

Unraveling the Prognostic Significance of BRCA1-Associated Protein 1 (BAP1) Expression in Advanced and Castrate-Resistant Prostate Cancer
 , , ,
, , , 
Simple Summary
Abstract
1. Introduction
2. Methods
2.1. Study Population, Tissue Microarray Construction, and Pathological Analysis
2.2. Immunohistochemistry
2.3. BAP1 Gene Expression Analysis in the Public Cancer Database
2.4. Statistical Analysis
3. Results
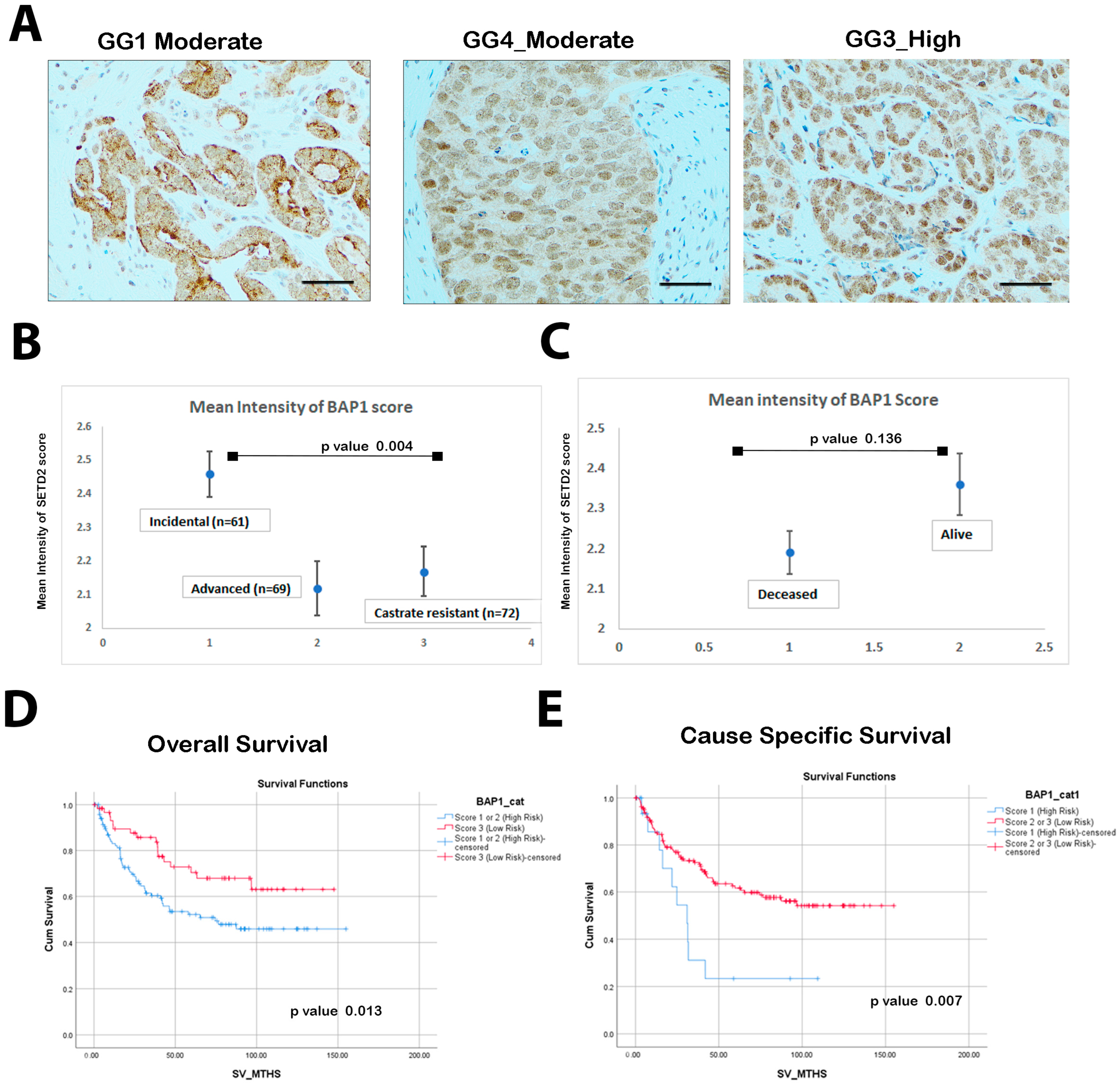
3.1. Low BAP1 Expression Correlates with Poor Overall Survival (OS) and Cancer-Specific Survival (CSS)
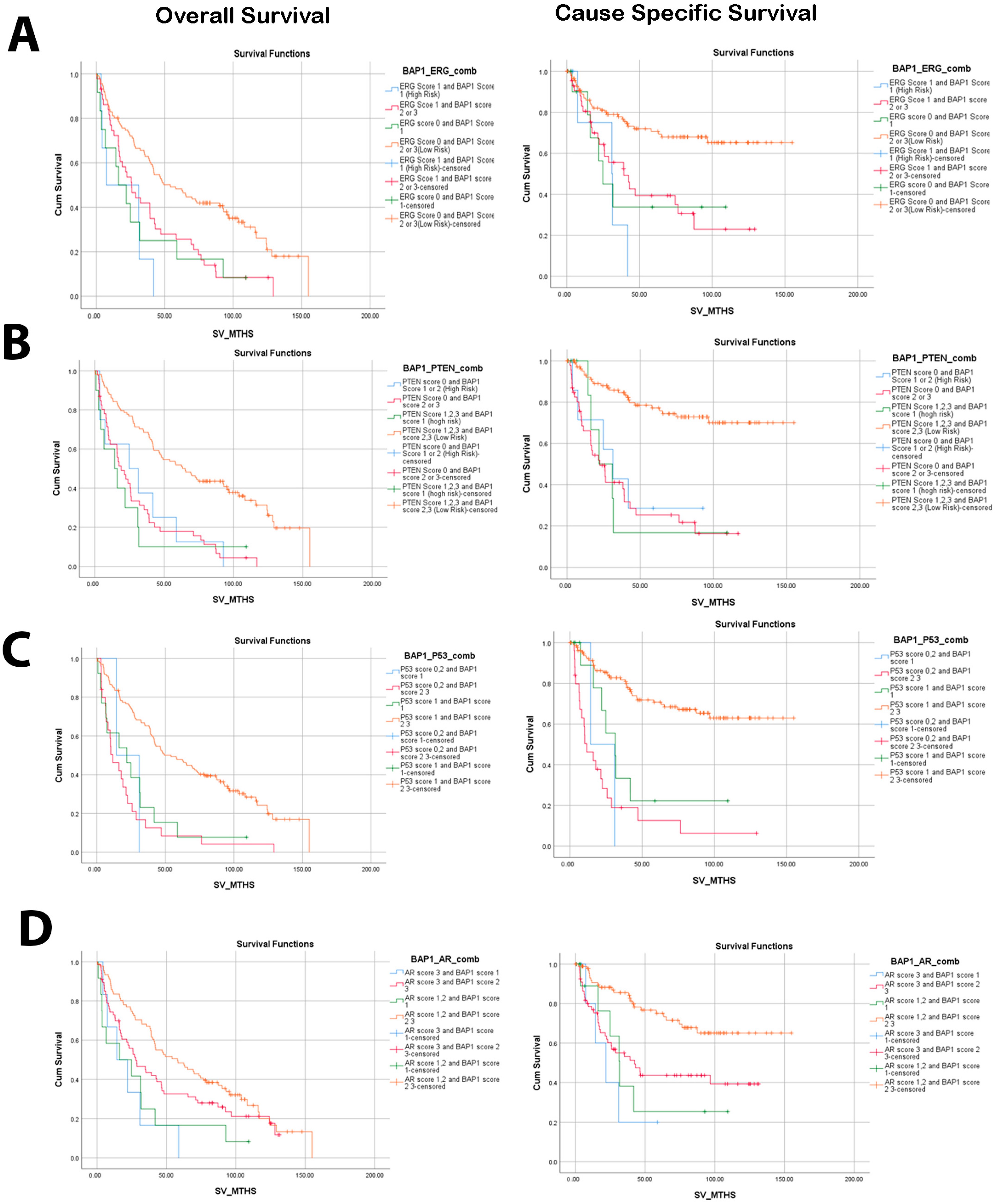
3.2. Low Nuclear BAP1 Expression Combined with Either ERG Expression, PTEN Loss, P53, or AR Mutant Associated with Poor OS and CSS
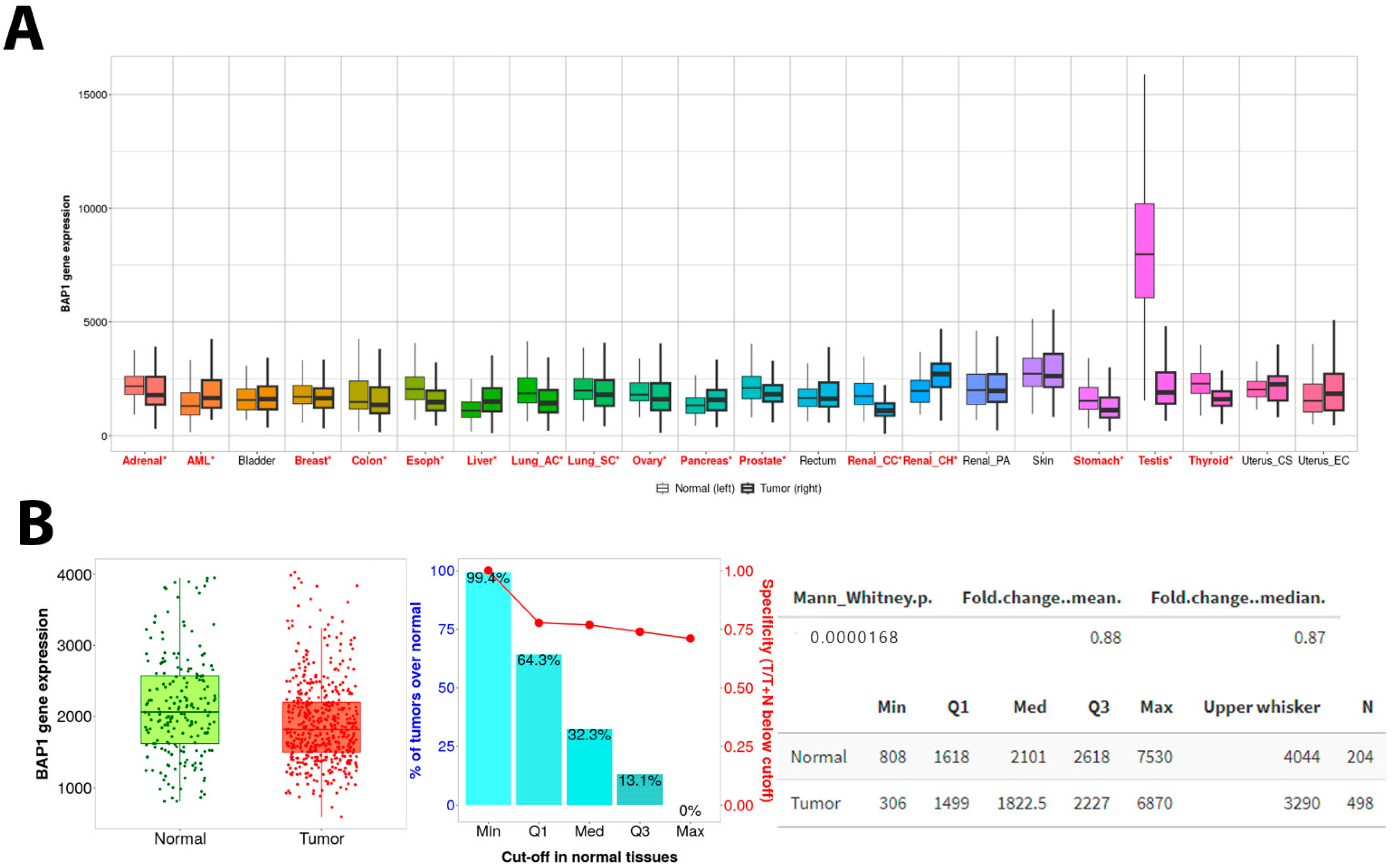
3.3. PanCancer Data Analysis Revealed BAP1 Expression in Different Cancers
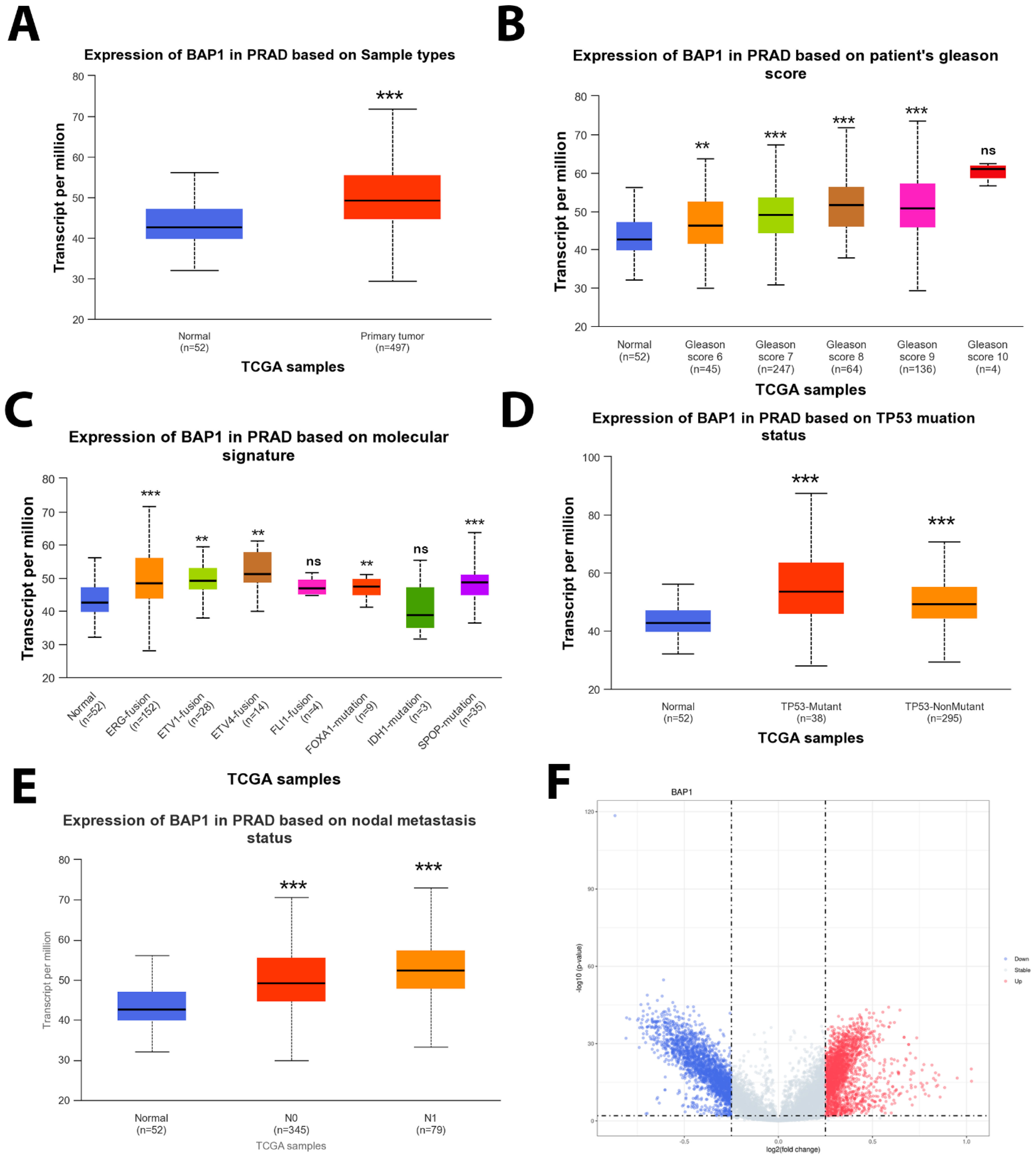
3.4. BAP1 Gene Expression in TCGA PRAD
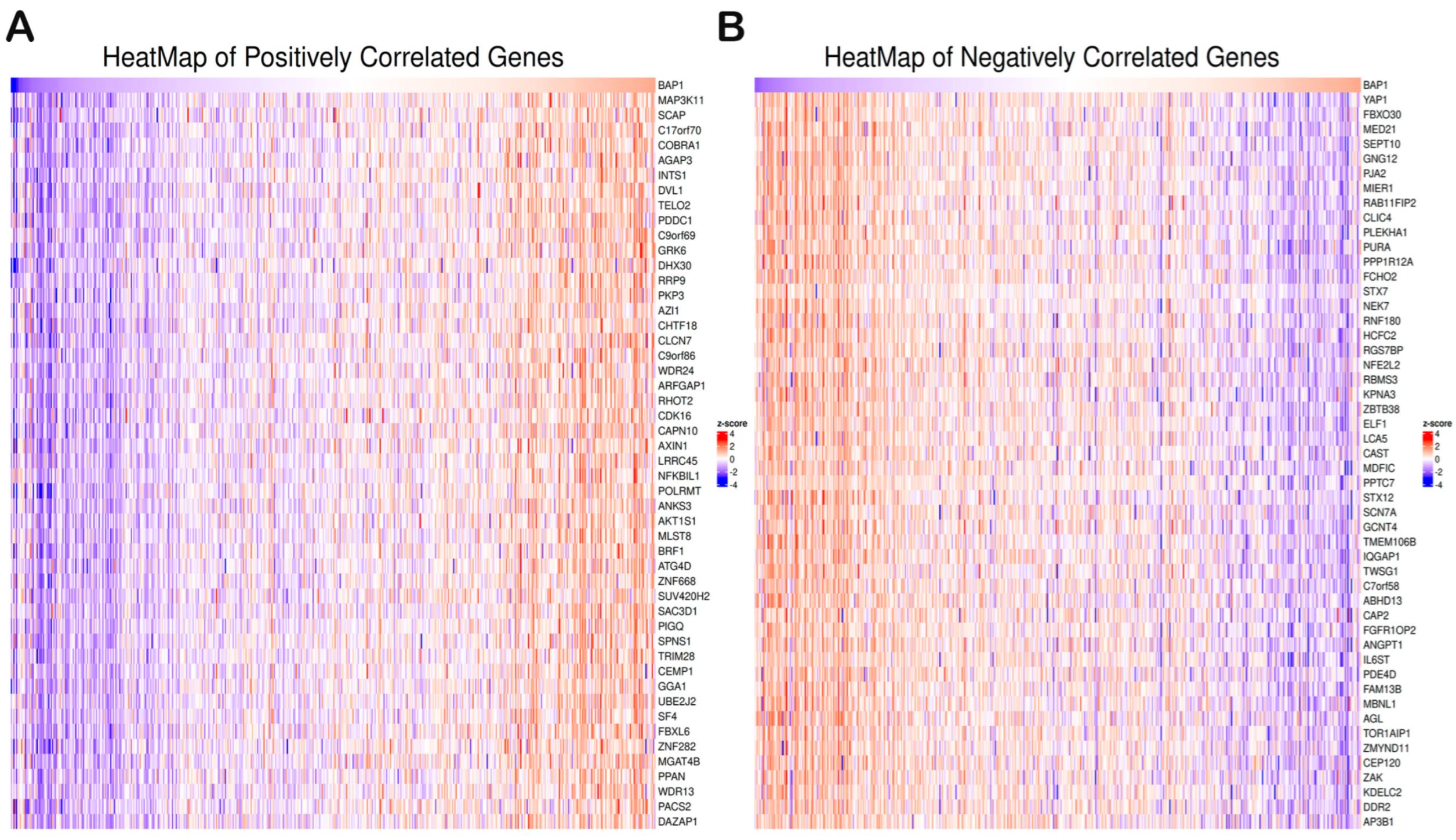
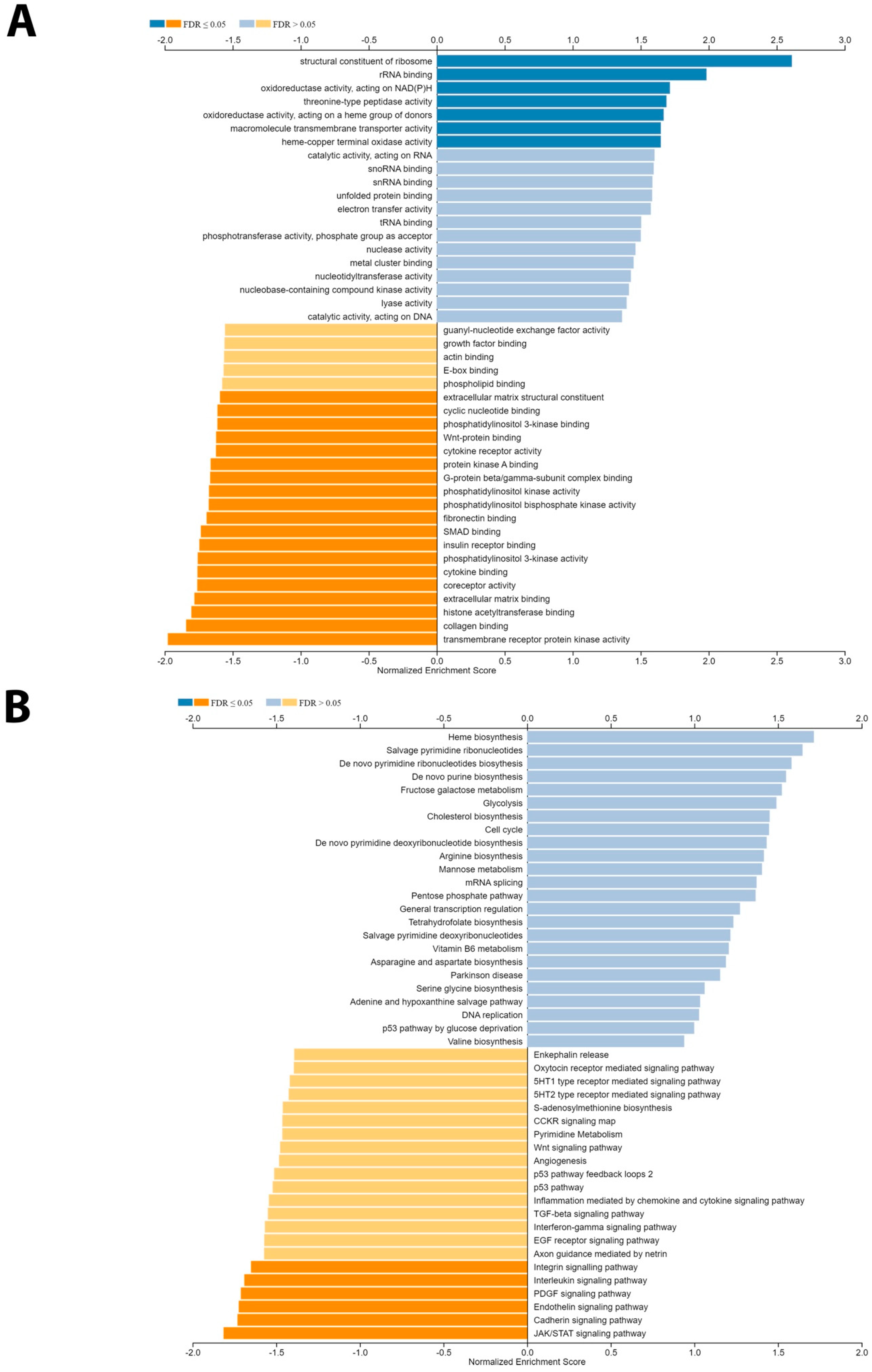
3.5. Gene Set Enrichment Analysis of BAP1 Gene Revealed Potential Role in PCa
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| IHC | Immunohistochemistry |
| TMA | Tissue microarray |
| GSEA | Gene set enrichment analysis |
| TCGA | The Cancer Genome Atlas |
| PRAD | Prostate Adenocarcinoma |
| CRPCa | Castration-resistant prostate cancer |
| PTEN | Phosphatase and tensin homolog |
| ERG | ETS Transcription Factor ERG |
| CSS | Cause-specific survival |
| OS | Overall survival |
References
