
Genomic Evaluation of Harvest Weight Uniformity in Penaeus vannamei Under a 3FAM Design Incorporating Indirect Genetic Effect


Simple Summary
Abstract
1. Introduction
2. Materials and Methods
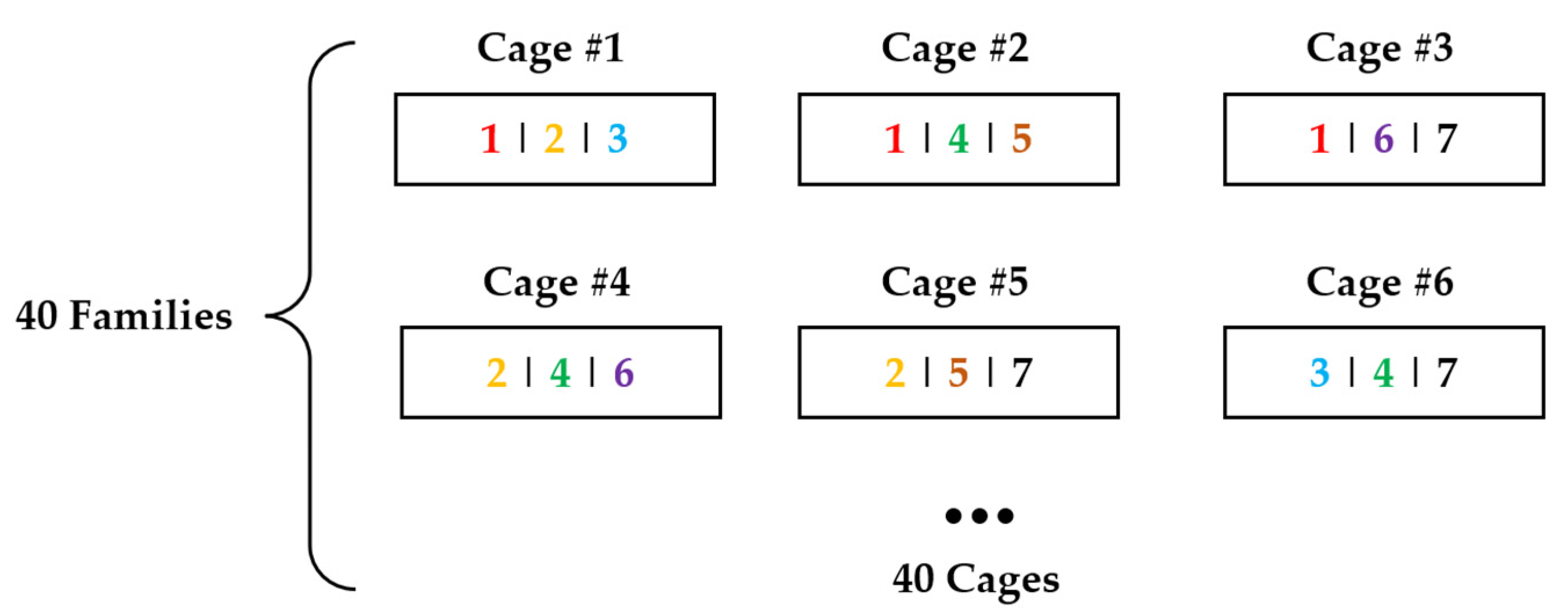
2.1. Samples and Experimental Design
2.2. Genotype and Quality Control
2.3. Statistical Analysis
2.4. Genetic Parameters
2.4.1. Estimation of Genetic Parameters for HW
2.4.2. Estimation of Genetic Parameters for HWU
2.5. Cross Validation
3. Results
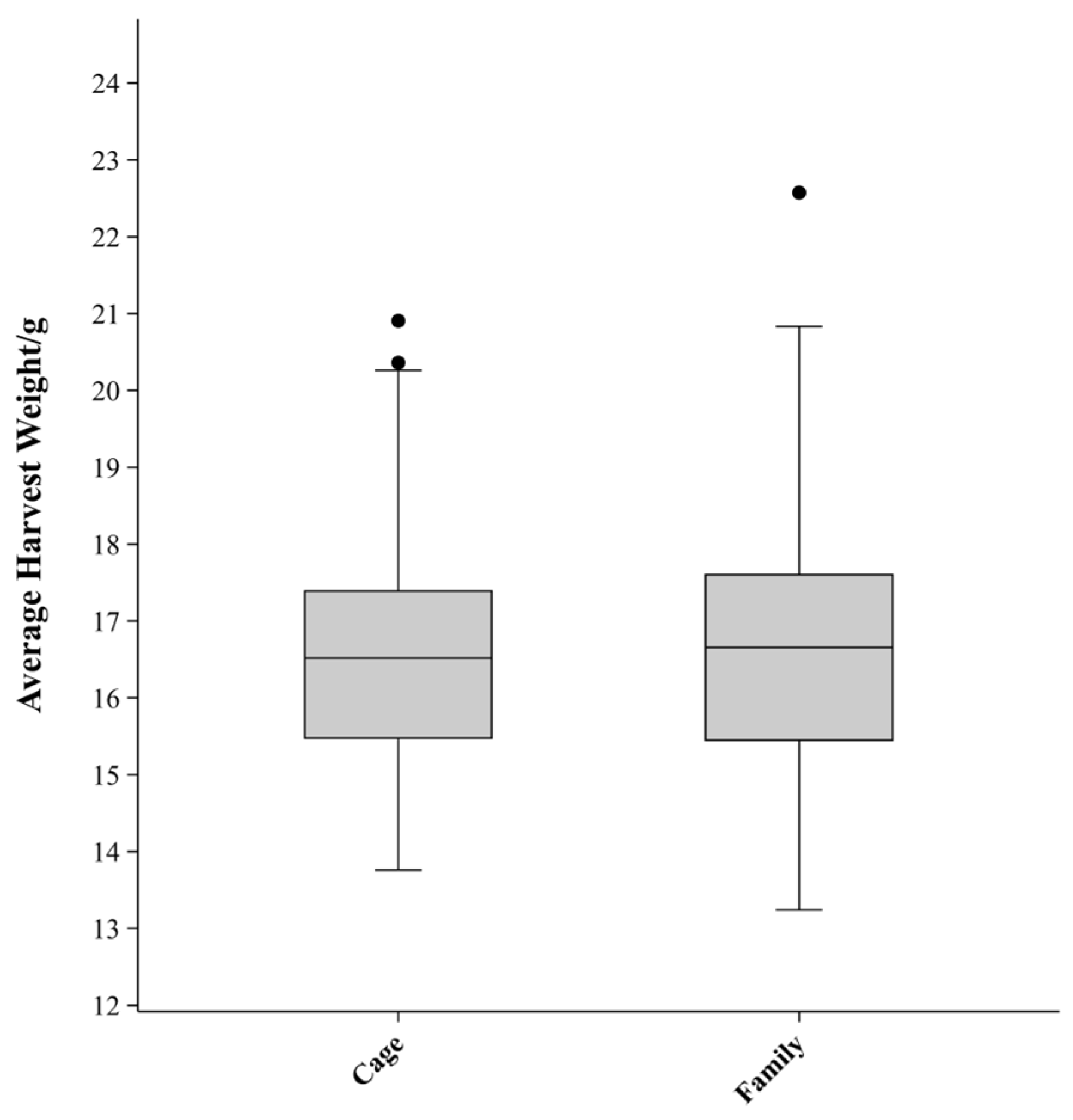
3.1. Descriptive Statistics
3.2. Genetic Parameters for HW and HWU
3.3. Prediction Accuracy
4. Discussion
4.1. Genetic Parameters of HWU
4.2. The Influence of IGEs
4.3. Genetic Correlation Between HW and HWU
4.4. The Prediction Accuracy
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2022: Towards Blue Transformation; FAO: Rome, Italy, 2022. [Google Scholar]
- Garcia, B.F.; Montaldo, H.H.; Iung, L.H.; Carvalheiro, R. Effect of harvest weight and its uniformity on survival in Litopenaeus vannamei reared in different systems. Aquaculture 2021, 531, 735891. [Google Scholar] [CrossRef]
- Kong, Z.; Kong, J.; Hao, D.; Lu, X.; Jian, T.; Meng, X.; Luo, K.; Cao, B.; Sui, J.; Li, X.; et al. Reducing the Common Environmental Effect on Litopenaeus vannamei Body Weight by Rearing Communally at Early Developmental Stages and Using a Reconstructed Pedigree. J. Ocean. Univ. China 2020, 19, 923–930. [Google Scholar] [CrossRef]
- Mulder, H.A.; Bijma, P.; Hill, W.G. Prediction of breeding values and selection responses with genetic heterogeneity of environmental variance. Genetics 2007, 175, 1895–1910. [Google Scholar] [CrossRef] [PubMed]
- Sorensen, D.; Waagepetersen, R. Normal linear models with genetically structured residual variance heterogeneity: A case study. Genet. Res. 2003, 82, 207–222. [Google Scholar] [CrossRef]
- García-Ballesteros, S.; Villanueva, B.; Fernández, J.; Gutiérrez, J.P.; Cervantes, I. Genetic parameters for uniformity of harvest weight in Pacific white shrimp (Litopenaeus vannamei). Genet. Sel. Evol. 2021, 53, 26. [Google Scholar] [CrossRef]
- Marjanovic, J.; Mulder, H.A.; Khaw, H.L.; Bijma, P. Genetic parameters for uniformity of harvest weight and body size traits in the GIFT strain of Nile tilapia. Genet. Sel. Evol. 2016, 48, 41. [Google Scholar] [CrossRef]
- Berghof, T.V.; Poppe, M.; Mulder, H.A. Opportunities to improve resilience in animal breeding programs. Front. Genet. 2019, 9, 692. [Google Scholar] [CrossRef]
