Predicting the Potential Suitable Distribution of Albizia odoratissima (L. f.) Benth. Under Climate Change Based on the Biomod2 Model


Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Data Collection
2.2. Environmental Variables
2.3. Model Calibration
3. Results
3.1. Model Validation
3.2. Selection of Environmental Variables
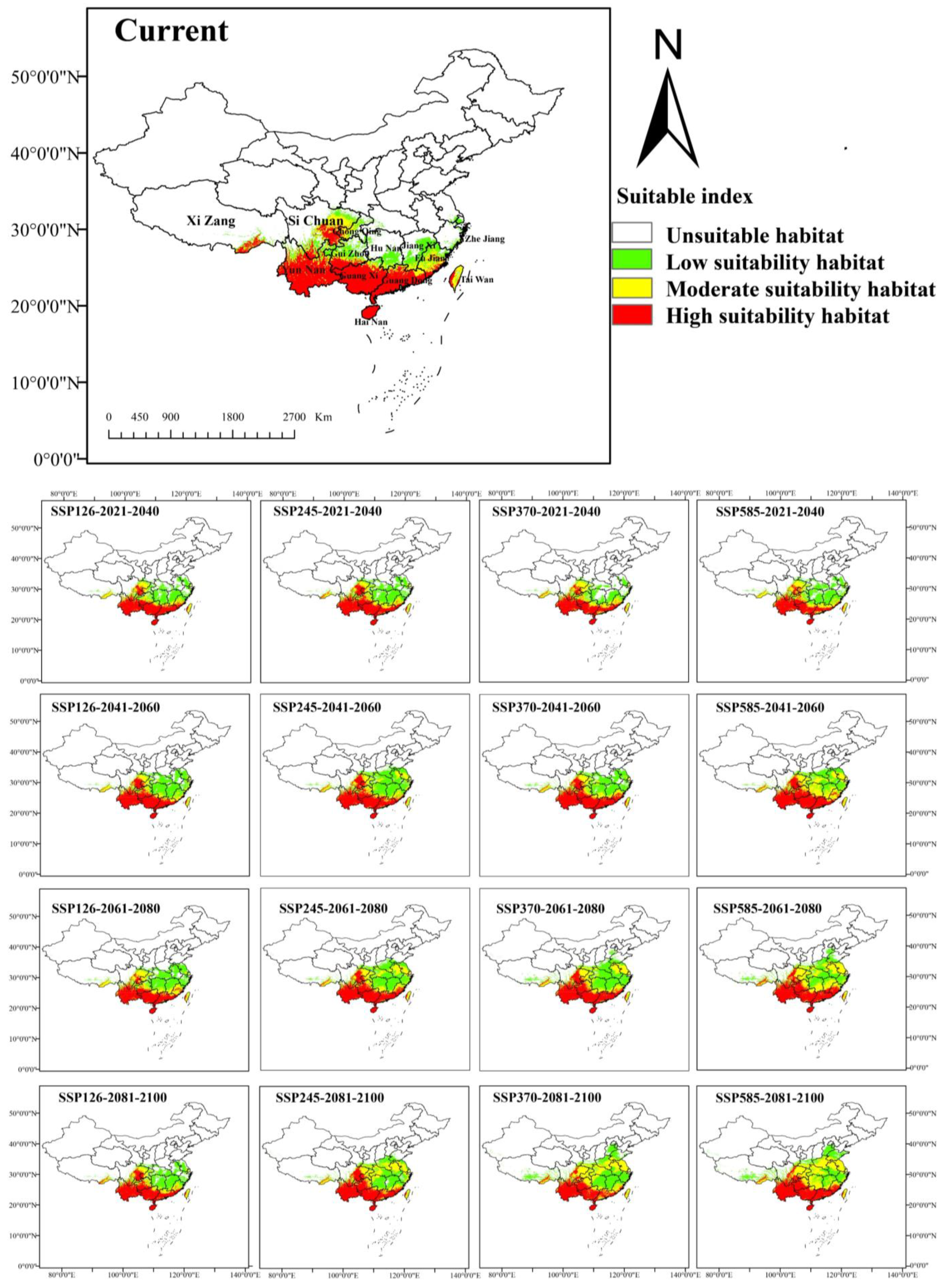
3.3. Current Potential Geographical Distribution in China
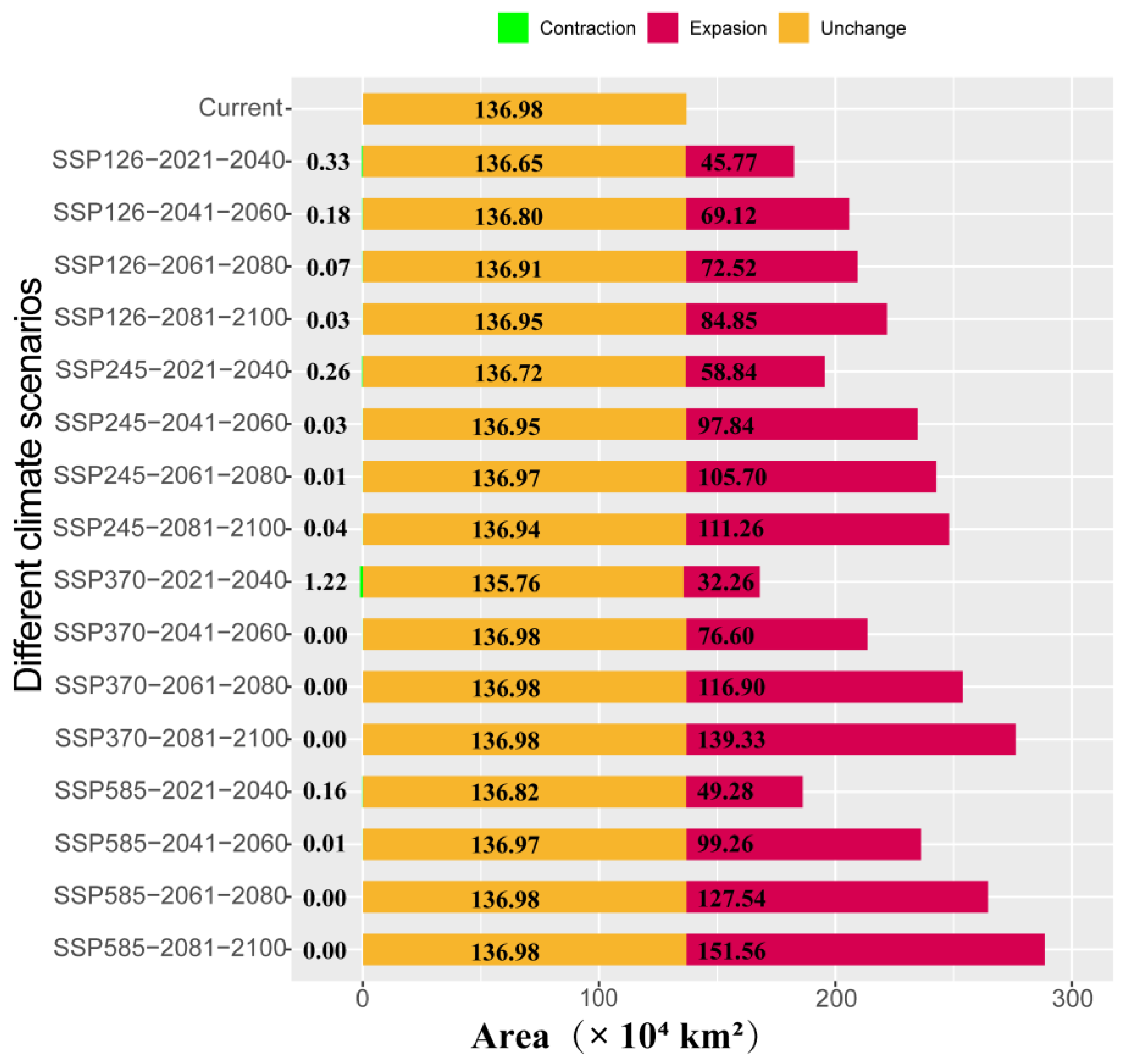
3.4. Distribution Changes Under Future Climate Conditions
4. Discussion
4.1. Evaluation of Modeling Performance
4.2. Critical Environmental Factors Influencing A. odoratissima Distribution
4.3. Impact of Climatic Changes on Suitable Habitats for A. odoratissima
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kelly, A.E.; Goulden, M.L. Rapid shifts in plant distribution with recent climate change. Proc. Natl. Acad. Sci. USA 2008, 105, 11823–11826. [Google Scholar] [CrossRef] [PubMed]
- Kosanic, A.; Anderson, K.; Harrison, S.; Turkington, T.; Bennie, J. Changes in the geographical distribution of plant species and climatic variables on the West Cornwall peninsula (South West UK). PLoS ONE 2018, 13, e0191021. [Google Scholar] [CrossRef] [PubMed]
- Varol, T.; Canturk, U.; Cetin, M.; Ozel, H.B.; Sevik, H. Impacts of climate change scenarios on European ash tree (Fraxinus excelsior L.) in Turkey. For. Ecol. Manag. 2021, 491, 119199. [Google Scholar] [CrossRef]
- Demiral, E.E.; Sağlam, Ü. Sustainable production assessment of the 50 US states. J. Clean. Prod. 2023, 419, 138086. [Google Scholar] [CrossRef]
- Huang, Z.; Xie, L.; Wang, H.; Zhong, J.; Li, Y.; Liu, J.; Ou, Z.; Liang, X.; Li, Y.; Huang, H.; et al. Geographic distribution and impacts of climate change on the suitable habitats of Zingiber species in China. Ind. Crops Prod. 2019, 138, 111429. [Google Scholar] [CrossRef]
- Leimu, R.; Vergeer, P.; Angeloni, F.; Ouborg, N.J. Habitat fragmentation, climate change, and inbreeding in plants. Ann. N. Y. Acad. Sci. 2010, 1195, 84–98. [Google Scholar] [CrossRef] [PubMed]