- Berenguer, C.V.; Pereira, F.; Camara, J.S.; Pereira, J.A.M. Underlying Features of Prostate Cancer-Statistics, Risk Factors, and Emerging Methods for Its Diagnosis. Curr. Oncol. 2023, 30, 2300–2321. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Wagle, N.S.; Jemal, A. Cancer statistics, 2023. Ca Cancer J. Clin. 2023, 73, 17–48. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.T.; Barocas, D.; Carlsson, S.; Coakley, F.; Eggener, S.; Etzioni, R.; Fine, S.W.; Han, M.; Kim, S.K.; Kirkby, E. Early detection of prostate cancer: AUA/SUO guideline part I: Prostate cancer screening. J. Urol. 2023, 210, 46–53. [Google Scholar] [CrossRef]
- Sekhoacha, M.; Riet, K.; Motloung, P.; Gumenku, L.; Adegoke, A.; Mashele, S. Prostate cancer review: Genetics, diagnosis, treatment options, and alternative approaches. Molecules 2022, 27, 5730. [Google Scholar] [CrossRef]
- Diamantopoulos, M.A.; Georgoulia, K.K.; Levis, P.; Kotronopoulos, G.; Stravodimos, K.; Kontos, C.K.; Avgeris, M.; Scorilas, A. 28S rRNA-Derived Fragments Represent an Independent Molecular Predictor of Short-Term Relapse in Prostate Cancer. Int. J. Mol. Sci. 2023, 25, 239. [Google Scholar] [CrossRef]
- Bolzati, C.; Gobbi, C.; Ferro-Flores, G.; Turato, S.; Ocampo-Garcia, B.; Carpanese, D.; Marzano, C.; Spolaore, B.; Fracasso, G.; Rosato, A. Development and Characterization of 99mTc-scFvD2B as a Potential Radiopharmaceutical for SPECT Imaging of Prostate Cancer. Int. J. Mol. Sci. 2023, 25, 492. [Google Scholar] [CrossRef]
- Leslie, S.W.; Soon-Sutton, T.L.; Skelton, W.P. Prostate Cancer. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Valle, L.F.; Nickols, N.G.; Hausler, R.; Alba, P.R.; Anglin-Foote, T.; Perez, C.; Yamoah, K.; Rose, B.S.; Kelley, M.J.; DuVall, S.L. Actionable Genomic Alterations in Prostate Cancer Among Black and White United States Veterans. Oncologist 2023, 28, e473–e477. [Google Scholar] [CrossRef] [PubMed]
