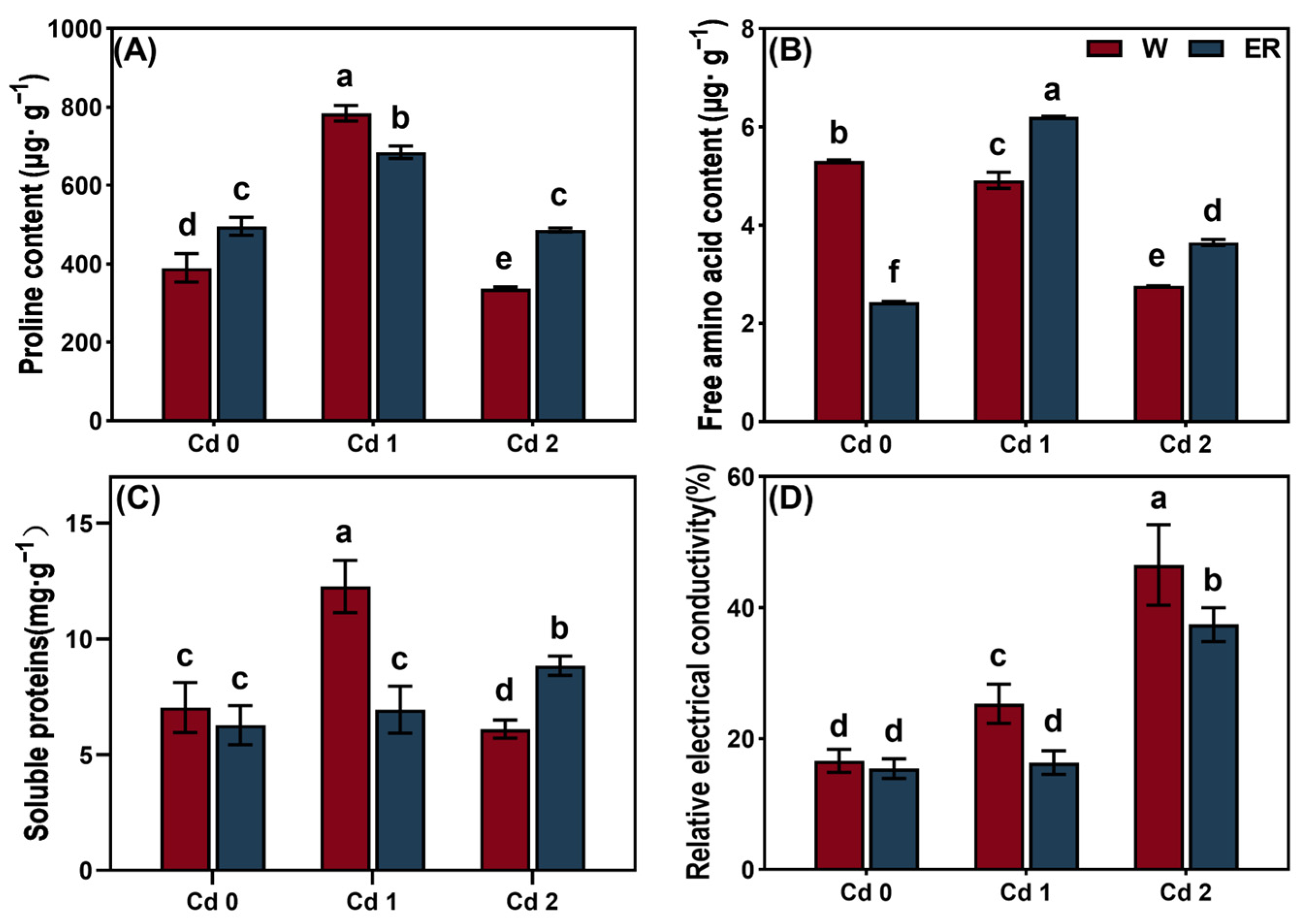
3.3. BR Treatment Regulated the Osmotic Regulation of V. angularis Leaves Under Cd Stress
3.4. BR Treatment Regulated Chlorophyll Content and Chlorophyll Fluorescence of V. angularis Leaves Under Cd Stress
3.5. BR Treatment Improved Microstructure of V. angularis Leaves Under Cd Stress
3.6. Effect of BR Treatment on Transcription of Genes in V. angularis Leaves Under Cd Stress
3.7. The BR Treatment Regulated the Photosynthetic Capacity of V. angularis Leaves Under Cd Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Thévenod, F.; Lee, W.K. Toxicology of cadmium and its damage to mammalian organs. Met. Ions Life Sci. 2013, 11, 415–490. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.C.; Zhang, Q.C.; Yan, C.A.; Tang, G.Y.; Zhang, M.Y.; Ma, L.Q.; Gu, R.H.; Xiang, P. Heavy metal(loid)s in agriculture soils, rice, and wheat across China: Status assessment and spatiotemporal analysis. Sci. Total Environ. 2023, 882, 163361. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Hu, J.; Yu, Y.; Cheng, X.; Du, Y.; Zhao, Q.; Du, J. Interactive effects of drought and cadmium stress on adzuki bean seedling growth, DNA damage repair, and Cd accumulation. Sci. Hortic. 2024, 324, 112624. [Google Scholar] [CrossRef]
- Sarwar, N.; Imran, M.; Shaheen, M.R.; Ishaque, W.; Kamran, M.A.; Matloob, A.; Rehim, A.; Hussain, S. Phytoremediation strategies for soils contaminated with heavy metals: Modifications and future perspectives. Chemosphere 2017, 171, 710–721. [Google Scholar] [CrossRef]
- Rafique, M.; Ortas, I.; Rizwan, M.; Sultan, T.; Chaudhary, H.J.; Işik, M.; Aydin, O. Effects of Rhizophagus clarus and biochar on growth, photosynthesis, nutrients, and cadmium (Cd) concentration of maize (Zea mays) grown in Cd-spiked soil. Environ. Sci. Pollut. Res. Int. 2019, 26, 20689–20700. [Google Scholar] [CrossRef]
- Zhao, Q.; Wang, H.; Du, Y.; Rogers, H.J.; Wu, Z.; Jia, S.; Yao, X.; Xie, F.; Liu, W. MSH2 and MSH6 in Mismatch Repair System Account for Soybean (Glycine max (L.) Merr.) Tolerance to Cadmium Toxicity by Determining DNA Damage Response. J. Agric. Food Chem. 2020, 68, 1974–1985. [Google Scholar] [CrossRef]
- Wang, M.; Duan, S.; Zhou, Z.; Chen, S.; Wang, D. Foliar spraying of melatonin confers cadmium tolerance in Nicotiana tabacum L. Ecotoxicol. Environ. Saf. 2019, 170, 68–76. [Google Scholar] [CrossRef]
- Wang, H.; Cao, Q.; Zhao, Q.; Arfan, M.; Liu, W. Mechanisms used by DNA MMR system to cope with cadmium-induced DNA damage in plants. Chemosphere 2020, 246, 125614. [Google Scholar] [CrossRef]