- Sonesson, A.K.; Ødegård, J.; Rönnegård, L. Genetic heterogeneity of within-family variance of body weight in Atlantic salmon (Salmo salar). Genet. Sel. Evol. 2013, 45, 41. [Google Scholar] [CrossRef]
- Rönnegård, L.; Felleki, M.; Fikse, F.; Mulder, H.A.; Strandberg, E. Genetic heterogeneity of residual variance: Estimation of variance components using double hierarchical generalized linear models. Genet. Sel. Evol. 2010, 42, 8. [Google Scholar] [CrossRef]
- Felleki, M.; Lee, D.; Lee, Y.; Gilmour, A.R.; Rönnegård, L. Estimation of breeding values for mean and dispersion, their variance and correlation using double hierarchical generalized linear models. Genet. Res. 2012, 94, 307–317. [Google Scholar] [CrossRef]
- Hill, W.G.; Mulder, H.A. Genetic analysis of environmental variation. Genet. Res. 2010, 92, 381–395. [Google Scholar] [CrossRef]
- Sae-Lim, P.; Kause, A.; Janhunen, M.; Vehviläinen, H.; Koskinen, H.; Gjerde, B.; Lillehammer, M.; Mulder, H.A. Genetic (co) variance of rainbow trout (Oncorhynchus mykiss) body weight and its uniformity across production environments. Genet. Sel. Evol. 2015, 47, 46. [Google Scholar] [CrossRef]
- Sell-Kubiak, E.; Bijma, P.; Knol, E.F.; Mulder, H.A. Comparison of methods to study uniformity of traits: Application to birth weight in pigs. J. Anim. Sci. 2015, 93, 900–911. [Google Scholar] [CrossRef]
- Iung, L.H.D.S.; Mulder, H.A.; Neves, H.H.D.R.; Carvalheiro, R. Genomic regions underlying uniformity of yearling weight in Nellore cattle evaluated under different response variables. BMC Genom. 2018, 19, 619. [Google Scholar] [CrossRef]
- Sae-Lim, P.; Kause, A.; Lillehammer, M.; Mulder, H.A. Estimation of breeding values for uniformity of growth in Atlantic salmon (Salmo salar) using pedigree relationships or single-step genomic evaluation. Genet. Sel. Evol. 2017, 49, 33. [Google Scholar] [CrossRef]
- Hill, W.G.; Mackay, T.F. DS Falconer and Introduction to quantitative genetics. Genetics 2004, 167, 1529–1536. [Google Scholar] [CrossRef]
- Bijma, P. The quantitative genetics of indirect genetic effects: A selective review of modelling issues. Heredity 2014, 112, 61–69. [Google Scholar] [CrossRef]
- Hart, P.J.; Salvanes, A.G.V. Individual variation in competitive performance of juvenile cod and its consequences for growth. J. Mar. Biol. Assoc. UK 2000, 80, 569–570. [Google Scholar] [CrossRef]