- Stopsack, K.H.; Su, X.A.; Vaselkiv, J.B.; Graff, R.E.; Ebot, E.M.; Pettersson, A.; Lis, R.T.; Fiorentino, M.; Loda, M.; Penney, K.L. Transcriptomes of prostate cancer with TMPRSS2: ERG and other ETS fusions. Mol. Cancer Res. 2023, 21, 14–23. [Google Scholar] [CrossRef]
- Arigbede, O.; Buxbaum, S.G.; Falzarano, S.; Rhie, S.K. Genomic landscape of prostate cancer: Bioinformatic analysis of TCGA PanCancer data by race and identification of key pathways and genes in PTEN mutation. Cancer Res. 2023, 83, 4266. [Google Scholar] [CrossRef]
- Kwan, E.M.; Wyatt, A.W. Androgen receptor genomic alterations and treatment resistance in metastatic prostate cancer. Prostate 2022, 82, S25–S36. [Google Scholar]
- Ding, D.; Blee, A.M.; Zhang, J.; Pan, Y.; Becker, N.A.; Maher, L.J., 3rd; Jimenez, R.; Wang, L.; Huang, H. Gain-of-function mutant p53 together with ERG proto-oncogene drive prostate cancer by beta-catenin activation and pyrimidine synthesis. Nat. Commun. 2023, 14, 4671. [Google Scholar] [CrossRef]
- Khan, S.; Baligar, P.; Tandon, C.; Nayyar, J.; Tandon, S. Molecular heterogeneity in prostate cancer and the role of targeted therapy. Life Sci. 2023, 336, 122270. [Google Scholar] [CrossRef] [PubMed]
- Deng, R.; Guo, Y.; Li, L.; He, J.; Qiang, Z.; Zhang, H.; Chen, R.; Wang, Y.; Zhao, X.; Yu, J. BAP1 suppresses prostate cancer progression by deubiquitinating and stabilizing PTEN. Mol. Oncol. 2021, 15, 279–298. [Google Scholar] [CrossRef] [PubMed]
- Park, C.M.; Lee, J.E.; Kim, J.H. BAP1 functions as a tumor promoter in prostate cancer cells through EMT regulation. Genet. Mol. Biol. 2020, 43, e20190328. [Google Scholar] [CrossRef]
- Carbone, M.; Harbour, J.W.; Brugarolas, J.; Bononi, A.; Pagano, I.; Dey, A.; Krausz, T.; Pass, H.I.; Yang, H.; Gaudino, G. Biological Mechanisms and Clinical Significance of BAP1 Mutations in Human Cancer. Cancer Discov. 2020, 10, 1103–1120. [Google Scholar] [CrossRef] [PubMed]