- Belyaeva, E.A.; Dymkowska, D.; Wieckowski, M.R.; Wojtczak, L. Mitochondria as an important target in heavy metal toxicity in rat hepatoma AS-30D cells. Toxicol. Appl. Pharmacol. 2008, 231, 34–42. [Google Scholar] [CrossRef]
- Mansoor, S.; Ali, A.; Kour, N.; Bornhorst, J.; AlHarbi, K.; Rinklebe, J.; Abd El Moneim, D.; Ahmad, P.; Chung, Y.S. Heavy metal Induced oxidative stress mitigation and ROS scavenging in plants. Plants 2023, 12, 3003. [Google Scholar] [CrossRef]
- Luo, S.; Tang, Z.; Yu, J.; Liao, W.; Xie, J.; Lv, J.; Feng, Z.; Dawuda, M.M. Hydrogen sulfide negatively regulates cd-induced cell death in cucumber (Cucumis sativus L.) root tip cells. BMC Plant Biol. 2020, 20, 480. [Google Scholar] [CrossRef] [PubMed]
- Zoschke, R.; Bock, R. Chloroplast translation: Structural and functional organization, operational control, and regulation. Plant Cell 2018, 30, 745–770. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Shen, W.; Gu, Y.; Hu, J.; Ma, Y.; Zhang, X.; Du, Y.; Zhang, Y.; Du, J. Exogenous melatonin mitigates saline-alkali stress by decreasing DNA oxidative damage and enhancing photosynthetic carbon metabolism in soybean (Glycine max [L.] Merr.) leaves. Physiol. Plant. 2023, 175, e13983. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Ouyang, Y.; Fan, Y.; Qiu, B.; Zhang, G.; Zeng, F. The pathway of transmembrane cadmium influx via calcium-permeable channels and its spatial characteristics along rice root. J. Exp. Bot. 2018, 69, 5279–5291. [Google Scholar] [CrossRef]
- Grajek, H.; Rydzyński, D.; Piotrowicz-Cieślak, A.; Herman, A.; Maciejczyk, M.; Wieczorek, Z. Cadmium ion-chlorophyll interaction—Examination of spectral properties and structure of the cadmium-chlorophyll complex and their relevance to photosynthesis inhibition. Chemosphere 2020, 261, 127434. [Google Scholar] [CrossRef]
- Barnett, J.Z. Effects of Light Quality and Light Quantity on the Growth Kinetics of a Louisiana Native Microalgal/Cyanobacterial Co-Culture. Master’s Thesis, Louisiana State University and Agricultural & Mechanical College, Baton Rouge, LA, USA, 2015. [Google Scholar]
- Behrendt, L.; Larkum, A.W.; Norman, A.; Qvortrup, K.; Chen, M.; Ralph, P.; Sørensen, S.J.; Trampe, E.; Kühl, M. Endolithic chlorophyll d-containing phototrophs. ISME J. 2011, 5, 1072–1076. [Google Scholar] [CrossRef]
- Kume, A.; Akitsu, T.; Nasahara, K.N. Why is chlorophyll b only used in light-harvesting systems? J. Plant Res. 2018, 131, 961–972. [Google Scholar] [CrossRef]
- Büchel, C. Light harvesting complexes in chlorophyll c-containing algae. Biochim. Biophys. Acta-Bioenerg. 2020, 1861, 148027. [Google Scholar] [CrossRef]
- Yamatani, H.; Ito, T.; Nishimura, K.; Yamada, T.; Sakamoto, W.; Kusaba, M. Genetic analysis of chlorophyll synthesis and degradation regulated by balance of chlorophyll metabolism. Plant Physiol. 2022, 189, 419–432. [Google Scholar] [CrossRef]
