
Exogenous Glycinebetaine Regulates the Contrasting Responses in Leaf Physiochemical Attributes and Growth of Maize under Drought and Flooding Stresses
 ,
,  ,
,
Abstract
Simple Summary
Abstract

1. Introduction
2. Materials and Methods
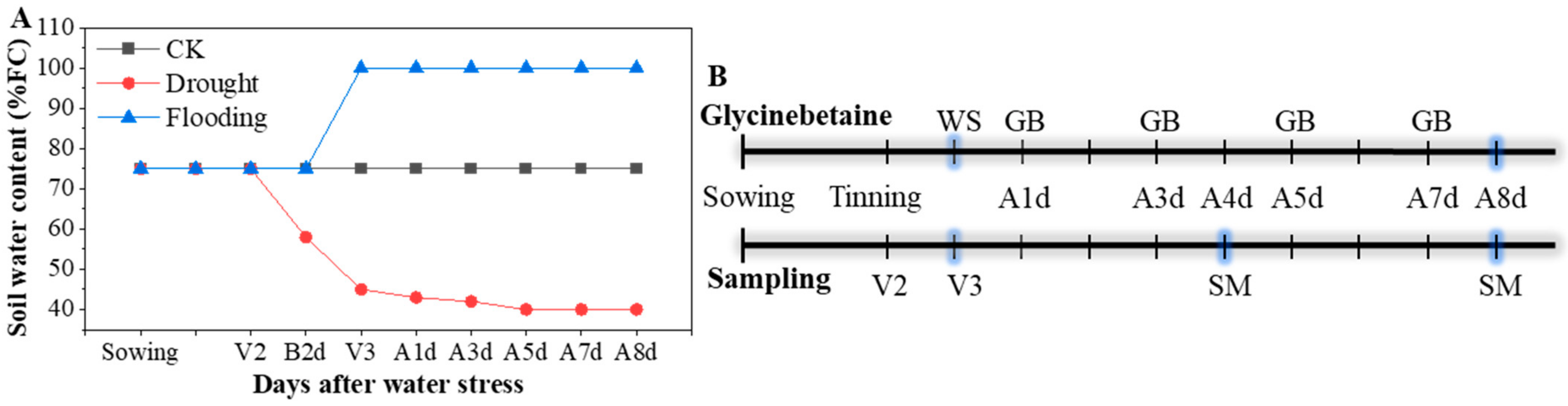
2.1. Plant Materials and Experimental Design
2.2. Sampling and Measurements