- Steurer, S.; Schwemmer, L.; Hube-Magg, C.; Buscheck, F.; Hoflmayer, D.; Tsourlakis, M.C.; Clauditz, T.S.; Luebke, A.M.; Simon, R.; Sauter, G.; et al. Nuclear up regulation of the BRCA1-associated ubiquitinase BAP1 is associated with tumor aggressiveness in prostate cancers lacking the TMPRSS2:ERG fusion. Oncotarget 2019, 10, 7096–7111. [Google Scholar] [CrossRef]
- Oh, H.; Lee, Y.J.; Kang, S.G.; Ahn, B.; Kim, E.; Chae, Y.S.; Lee, Y.; Lee, J.H.; Kim, C.H. BRCA1-associated protein 1 expression and prognostic role in prostate adenocarcinoma. Investig. Clin. Urol. 2020, 61, 166–172. [Google Scholar] [CrossRef]
- Bartha, A.; Gyorffy, B. TNMplot.com: A Web Tool for the Comparison of Gene Expression in Normal, Tumor and Metastatic Tissues. Int. J. Mol. Sci. 2021, 22, 2622. [Google Scholar] [CrossRef]
- Chandrashekar, D.S.; Karthikeyan, S.K.; Korla, P.K.; Patel, H.; Shovon, A.R.; Athar, M.; Netto, G.J.; Qin, Z.S.; Kumar, S.; Manne, U.; et al. UALCAN: An update to the integrated cancer data analysis platform. Neoplasia 2022, 25, 18–27. [Google Scholar] [CrossRef]
- Li, Q.; Gamallat, Y.; Rokne, J.G.; Bismar, T.A.; Alhajj, R. BioLake: An RNA expression analysis framework for prostate cancer biomarker powered by data lakehouse. BMC Bioinform. 2025, 26, 37. [Google Scholar]
- Vasaikar, S.V.; Straub, P.; Wang, J.; Zhang, B. LinkedOmics: Analyzing multi-omics data within and across 32 cancer types. Nucleic Acids Res. 2018, 46, D956–D963. [Google Scholar] [CrossRef]
- Vicentini, C.; Cantù, C.; Antonello, D.; Simbolo, M.; Mafficini, A.; Luchini, C.; Rusev, B.; Porcaro, A.B.; Iacovelli, R.; Fassan, M. ERG alterations and mTOR pathway activation in primary prostate carcinomas developing castration-resistance. Pathol.-Res. Pract. 2018, 214, 1675–1680. [Google Scholar] [CrossRef] [PubMed]