- Masuda, T.; Fujita, Y. Regulation and evolution of chlorophyll metabolism. Photochem. Photobiol. Sci. 2008, 7, 1131–1149. [Google Scholar] [CrossRef]
- Kobayashi, K.; Masuda, T. 97 Tetrapyrrole biosynthesis in plant systems. In Handbook of Porphyrin Science; World Scientific Publishing Company: Singapore, 2012; pp. 141–211. [Google Scholar]
- Li, X.; Zhang, W.; Niu, D.; Liu, X. Effects of abiotic stress on chlorophyll metabolism. Plant Sci. 2024, 342, 112030. [Google Scholar] [CrossRef] [PubMed]
- Müller, T.; Moser, S.; Ongania, K.H.; Pruzinska, A.; Hörtensteiner, S.; Kräutler, B. A divergent path of chlorophyll breakdown in the model plant Arabidopsis thaliana. ChemBioChem 2006, 7, 40–42. [Google Scholar] [CrossRef] [PubMed]
- Hörtensteiner, S. Chlorophyll degradation during senescence. Annu. Rev. Plant Biol. 2006, 57, 55–77. [Google Scholar] [CrossRef] [PubMed]
- Hörtensteiner, S.; Kräutler, B. Chlorophyll breakdown in higher plants. Biochim. Biophys. Acta 2011, 1807, 977–988. [Google Scholar] [CrossRef]
- Khan, K.Y.; Ali, B.; Stoffella, P.J.; Cui, X.; Yang, X.; Guo, Y. Study amino acid contents, plant growth variables and cell ultrastructural changes induced by cadmium stress between two contrasting cadmium accumulating cultivars of Brassica rapa ssp. chinensis L. (pak choi). Ecotoxicol. Environ. Saf. 2020, 200, 110748. [Google Scholar] [CrossRef]
- Li, X.; Wang, S.; Chen, X.; Cong, Y.; Cui, J.; Shi, Q.; Liu, H.; Diao, M. The positive effects of exogenous sodium nitroprusside on the plant growth, photosystem II efficiency and Calvin cycle of tomato seedlings under salt stress. Sci. Hortic. 2022, 299, 111016. [Google Scholar] [CrossRef]
- Rhimi, N.; Hajji, M.; Elkhouni, A.; Ksiaa, M.; Rabhi, M.; Lefi, E.; Smaoui, A.; Hessini, K.; Hamzaoui, A.H.; Cabassa-Hourton, C.; et al. Silicon reduces cadmium accumulation and improves growth and stomatal traits in sea barley (Hordeum marinum Huds.) exposed to cadmium stress. J. Soil Sci. Plant Nutr. 2024, 24, 2232–2248. [Google Scholar] [CrossRef]
- Chen, H.; Huang, X.; Chen, H.; Zhang, S.; Fan, C.; Fu, T.; He, T.; Gao, Z. Effect of silicon spraying on rice photosynthesis and antioxidant defense system on cadmium accumulation. Sci. Rep. 2024, 14, 15265. [Google Scholar] [CrossRef]
- Wu, J.; Qian, C.; Liu, Z.; Zhong, X. Effect of cadmium stress on the absorption and transportation of metal micronutrient elements in Hybrid Pennisetum. Pratac. Sci. 2023, 40, 133–143. (In Chinese) [Google Scholar] [CrossRef]