- Brown, J. SDMtoolbox: A python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. Methods Ecol. Evol. 2014, 5, 694–700. [Google Scholar] [CrossRef]
- Franklin, J. Species distribution models in conservation biogeography: Developments and challenges. Divers. Distrib. 2013, 19, 1217–1223. [Google Scholar] [CrossRef]
- Booth, T.H.; Nix, H.A.; Busby, J.R.; Hutchinson, M.F. Bioclim: The first species distribution modelling package, its early applications and relevance to most current MaxEnt studies. Divers. Distrib. 2014, 20, 1–9. [Google Scholar] [CrossRef]
- Stockman, A.; Beamer, D.; Bond, J. An evaluation of a GARP model as an approach to predicting the spatial distribution of non-vagile invertebrate species. Divers. Distrib. 2006, 12, 81–89. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Aguirre-Gutiérrez, J.; Carvalheiro, L.G.; Polce, C.; van Loon, E.E.; Raes, N.; Reemer, M.; Biesmeijer, J.C. Fit-for-purpose: Species distribution model performance depends on evaluation criteria—Dutch Hoverflies as a case study. PLoS ONE 2013, 8, e63708. [Google Scholar] [CrossRef]
- Arenas-Castro, S.; Regos, A.; Martins, I.; Honrado, J.; Alonso, J. Effects of input data sources on species distribution model predictions across species with different distributional ranges. J. Biogeogr. 2022, 49, 1299–1312. [Google Scholar] [CrossRef]
- Hao, T.; Elith, J.; Guillera-Arroita, G.; Lahoz-Monfort, J.J. A review of evidence about use and performance of species distribution modelling ensembles like BIOMOD. Divers. Distrib. 2019, 25, 839–852. [Google Scholar] [CrossRef]
- Cao, G.; Yuan, X.; Shu, Q.; Gao, Y.; Wu, T.; Xiao, C.; Xu, J.; Zhang, Y. Prediction of the potentially suitable areas of Eucommia ulmoides Oliver in China under climate change based on optimized Biomod2 and MaxEnt models. Front. Plant Sci. 2024, 15, 1359271. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Guo, S.; Ma, C.; Yang, J.; Kang, X.; Li, R. Impact of Climate Change on the Potential Geographical Distribution Patterns of Luculia pinceana Hook. f. since the Last Glacial Maximum. Forests 2024, 15, 253. [Google Scholar] [CrossRef]