- Cutts, C.J.; Metcalfe, N.B.; Taylor, A.C. Aggression and growth depression in juvenile Atlantic salmon: The consequences of individual variation in standard metabolic rate. J. Fish. Biol. 1998, 52, 1026–1037. [Google Scholar] [CrossRef]
- Wolf, J.B.; Brodie, E.D., III. The coadaptation of parental and offspring characters. Evolution 1998, 52, 299–308. [Google Scholar] [CrossRef]
- Guo, H.; Zhang, X.; Johnsson, J.I. Effects of size distribution on social interactions and growth of juvenile black rockfish (Sebastes schlegelii). Appl. Anim. Behav. Sci. 2017, 194, 135–142. [Google Scholar] [CrossRef]
- Khaw, H.L.; Ponzoni, R.W.; Yee, H.Y.; bin Aziz, M.A.; Bijma, P. Genetic and non-genetic indirect effects for harvest weight in the GIFT strain of Nile tilapia (Oreochromis niloticus). Aquaculture 2016, 450, 154–161. [Google Scholar] [CrossRef]
- Wang, N.; Hayward, R.S.; Noltie, D.B. Effects of social interaction on growth of juvenile hybrid sunfish held at two densities. N. Am. J. Aquac. 2000, 62, 161–167. [Google Scholar] [CrossRef]
- Luan, S.; Qiang, G.; Cao, B.; Luo, K.; Meng, X.; Chen, B.; Kong, J. Feed competition reduces heritable variation for body weight in Litopenaeus vannamei. Genet. Sel. Evol. 2020, 52, 45. [Google Scholar] [CrossRef]
- Moore, A.J.; Brodie, E.D., III; Wolf, J.B. Interacting phenotypes and the evolutionary process: I. Direct and indirect genetic effects of social interactions. Evolution 1997, 51, 1352–1362. [Google Scholar] [CrossRef]
- Sae-Lim, P.; Bijma, P. Comparison of designs for estimating genetic parameters and obtaining response to selection for social interaction traits in aquaculture. Aquaculture 2016, 451, 330–339. [Google Scholar] [CrossRef]
- Ødegård, J.; Olesen, I. Comparison of testing designs for genetic evaluation of social effects in aquaculture species. Aquaculture 2011, 317, 74–78. [Google Scholar] [CrossRef]
- Bijma, P. Estimating indirect genetic effects: Precision of estimates and optimum designs. Genetics 2010, 186, 1013–1028. [Google Scholar] [CrossRef]
- Legarra, A.; Aguilar, I.; Misztal, I. A relationship matrix including full pedigree and genomic information. J. Dairy Sci. 2009, 92, 4656–4663. [Google Scholar] [CrossRef]
- Kudinov, A.A.; Nousiainen, A.; Koskinen, H.; Kause, A. Single-step genomic prediction for body weight and maturity age in Finnish rainbow trout (Oncorhynchus mykiss). Aquaculture 2024, 585, 740677. [Google Scholar] [CrossRef]