- Walker, S.R.; Abdelsalam, R.; Ghosh, S.; Livingstone, J.; Palanisamy, N.; Boutros, P.C.; Yip, S.M.; Lees-Miller, S.P.; Bismar, T.A. Decreased ATM Protein Expression Is Substantiated with PTEN Loss in Defining Aggressive Phenotype of Prostate Cancer Associated with Lethal Disease. Eur. Urol. Open Sci. 2021, 29, 93–101. [Google Scholar] [CrossRef]
- Helgadottir, H.; Schultz, K.; Lapins, J.; Vassilaki, I.; All-Eriksson, C.; Hoiom, V. BAP1 genetic testing among melanoma and cancer-prone families in Sweden. Acta Oncol. 2023, 62, 565–570. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.; Park, S.G.; Lee, S.A.; Kim, S.; Lee, D.; Shirbhate, M.E.; Youn, S.Y.; Kim, K.M.; Cha, S.S.; Kwon, J. Targeting BAP1 with small compound inhibitor for colon cancer treatment. Sci. Rep. 2023, 13, 2264. [Google Scholar] [CrossRef]
- Caporali, S.; Butera, A.; Amelio, I. BAP1 in cancer: Epigenetic stability and genome integrity. Discov. Oncol. 2022, 13, 117. [Google Scholar] [CrossRef] [PubMed]
- Kwon, J.; Lee, D.; Lee, S.A. BAP1 as a guardian of genome stability: Implications in human cancer. Exp. Mol. Med. 2023, 55, 745–754. [Google Scholar] [CrossRef]
- Lu, W.T.; Li, M.R.; Yang, Y.B.; Sun, Y.Y.; Dong, C.M. Research Progress of BAP1 in Structure, Function, and Cancer. Protein Pept. Lett. 2023, 30, 552–561. [Google Scholar] [CrossRef]
- Voulgari, O.; Goutas, D.; Pergaris, A.; Belogiannis, K.; Thymara, E.; Kavantzas, N.; Lazaris, A.C. Correlations of PTEN and ERG Immunoexpression in Prostate Carcinoma and Lesions Related to Its Natural History: Clinical Perspectives. Curr. Issues Mol. Biol. 2023, 45, 2767–2780. [Google Scholar] [CrossRef]