- Ghehsareh Ardestani, E.; Heidari Ghahfarrokhi, Z. Ensembpecies distribution modeling of Salvia hydrangea under future climate change scenarios in Central Zagros Mountains, Iran. Glob. Ecol. Conserv. 2021, 26, e01488. [Google Scholar] [CrossRef]
- Sowmya Dhanalakshmi, C.; Kaliappan, S.; Mohammed Ali, H.; Sekar, S.; Depoures, M.V.; Patil, P.P.; Subbaiah, B.S.; Socrates, S.; Birhanu, H.A. Flash Pyrolysis Experiment on Albizia odoratissima Biomass under Different Operating Conditions: A Comparative Study on Bio-Oil, Biochar, and Noncondensable Gas Products. J. Chem. 2022, 2022, 9084029. [Google Scholar] [CrossRef]
- Banothu, V.; Neelagiri, C.; Adepally, U.; Lingam, J.; Bommareddy, K. Phytochemical screening and evaluation of in vitro antioxidant and antimicrobial activities of the indigenous medicinal plant Albizia odoratissima. Pharm. Biol. 2017, 55, 1155–1161. [Google Scholar] [CrossRef] [PubMed]
- Koteswara Rao, Y.; Vijaya Bhaskar Reddy, M.; Venkata Rao, C.; Gunasekar, D.; Blond, A.; Caux, C.; Bodo, B. Two new 5-deoxyflavones from Albizia odoratissima. Chem. Pharm. Bull. 2002, 50, 1271–1272. [Google Scholar] [CrossRef]
- Kumar, D.; Kumar, S.; Kohli, S.; Arya, R.; Gupta, J. Antidiabetic activity of methanolic bark extract of Albizia odoratissima Benth. In alloxan induced diabetic albino mice. Asian Pac. J. Trop. Med. 2011, 4, 900–903. [Google Scholar] [CrossRef]
- Azani, N.; Babineau, M.; Bailey, C.D.; Banks, H.; Barbosa, A.; Barbosa Pinto, R.; Boatwright, J.; Borges, L.; Brown, G.; Bruneau, A.; et al. A new subfamily classification of the Leguminosae based on a taxonomically comprehensive phylogeny. Taxon 2017, 66, 44–77. [Google Scholar] [CrossRef]