- Yoshida, G.M.; Lhorente, J.P.; Correa, K.; Soto, J.; Salas, D.; Yáñez, J.M. Genome-wide association study and cost-efficient genomic predictions for growth and fillet yield in Nile tilapia (Oreochromis niloticus). G3-Genes Genom. Genet. 2019, 9, 2597–2607. [Google Scholar] [CrossRef] [PubMed]
- Garcia, A.L.; Bosworth, B.; Waldbieser, G.; Misztal, I.; Tsuruta, S.; Lourenco, D.A. Development of genomic predictions for harvest and carcass weight in channel catfish. Genet. Sel. Evol. 2018, 50, 66. [Google Scholar] [CrossRef]
- Liu, J.; Yang, G.; Kong, J.; Xia, Z.; Sui, J.; Tang, Q.; Luo, K.; Dai, P.; Lu, X.; Meng, X.; et al. Using single-step genomic best linear unbiased prediction to improve the efficiency of genetic evaluation on body weight in Macrobrachium rosenbergii. Aquaculture 2020, 528, 735577. [Google Scholar] [CrossRef]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.; Bender, D.; Maller, J.; Sklar, P.; De Bakker, P.I.; Daly, M.J.; et al. PLINK: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Gilmour, A.R.; Gogel, B.J.; Cullis, B.R.; Welham, S.J.; Thompson, R. ASReml User Guide Release 4.2 Functional Specification; VSN International Ltd.: Hemel Hempstead, UK, 2021. [Google Scholar]
- Bijma, P.; Muir, W.M.; Van Arendonk, J.A. Multilevel selection 1: Quantitative genetics of inheritance and response to selection. Genetics 2007, 175, 277–288. [Google Scholar] [CrossRef]
- Iung, L.H.D.S.; Carvalheiro, R.; Neves, H.H.D.R.; Mulder, H.A. Genetics and genomics of uniformity and resilience in livestock and aquaculture species: A review. J. Anim. Breed. Genet. 2020, 137, 263–280. [Google Scholar] [CrossRef] [PubMed]
- Elgersma, G.G.; de Jong, G.; van der Linde, R.; Mulder, H.A. Fluctuations in milk yield are heritable and can be used as a resilience indicator to breed healthy cows. J. Dairy Sci. 2018, 101, 1240–1250. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, H.M.; Monsen, B.B.; Ødegård, J.; Bijma, P.; Damsgård, B.; Toften, H.; Olesen, I. Direct and social genetic parameters for growth and fin damage traits in Atlantic cod (Gadus morhua). Genet. Sel. Evol. 2014, 46, 5. [Google Scholar] [CrossRef]
- Ellen, E.D.; Rodenburg, T.B.; Albers, G.A.; Bolhuis, J.E.; Camerlink, I.; Duijvesteijn, N.; Knol, E.F.; Muir, W.M.; Peeters, K.; Reimert, I.; et al. The prospects of selection for social genetic effects to improve welfare and productivity in livestock. Front. Genet. 2014, 5, 377. [Google Scholar] [CrossRef]