- Bhat, G.; Ahamed, M.A.; Saroha, N.; Prasad, G. Clinicopathological correlation of p53 expression in benign prostatic hyperplasia and prostate adenocarcinoma. Indian J. Pathol. Oncol. 2023, 9, 60–64. [Google Scholar]
- Fujita, K.; Nonomura, N. Role of androgen receptor in prostate cancer: A review. World J. Men’s Health 2019, 37, 288–295. [Google Scholar] [CrossRef] [PubMed]
- Couñago, F.; López-Campos, F.; Díaz-Gavela, A.A.; Almagro, E.; Fenández-Pascual, E.; Henríquez, I.; Lozano, R.; Linares Espinós, E.; Gómez-Iturriaga, A.; de Velasco, G. Clinical applications of molecular biomarkers in prostate cancer. Cancers 2020, 12, 1550. [Google Scholar] [CrossRef] [PubMed]
- Egevad, L.; Delahunt, B.; Kristiansen, G.; Samaratunga, H.; Varma, M. Contemporary prognostic indicators for prostate cancer incorporating International Society of Urological Pathology recommendations. Pathology 2018, 50, 60–73. [Google Scholar] [CrossRef] [PubMed]
- Koussounadis, A.; Langdon, S.P.; Um, I.H.; Harrison, D.J.; Smith, V.A. Relationship between differentially expressed mRNA and mRNA-protein correlations in a xenograft model system. Sci. Rep. 2015, 5, 10775. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Score 1 (n = 22) | Score 2 (n = 110) | Score 3 (n = 70) |
|---|---|---|---|
| Survival | |||
| Deceased | 17 (85.0) | 78 (71.6) | 44 (64.7) |
| Alive | 13 (46.4) | 31 (32.6) | 13 (18.3) |
| Cancer subgroup | |||
| Incidental | 1 (4.5) | 31 (28.2) | 29 (41.4) |
| Advanced | 12 (54.5) | 37 (33.6) | 20 (28.6) |
| Castrate-resistant | 9 (40.9) | 42 (38.2) | 21 (30.0) |
| PTEN | |||
| Loss | 9 (45.0) | 30 (29.4) | 16 (24.6) |
| Gain | 11 (55.0) | 72 (70.6) | 49 (75.4) |
| ERG | |||
| Negative | 13 (65.0) | 74 (71.80 | 50 (76.9) |
| Positive | 7 (35.0) | 29 (28.2) | 15 (23.1) |
| AR | |||