2.2.1. Determination of Reactive Oxygen Species
2.2.2. Determination of SOD, POD, APX Activities and Proline Content
2.2.3. Assessment of Morphological Characteristics and Crop Biomass
2.2.4. Measurement of Leaf Relative Water Content
2.2.5. Tolerance Analysis
2.3. Statistical Analysis
3. Results
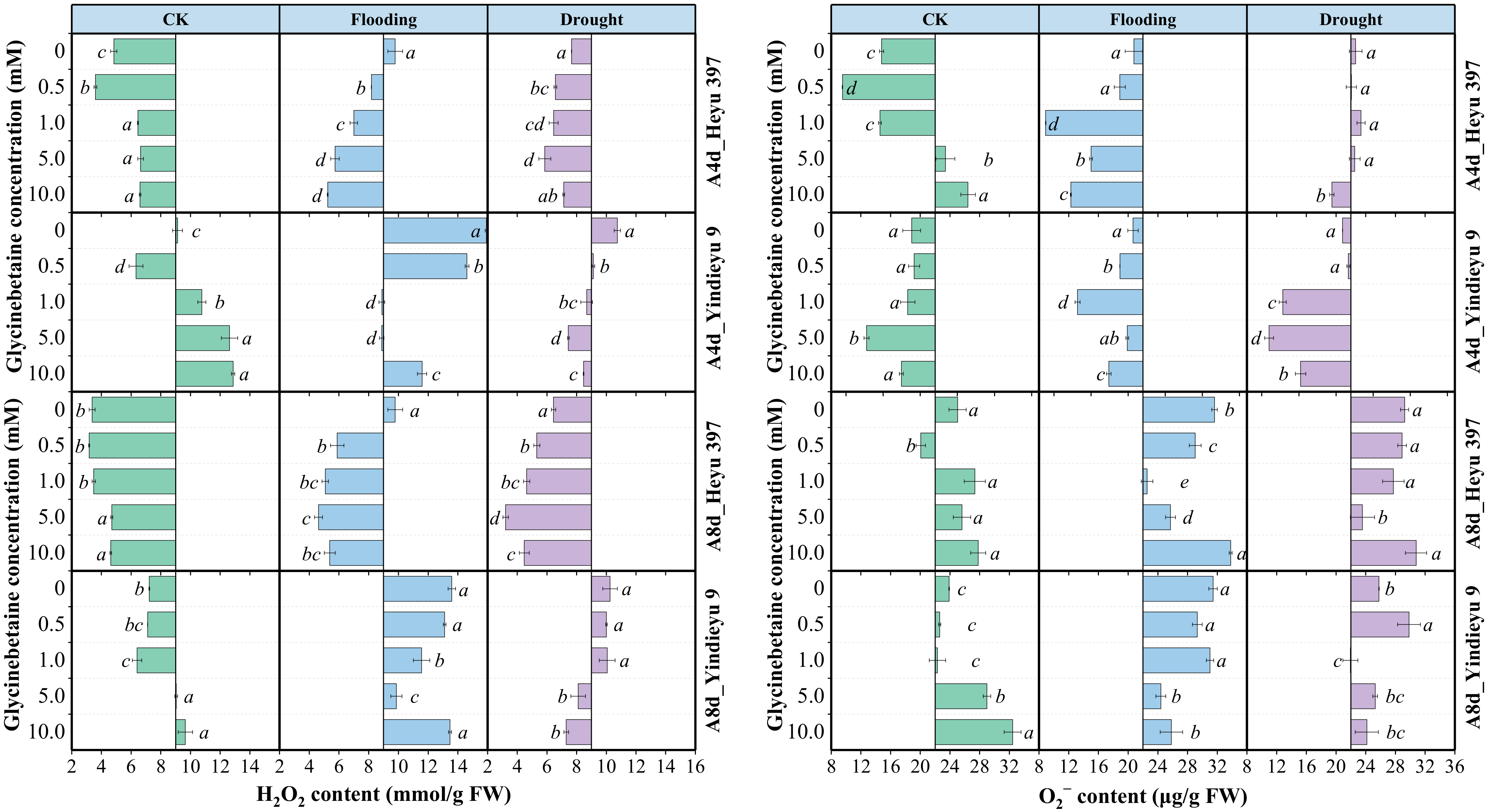