- Piao, L.; Wang, Y.; Liu, X.; Sun, G.; Zhang, S.; Yan, J.; Chen, Y.; Meng, Y.; Li, M.; Gu, W. Exogenous Hemin alleviated cadmium stress in maize (Zea mays L.) by enhancing leaf photosynthesis, AsA-GSH cycle and polyamine metabolism. Front. Plant Sci. 2022, 13, 993675. [Google Scholar] [CrossRef]
- Nolan, T.M.; Vukašinović, N.; Liu, D.; Russinova, E.; Yin, Y. Brassinosteroids: Multidimensional regulators of plant growth, development, and stress responses. Plant Cell 2020, 32, 295–318. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Wang, S.; You, X.; Wen, Z.; Huang, G.; Huang, C.; Li, Q.; Chen, K.; Zhao, Y.; Gu, M.; et al. Effect of foliar spraying of gibberellins and brassinolide on cadmium accumulation in rice. Toxics 2023, 11, 364. [Google Scholar] [CrossRef] [PubMed]
- Chmur, M.; Bajguz, A. Comparative efficacy of melatonin brassinolide in mitigating the adverse effects of cadmium on Wolffia arrhiza. Int. J. Mol. Sci. 2025, 26, 692. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Feng, B.; Zhou, Y.; Liu, C.; Gong, X. Exogenous brassinosteroids increases tolerance to shading by altering stress responses in mung bean (Vigna radiata L.). Photosynth. Res. 2022, 151, 279–294. [Google Scholar] [CrossRef]
- Tian, F.; Han, C.; Chen, X.; Wu, X.; Mi, J.; Wan, X.; Liu, Q.; He, F.; Chen, L.; Yang, H.; et al. PscCYP716A1-mediated brassinolide biosynthesis increases cadmium tolerance and enrichment in poplar. Front. Plant Sci. 2022, 13, 919682. [Google Scholar] [CrossRef]
- Zhang, S. Plant Physiology Experimental Techniques Tutorial; Science Press: Beijing, China, 2011. (In Chinese) [Google Scholar]
- Gao, J.F. Experimental Guidance for Plant Physiology; Higher Education Press: Beijing, China, 2006. (In Chinese) [Google Scholar]
- Li, H. Principles and Techniques of Plant Physiological Biochemical Experimental; Higher Education Press: Beijing, China, 2000. (In Chinese) [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using Real-time Quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Tang, Z.; Wang, H.Q.; Chen, J.; Chang, J.D.; Zhao, F.J. Molecular mechanisms underlying the toxicity and detoxification of trace metals and metalloids in plants. J. Integr. Plant Biol. 2023, 65, 570–593. [Google Scholar] [CrossRef]
- Kolahi, M.; Mohajel Kazemi, E.; Yazdi, M.; Goldson-Barnaby, A. Oxidative stress induced by cadmium in lettuce (Lactuca sativa Linn.): Oxidative stress indicators and prediction of their genes. Plant Physiol. Biochem. 2020, 146, 71–89. [Google Scholar] [CrossRef]
- Khursheed, M.; Shahbaz, M.; Ramzan, T.; Haider, A.; Maqsood, M.F.; Khan, A.; Zulfiqar, U.; Jamil, M.; Hussain, S.; Al-Ghamdi, A.A.; et al. Enhancing wheat tolerance to cadmium stress through moringa leaf extract foliar application. Scientifica 2024, 2024, 2919557. [Google Scholar] [CrossRef]