| Score 1, 2 | 14 (63.6) | 54 (50.5) | 50 (74.6) |
| Score 3 | 8 (36.4) | 53 (49.5) | 17 (25.4) |
| P53 | |||
| Score 1 | 14 (73.7) | 88 (82.2) | 60 (89.6) |
| Score 0, 2 | 5 (26.3) | 19 (17.8) | 7 (10.4) |
| Variables | Overall Survival HR (95% CI) | p-Value | Cause-Specific Survival HR (95% CI) | p-Value |
|---|---|---|---|---|
| PTEN (score 1, 2 or 3) | ||||
| Loss—score 0 | 2.71 (1.89–3.89) | <0.0001 | 4.50 (2.76–7.34) | <0.0001 |
| ERG (negative) | ||||
| Positive | 1.93 (1.34–2.77) | <0.0001 | 2.53 (1.55–4.11) | <0.0001 |
| GS (≤6) | ||||
| GS 3 + 4 | 2.03 (0.98–4.20) | 0.056 | 24.18 (2.70–216.62) | <0.0001 |
| GS 4 + 3 | 1.41 (0.72–2.77) | 0.322 | 9.68 (1.01–93.15) | 0.049 |
| GS 8 | 5.09 (2.47–10.49) | <0.0001 | 71.43 (8.68–588.10) | <0.0001 |
| GS 9, 10 | 6.02 (3.80–9.51) | <0.0001 | 107.23 (14.73–780.83) | <0.0001 |
| BAP1 low-risk (score 2, 3) | ||||
| BAP1 high risk (score 1) | 2.31 (1.38–3.86) | 0.001 | 2.44 (1.24–4.78) | 0.01 |
| Combination PTEN and BAP1 (PTEN score 1, 2, 3 and BAP1 score 2, 3) | ||||
| PTEN score 0 and BAP1 score 1 | 2.83 (1.35–5.91) | 0.006 | 4.30 (1.64–11.28) | 0.003 |
| PTEN score 0 and BAP1 score 2, 3 | 3.03 (2.05–4.46) | <0.0001 | 5.43 (3.19–9.24) | <0.0001 |
| PTEN score 1–3 and BAP1 score 1 | 3.57 (1.77–7.20) | <0.0001 | 4.95 (1.88–13.03) | 0.001 |
| Combination PTEN and BAP1 (PTEN score 1, 2, 3 and BAP1 score 2, 3) * | ||||
| PTEN score 0 and BAP1 score 1 | 2.34 (1.07–5.12) | 0.033 | 2.99 (1.06–8.43) | 0.038 |
| PTEN score 0 and BAP1 score 2, 3 | 1.63 (1.06–2.53) | 0.027 | 1.91 (1.10–3.33) | 0.023 |
| PTEN score 1–3 and BAP score 1 | 1.36 (0.65–2.86) | 0.421 | 1.13 (0.42–3.03) | 0.808 |
| Combination ERG and BAP1 (ERG negative and BAP1 score 0–2) | ||||
| ERG positive and BAP1 score 1 | 3.79 (1.62–6.82) | 0.002 | 4.72 (1.65–13.52) | 0.004 |