- Luan, S.; Luo, K.; Chai, Z.; Cao, B.; Meng, X.; Lu, X.; Liu, N.; Xu, S.; Kong, J. An analysis of indirect genetic effects on adult body weight of the Pacific white shrimp Litopenaeus vannamei at low rearing density. Genet. Sel. Evol. 2015, 47, 95. [Google Scholar] [CrossRef]
- Houle, D. Comparing evolvability and variability of quantitative traits. Genetics 1992, 130, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Dai, P.; Kong, J.; Meng, X.; Sui, J.; Luo, K.; Chen, B.; Fu, Q.; Cao, B.; Cao, J.; et al. Assessing accuracy of genomic breeding values of selection candidates under biosecurity restrictions by progeny testing in Chinese shrimp Fenneropenaeus chinensis. Aquaculture 2023, 566, 739181. [Google Scholar] [CrossRef]
- Jurcic, E.J.; Villalba, P.V.; Pathauer, P.S.; Palazzini, D.A.; Oberschelp, G.P.; Harrand, L.; Garcia, M.N.; Aguirre, N.C.; Acuña, C.V.; Martínez, M.C.; et al. Single-step genomic prediction of Eucalyptus dunnii using different identity-by-descent and identity-by-state relationship matrices. Heredity 2021, 127, 176–189. [Google Scholar] [CrossRef]
- Mulder, H.A.; Crump, R.E.; Calus, M.P.L.; Veerkamp, R.F. Unraveling the genetic architecture of environmental variance of somatic cell score using high-density single nucleotide polymorphism and cow data from experimental farms. J. Dairy Sci. 2013, 96, 7306–7317. [Google Scholar] [CrossRef]
- Sell-Kubiak, E.; Wang, S.; Knol, E.F.; Mulder, H.A. Genetic analysis of within-litter variation in piglets’ birth weight using genomic or pedigree relationship matrices. J. Anim. Sci. 2015, 93, 1471–1480. [Google Scholar] [CrossRef] [PubMed]
- Poulsen, B.G.; Ask, B.; Nielsen, H.M.; Ostersen, T.; Christensen, O.F. Prediction of genetic merit for growth rate in pigs using animal models with indirect genetic effects and genomic information. Genet. Sel. Evol. 2020, 52, 58. [Google Scholar] [CrossRef]
- Leite, N.G.; Knol, E.; Tsuruta, S.; Nuphaus, S.; Vogelzang, R.; Lourenco, D. Using social interaction models for genetic analysis of skin damage in gilts. Genet. Sel. Evol. 2023, 55, 52. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Variance Components | Formula | Explanation |
|---|---|---|
| ) | ||
| ) | A_NoIGE and H_NoIGE | |
| ) |
| Sex | N1 | N2 | Mean of M1BW/g | Mean/g | Minimum/g | Maximum/g | Standard Deviation | Coefficient of Variation/% | Survival Rate/% |
|---|---|---|---|---|---|---|---|---|---|
| All | 1440 | 1335 | 4.81 | 16.65 | 7.2 | 26.8 | 2.64 | 15.84 | 92.71 |