- Ding, M.; Wang, L.; Sun, Y.; Zhang, J.; Chen, Y.; Wang, X.; Liu, L. Transcriptome analysis of brassinolide under low temperature stress in winter wheat. AoB Plants 2023, 15, plad005. [Google Scholar] [CrossRef]
- Naservafaei, S.; Sohrabi, Y.; Moradi, P.; Mac Sweeney, E.; Mastinu, A. Biological response of Lallemantia iberica to brassinolide treatment under different watering conditions. Plants 2021, 10, 496. [Google Scholar] [CrossRef] [PubMed]
- Mu, D.W.; Feng, N.J.; Zheng, D.F.; Zhou, H.; Liu, L.; Chen, G.J.; Mu, B. Physiological mechanism of exogenous brassinolide alleviating salt stress injury in rice seedlings. Sci. Rep. 2022, 12, 20439. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.J.; Wang, Y.J.; Zhou, Y.H.; Tao, Y.; Mao, W.H.; Shi, K.; Asami, T.; Chen, Z.; Yu, J.Q. Reactive oxygen species are involved in brassinosteroid-induced stress tolerance in cucumber. Plant Physiol. 2009, 150, 801–814. [Google Scholar] [CrossRef] [PubMed]
- Waszczak, C.; Carmody, M.; Kangasjärvi, J. Reactive oxygen species in plant signaling. Annu. Rev. Plant Biol. 2018, 69, 209–236. [Google Scholar] [CrossRef]
- Asada, K. Production and scavenging of reactive oxygen species in chloroplasts and their functions. Plant Physiol. 2006, 141, 391–396. [Google Scholar] [CrossRef]
- Kerchev, P.I.; Van Breusegem, F. Improving oxidative stress resilience in plants. Plant J. 2022, 109, 359–372. [Google Scholar] [CrossRef]
- El-Mashad, A.A.; Mohamed, H.I. Brassinolide alleviates salt stress and increases antioxidant activity of cowpea plants (Vigna sinensis). Protoplasma 2012, 249, 625–635. [Google Scholar] [CrossRef]
- Song, Y.J.; Li, Y.; Leng, Y.; Li, S.W. 24-epibrassinolide improves differential cadmium tolerance of mung bean roots, stems, and leaves via amending antioxidative systems similar to that of abscisic acid. Environ. Sci. Pollut. Res. Int. 2021, 28, 52032–52045. [Google Scholar] [CrossRef]
- Lin, Y.; Fan, L.; Xia, X.; Wang, Z.; Yin, Y.; Cheng, Y.; Li, Z. Melatonin decreases resistance to postharvest green mold on citrus fruit by scavenging defense-related reactive oxygen species. Postharvest Biol. Technol. 2019, 153, 21–30. [Google Scholar] [CrossRef]
- Chen, L.; Shiotani, K.; Togashi, T.; Miki, D.; Aoyama, M.; Wong, H.L.; Kawasaki, T.; Shimamoto, K. Analysis of the Rac/Rop small GTPase family in rice: Expression, subcellular localization and role in disease resistance. Plant Cell Physiol. 2010, 51, 585–595. [Google Scholar] [CrossRef]