| ERG positive and BAP1 score 2, 3 | 1.97 (1.34–2.90) | 0.001 | 2.66 (1.57–4.50) | <0.0001 |
| ERG negative and BAP1 score 1 | 2.43 (1.29–4.60) | 0.006 | 2.83 (1.18–6.78) | 0.02- |
| Combination ERG and BAP1 (ERG negative and BAP1 score 0–2) * | ||||
| ERG positive and BAP1 score 1 | 1.65 (0.69–3.95) | 0.257 | 1.41 (0.49–4.06) | 0.53 |
| ERG positive and BAP1 score 2, 3 | 1.20 (0.79–1.84) | 0.395 | 1.12 (0.65–1.93) | 0.69 |
| ERG negative and BAP1 score 1 | 1.37 (0.71–2.64) | 0.347 | 1.12 (0.46–2.72) | 0.799 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salut, N.; Gamallat, Y.; Seyedi, S.; Felipe Lima, J.; Ghosh, S.; Bismar, T.A. Unraveling the Prognostic Significance of BRCA1-Associated Protein 1 (BAP1) Expression in Advanced and Castrate-Resistant Prostate Cancer. Biology 2025, 14, 315. https://doi.org/10.3390/biology14030315
Salut N, Gamallat Y, Seyedi S, Felipe Lima J, Ghosh S, Bismar TA. Unraveling the Prognostic Significance of BRCA1-Associated Protein 1 (BAP1) Expression in Advanced and Castrate-Resistant Prostate Cancer. Biology. 2025; 14(3):315. https://doi.org/10.3390/biology14030315
Chicago/Turabian StyleSalut, Norel, Yaser Gamallat, Sima Seyedi, Joema Felipe Lima, Sunita Ghosh, and Tarek A. Bismar. 2025. "Unraveling the Prognostic Significance of BRCA1-Associated Protein 1 (BAP1) Expression in Advanced and Castrate-Resistant Prostate Cancer" Biology 14, no. 3: 315. https://doi.org/10.3390/biology14030315
APA StyleSalut, N., Gamallat, Y., Seyedi, S., Felipe Lima, J., Ghosh, S., & Bismar, T. A. (2025). Unraveling the Prognostic Significance of BRCA1-Associated Protein 1 (BAP1) Expression in Advanced and Castrate-Resistant Prostate Cancer. Biology, 14(3), 315. https://doi.org/10.3390/biology14030315