- An, Q.; Feng, Y.; Yang, Z.; Hu, L.; Wu, D.; Gong, G. Differences in Albizia odoratissima genetic diversity between Hainan Island and mainland populations in China. Front. Plant Sci. 2024, 15, 1369409. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Gao, F.; Wang, Z.; Yin, G.; Wen, S.; Ou, H.; Liu, Z. Transcriptome and Metabolome Analyses Reveal the Molecular Mechanisms of Albizia odoratissima’s Response to Drought Stress. Plants 2024, 13, 2732. [Google Scholar] [CrossRef] [PubMed]
- Warren, D.; Glor, R.; Turelli, M. ENMTools: A toolbox for comparative studies of environmental niche models. Ecography 2010, 33, 607–611. [Google Scholar] [CrossRef]
- Shi, X.; Yin, Q.; Sang, Z.; Zhu, Z.; Jia, Z.; Ma, L. Prediction of potentially suitable areas for the introduction of Magnolia wufengensis under climate change. Ecol. Indic. 2021, 127, 107762. [Google Scholar] [CrossRef]
- Yu, F.; Wang, T.; Groen, T.A.; Skidmore, A.K.; Yang, X.; Ma, K.; Wu, Z. Climate and land use changes will degrade the distribution of Rhododendrons in China. Sci. Total Environ. 2019, 659, 515–528. [Google Scholar] [CrossRef]
- Kass, J.M.; Muscarella, R.; Galante, P.J.; Bohl, C.L.; Pinilla-Buitrago, G.E.; Boria, R.A.; Soley-Guardia, M.; Anderson, R.P. ENMeval 2.0: Redesigned for customizable and reproducible modeling of species’ niches and distributions. Methods Ecol. Evol. 2021, 12, 1602–1608. [Google Scholar] [CrossRef]
- Muscarella, R.; Galante, P.J.; Soley-Guardia, M.; Boria, R.A.; Kass, J.M.; Uriarte, M.; Anderson, R.P. ENMeval: An R package for conducting spatially independent evaluations and estimating optimal model complexity for MaxEnt ecological niche models. Methods Ecol. Evol. 2014, 5, 1198–1205. [Google Scholar] [CrossRef]
- Wilfried, T.; Damien, G.; Maya, G.; Robin, E.; Frank, B.; Bruno, L.; Remi, P.; Helene, B. biomod2: Ensemble Platform for Species Distribution Modeling. 2024. Available online: https://biomodhub.github.io/biomod2/ (accessed on 30 September 2024).
- Hijmans, R.J.; Phillips, S.; Leathwick, J.; Elith, J. dismo: Species Distribution Modeling. R package version 1.3–14. 2023. Available online: https://CRAN.R-project.org/package=dismo (accessed on 30 September 2024).