- Smokvarska, M.; Francis, C.; Platre, M.P.; Fiche, J.B.; Alcon, C.; Dumont, X.; Nacry, P.; Bayle, V.; Nollmann, M.; Maurel, C.; et al. A plasma membrane nanodomain ensures signal specificity during osmotic signaling in plants. Curr. Biol. 2020, 30, 4654–4664.e4654. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Xu, L.; Xuan, P.; Tian, Z.; Liu, R. Thiourea and arginine synergistically preserve redox homeostasis and ionic balance for alleviating salinity stress in wheat. Sci. Rep. 2024, 14, 21375. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.; Zhao, Y.; Wang, W.; He, Y. Elevated temperature altered photosynthetic products in wheat seedlings and organic compounds and biological activity in rhizopshere soil under cadmium stress. Sci. Rep. 2015, 5, 14426. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Lei, Z.; Ye, R.; Zhou, W.; Zhou, Y.; Zou, Z.; Li, J.; Yi, L.; Dai, Z. Effects of cadmium on physiochemistry and bioactive substances of muskmelon (Cucumis melo L.). Molecules 2022, 27, 2913. [Google Scholar] [CrossRef]
- Wang, S.; Fu, Y.; Zheng, S.; Xu, Y.; Sun, Y. Phytotoxicity and accumulation of copper-based nanoparticles in Brassica under cadmium stress. Nanomaterials 2022, 12, 1497. [Google Scholar] [CrossRef]
- Zhang, Z.; Chao, M.; Wang, S.; Bu, J.; Tang, J.; Li, F.; Wang, Q.; Zhang, B. Proteome quantification of cotton xylem sap suggests the mechanisms of potassium-deficiency-induced changes in plant resistance to environmental stresses. Sci. Rep. 2016, 6, 21060. [Google Scholar] [CrossRef]
- Jalili, S.; Ehsanpour, A.A.; Javadirad, S.M. The role of melatonin on caspase-3-like activity and expression of the genes involved in programmed cell death (PCD) induced by in vitro salt stress in alfalfa (Medicago sativa L.) roots. Bot. Stud. 2022, 63, 19. [Google Scholar] [CrossRef]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: Effects, mechanisms and management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef]
- Jahan, M.S.; Hasan, M.M.; Alotaibi, F.S.; Alabdallah, N.M.; Alharbi, B.M.; Ramadan, K.M.A.; Bendary, E.S.A.; Alshehri, D.; Jabborova, D.; Al-Balawi, D.A.; et al. Exogenous putrescine increases heat tolerance in tomato seedlings by regulating chlorophyll metabolism and enhancing antioxidant defense efficiency. Plants 2022, 11, 1038. [Google Scholar] [CrossRef]
- Zhu, Y.-F.; Wu, Y.-X.; Hu, Y.; Jia, X.-M.; Zhao, T.; Cheng, L.; Wang, Y.-X. Tolerance of two apple rootstocks to short-term salt stress: Focus on chlorophyll degradation, photosynthesis, hormone and leaf ultrastructures. Acta Physiol. Plant 2019, 41, 87. [Google Scholar] [CrossRef]
- Tang, Y.; Zhang, J.; Wang, L.; Wang, H.; Long, H.; Yang, L.; Li, G.; Guo, J.; Wang, Y.; Li, Y.; et al. Water deficit aggravated the inhibition of photosynthetic performance of maize under mercury stress but is alleviated by brassinosteroids. J. Hazard. Mater. 2023, 443, 130365. [Google Scholar] [CrossRef]