| Male | / | 665 | 4.77 | 16.19 | 7.2 | 26.1 | 2.47 | 15.27 | / |
| Female | / | 670 | 4.88 | 17.10 | 8.1 | 26.8 | 2.72 | 15.91 | / |
| Parameters | A Matrix | H Matrix | ||
|---|---|---|---|---|
| A_NoIGE | A_IGE | H_NoIGE | H_IGE | |
| 0.139 ± 0.033 | 0.119 ± 0.031 | 0.139 ± 0.034 | 0.125 ± 0.033 | |
| _ | 0.00233 ± 0.0012 | _ | ||
| _ | 0.000136 ± 0.000117 | _ | 0.000187 ± 0.000088 | |
| 0.009 | 0.022 | 0.008 | 0.019 | |
| 0.109 | 0.246 | 0.137 | 0.228 | |
| 0.520 | 0.517 | 0.520 | 0.521 | |
| 0.229 | 0.246 | 0.226 | 0.242 | |
| 0.029 ± 0.012 | 0.008 ± 0.014 | 0.028 ± 0.011 | - | |
| 0.826 | 0.768 | 0.826 | 0.782 | |
| _ | 0.971 | _ | 1.072 | |
| 0.672 | 0.622 | 0.673 | 0.643 | |
| _ | 1.264 | _ | 1.380 | |
| 0.005 | 0.017 | 0.006 | 0.015 | |
| 0.340 | 0.528 | 0.383 | 0.506 | |
| _ | 0.578 ± 0.328 | _ | 0.509 ± 0.264 | |
| −0.862 ± 0.627 | 0.203 ± 0.213 | −0.683 ± 0.427 | 0.117 ± 0.230 | |
| Harvest Weight | Uniformity of Harvest Weight | |||
|---|---|---|---|---|
| Accuracy | Bias | Accuracy | Bias | |
| pBLUP | 0.362 | 1.159 | 0.076 | 1.253 |
| ssGBLUP | 0.385 | 1.212 | 0.084 | 1.425 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, S.; Xia, Y.; Kong, J.; Meng, X.; Luo, K.; Sui, J.; Dai, P.; Tan, J.; Li, X.; Cao, J.; et al. Genomic Evaluation of Harvest Weight Uniformity in Penaeus vannamei Under a 3FAM Design Incorporating Indirect Genetic Effect. Biology 2025, 14, 328. https://doi.org/10.3390/biology14040328
Gao S, Xia Y, Kong J, Meng X, Luo K, Sui J, Dai P, Tan J, Li X, Cao J, et al. Genomic Evaluation of Harvest Weight Uniformity in Penaeus vannamei Under a 3FAM Design Incorporating Indirect Genetic Effect. Biology. 2025; 14(4):328. https://doi.org/10.3390/biology14040328
Chicago/Turabian StyleGao, Siqi, Yan Xia, Jie Kong, Xianhong Meng, Kun Luo, Juan Sui, Ping Dai, Jian Tan, Xupeng Li, Jiawang Cao, and et al. 2025. "Genomic Evaluation of Harvest Weight Uniformity in Penaeus vannamei Under a 3FAM Design Incorporating Indirect Genetic Effect" Biology 14, no. 4: 328. https://doi.org/10.3390/biology14040328
APA StyleGao, S., Xia, Y., Kong, J., Meng, X., Luo, K., Sui, J., Dai, P., Tan, J., Li, X., Cao, J., Chen, B., Fu, Q., Xing, Q., Tian, Y., Liu, J., & Luan, S. (2025). Genomic Evaluation of Harvest Weight Uniformity in Penaeus vannamei Under a 3FAM Design Incorporating Indirect Genetic Effect. Biology, 14(4), 328. https://doi.org/10.3390/biology14040328