- ESRI. ArcGIS Desktop, version 10.7; Environmental Systems Research Institute: Redlands, CA, USA, 2019. [Google Scholar]
- Kaky, E.; Nolan, V.; Alatawi, A.; Gilbert, F. A comparison between Ensemble and MaxEnt species distribution modelling approaches for conservation: A case study with Egyptian medicinal plants. Ecol. Inform. 2020, 60, 101150. [Google Scholar] [CrossRef]
- Norberg, A.; Abrego, N.; Blanchet, F.G.; Adler, F.R.; Anderson, B.J.; Anttila, J.; Araújo, M.B.; Dallas, T.; Dunson, D.; Elith, J.; et al. A comprehensive evaluation of predictive performance of 33 species distribution models at species and community levels. Ecol. Monogr. 2019, 89, e01370. [Google Scholar] [CrossRef]
- Jia, T.; Qi, Y.; Zhao, H.; Xian, X.; Li, J.; Huang, H.; Yu, W.; Liu, W. Estimation of climate-induced increased risk of Centaurea solstitialis L. invasion in China: An integrated study based on biomod2. Front. Ecol. Evol. 2023, 11, 1113474. [Google Scholar] [CrossRef]
- Gu, R.; Wei, S.; Li, J.; Zheng, S.; Li, Z.; Liu, G.; Fan, S. Predicting the impacts of climate change on the geographic distribution of moso bamboo in China based on biomod2 model. Eur. J. For. Res. 2024, 143, 1499–1512. [Google Scholar] [CrossRef]
- Huang, D.; An, Q.; Huang, S.; Tan, G.; Quan, H.; Chen, Y.; Zhou, J.; Liao, H. Biomod2 modeling for predicting the potential ecological distribution of three Fritillaria species under climate change. Sci. Rep. 2023, 13, 18801. [Google Scholar] [CrossRef]
- Fang, J.; Lechowicz, M.J. Climatic limits for the present distribution of beech (Fagus L.) species in the world. J. Biogeogr. 2006, 33, 1804–1819. [Google Scholar] [CrossRef]
- Xie, C.; Li, M.; Chen, L.; Jim, C.Y. Climate-driven changes to the spatial–temporal pattern of endangered tree Toona ciliata Roem. in China. Theor. Appl. Climatol. 2024, 155, 2071–2085. [Google Scholar] [CrossRef]
- Liu, Y.; Tang, G.; Huang, X.; Wei, K.; Wu, S.; Wang, M.; Wang, Y.; Zhang, J.; Wang, Y. Unexpected deep mixing layer in the Sichuan Basin, China. Atmos. Res. 2021, 249, 105300. [Google Scholar] [CrossRef]
- He, Y.; Wang, G.; Ren, Y.; Gao, S.; Chu, D.; McKirdy, S.J. Machine learning ensemble model prediction of northward shift in potato cyst nematodes (Globodera rostochiensis and G. pallida) distribution under climate change conditions. J. Integr. Agric. 2024, 23, 3576–3591. [Google Scholar] [CrossRef]
- Zhao, G.; Cui, X.; Sun, J.; Li, T.; Wang, Q.; Ye, X.; Fan, B. Analysis of the distribution pattern of Chinese Ziziphus jujuba under climate change based on optimized biomod2 and MaxEnt models. Ecol. Indic. 2021, 132, 108256. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | TSS | ROC |
|---|---|---|
| EMmean | 0.894 | 0.972 |
| EMmedian | 0.896 | 0.967 |
| EMca | 0.897 | 0.979 |
| EMwmean | 0.895 | 0.973 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Ji, Q.; Yang, Y.; Gao, Y.; Xu, M.; Guan, Y. Predicting the Potential Suitable Distribution of Albizia odoratissima (L. f.) Benth. Under Climate Change Based on the Biomod2 Model. Biology 2025, 14, 180. https://doi.org/10.3390/biology14020180
Li Z, Ji Q, Yang Y, Gao Y, Xu M, Guan Y. Predicting the Potential Suitable Distribution of Albizia odoratissima (L. f.) Benth. Under Climate Change Based on the Biomod2 Model. Biology. 2025; 14(2):180. https://doi.org/10.3390/biology14020180
Chicago/Turabian StyleLi, Zhiting, Qiaomiao Ji, Yong Yang, Yunfei Gao, Meng Xu, and Yali Guan. 2025. "Predicting the Potential Suitable Distribution of Albizia odoratissima (L. f.) Benth. Under Climate Change Based on the Biomod2 Model" Biology 14, no. 2: 180. https://doi.org/10.3390/biology14020180
APA StyleLi, Z., Ji, Q., Yang, Y., Gao, Y., Xu, M., & Guan, Y. (2025). Predicting the Potential Suitable Distribution of Albizia odoratissima (L. f.) Benth. Under Climate Change Based on the Biomod2 Model. Biology, 14(2), 180. https://doi.org/10.3390/biology14020180