- Che, Y.; Fan, D.; Teng, Z.; Yao, T.; Wang, Z.; Zhang, H.; Sun, G.; Zhang, H.; Chow, W.S. Potassium alleviates over-reduction of the photosynthetic electron transport chain and helps to maintain photosynthetic function under salt-stress. Physiol. Plant. 2023, 175, e13981. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Plant Height (cm) | Leaf Area (cm2) | Leaf Dry Weight (g) | Above-Ground Dry Weight (g) | Growth Inhibition Ratio (%) | Cd Content (mg·g−1 DW) | ||
|---|---|---|---|---|---|---|---|---|
| Leaf | Stems | Petiole | ||||||
| W + Cd0 | 28.13 ± 1.10 ab | 521.16 ± 28.47 a | 1.41 ± 0.10 a | 2.22 ± 0.15 a | - | - | - | - |
| W + Cd1 | 24.80 ± 1.93 cd | 321.24 ± 15.94 c | 0.79 ± 0.02 c | 1.16 ± 0.02 c | 38.38 ± 0.66 b | 2.11 ± 0.33 b | 1.13 ± 0.07 b | 0.25 ± 0.01 ab |
| W + Cd2 | 22.43 ± 0.51 d | 276.50 ± 11.40 d | 0.77 ± 0.11 d | 1.16 ± 0.10 d | 47.62 ± 0.65 a | 3.98 ± 0.25 a | 1.68 ± 0.11 a | 0.26 ± 0.01 a |
| BR + Cd0 | 29.77 ± 2.64 ab | 506.69 ± 22.29 a | 1.43 ± 0.08 a | 2.21 ± 0.13 a | 0.28 ± 0.11 d | - | - | - |
| BR + Cd1 | 27.57 ± 0.83 abc | 435.56 ± 17.72 b | 1.15 ± 0.06 b | 1.75 ± 0.10 b | 16.51 ± 0.23 c | 1.68 ± 0.16 c | 0.89 ± 0.09 c | 0.21 ± 0.01 b |
| BR + Cd2 | 26.43 ± 0.81 bc | 433.35 ± 20.35 b | 1.16 ± 0.15 b | 1.80 ± 0.17 b | 16.89 ± 0.33 c | 2.55 ± 0.13 b | 1.13 ± 0.02 b | 0.22 ± 0.02 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, S.; Tang, Z.; Hou, J.; Gao, J.; Li, X.; Zhang, Y.; Zhao, Q. 2,4-Epibrassinolide Mitigates Cd Stress by Enhancing Chloroplast Structural Remodeling and Chlorophyll Metabolism in Vigna angularis Leaves. Biology 2025, 14, 674. https://doi.org/10.3390/biology14060674
Chen S, Tang Z, Hou J, Gao J, Li X, Zhang Y, Zhao Q. 2,4-Epibrassinolide Mitigates Cd Stress by Enhancing Chloroplast Structural Remodeling and Chlorophyll Metabolism in Vigna angularis Leaves. Biology. 2025; 14(6):674. https://doi.org/10.3390/biology14060674
Chicago/Turabian StyleChen, Suyu, Zihan Tang, Jialin Hou, Jie Gao, Xin Li, Yuxian Zhang, and Qiang Zhao. 2025. "2,4-Epibrassinolide Mitigates Cd Stress by Enhancing Chloroplast Structural Remodeling and Chlorophyll Metabolism in Vigna angularis Leaves" Biology 14, no. 6: 674. https://doi.org/10.3390/biology14060674
APA StyleChen, S., Tang, Z., Hou, J., Gao, J., Li, X., Zhang, Y., & Zhao, Q. (2025). 2,4-Epibrassinolide Mitigates Cd Stress by Enhancing Chloroplast Structural Remodeling and Chlorophyll Metabolism in Vigna angularis Leaves. Biology, 14(6), 674. https://doi.org/10.3390/biology14060674