3.1. Glycinebetaine Reduced the Accumulation of Reactive Oxygen Species under Water Stress
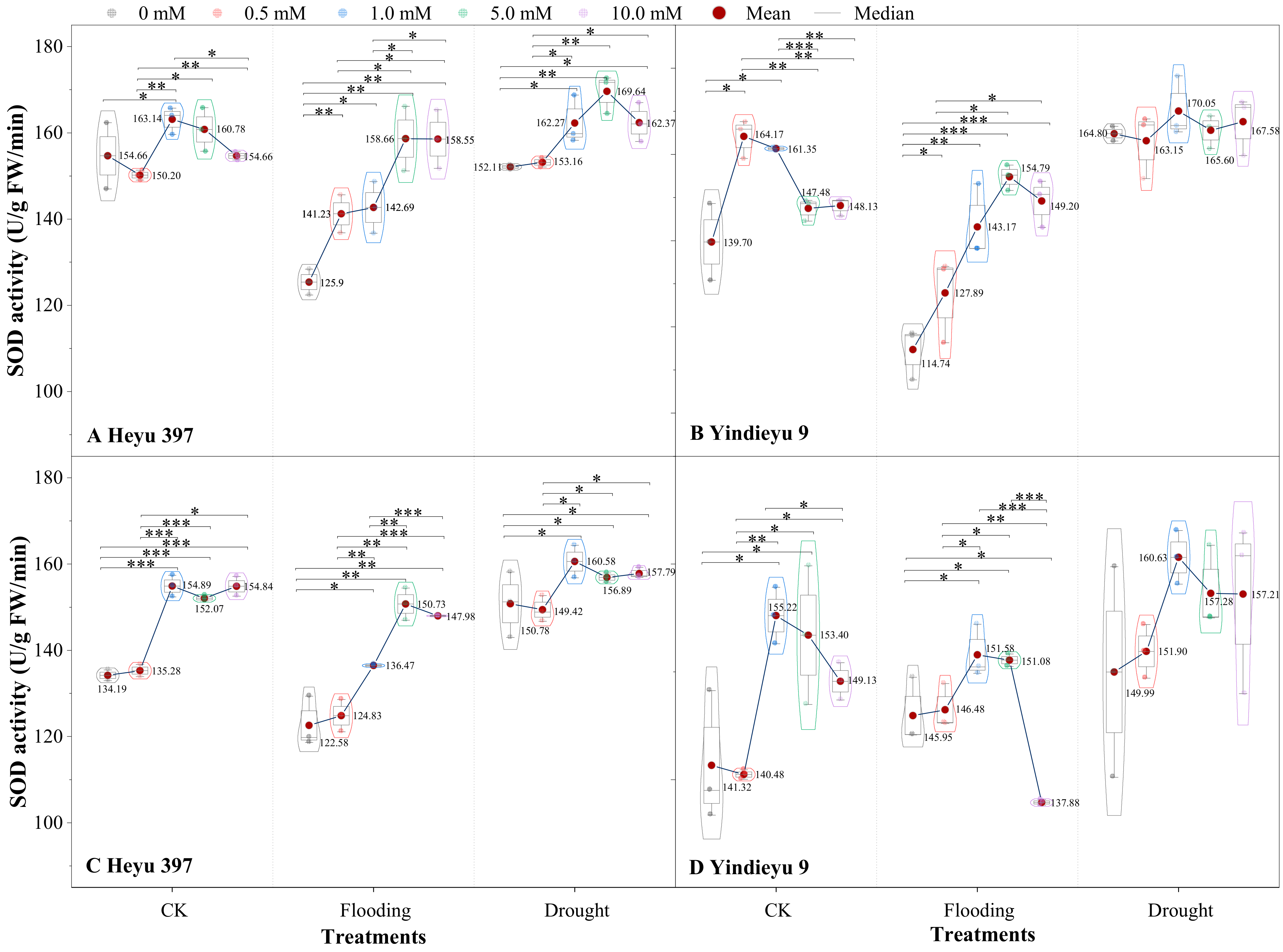
3.2. Glycinebetaine Was More Beneficial to Improving SOD Activity under Flooding Stress
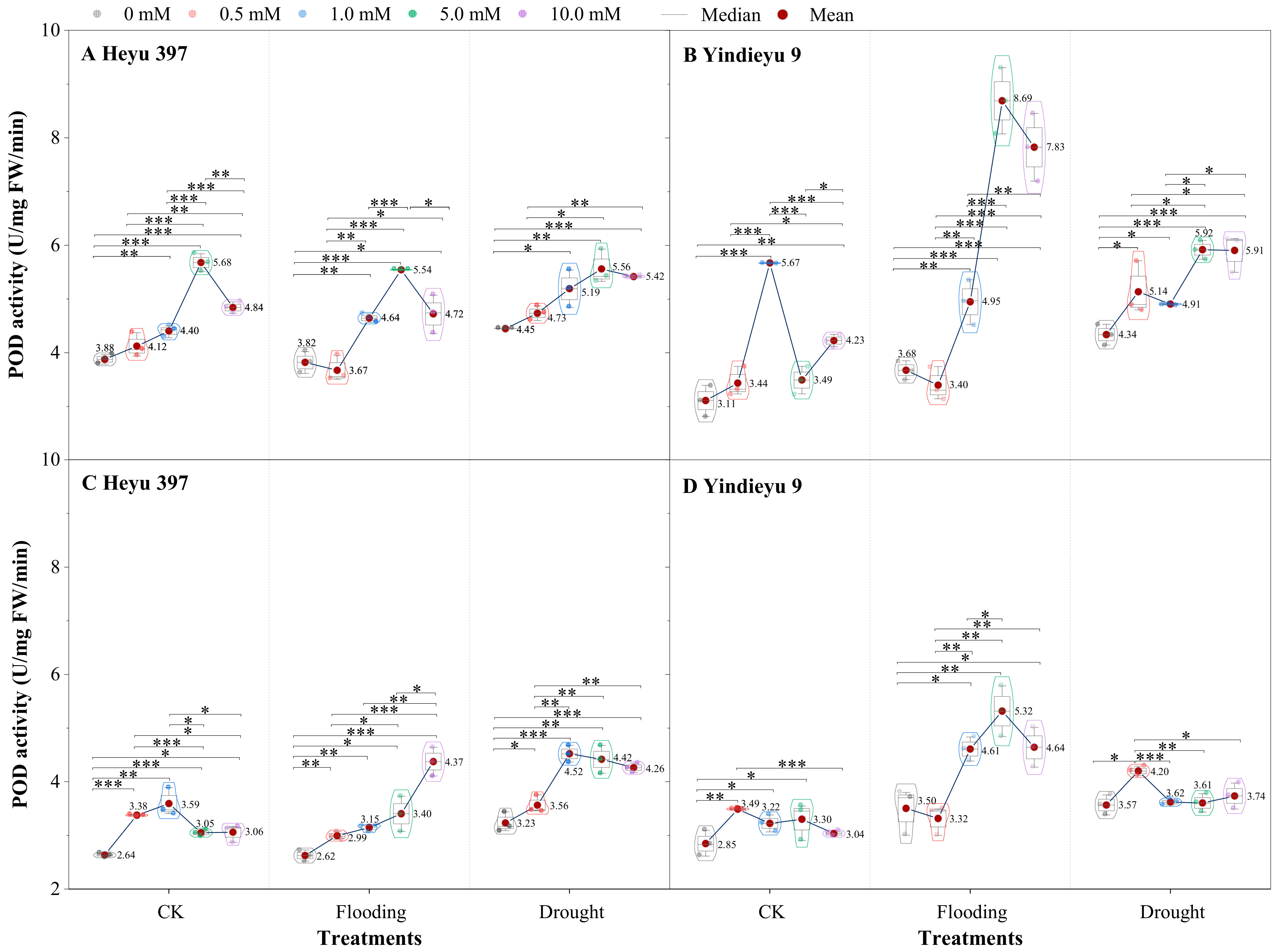
3.3. Glycinebetaine Enhanced the POD Activity under Water Stress
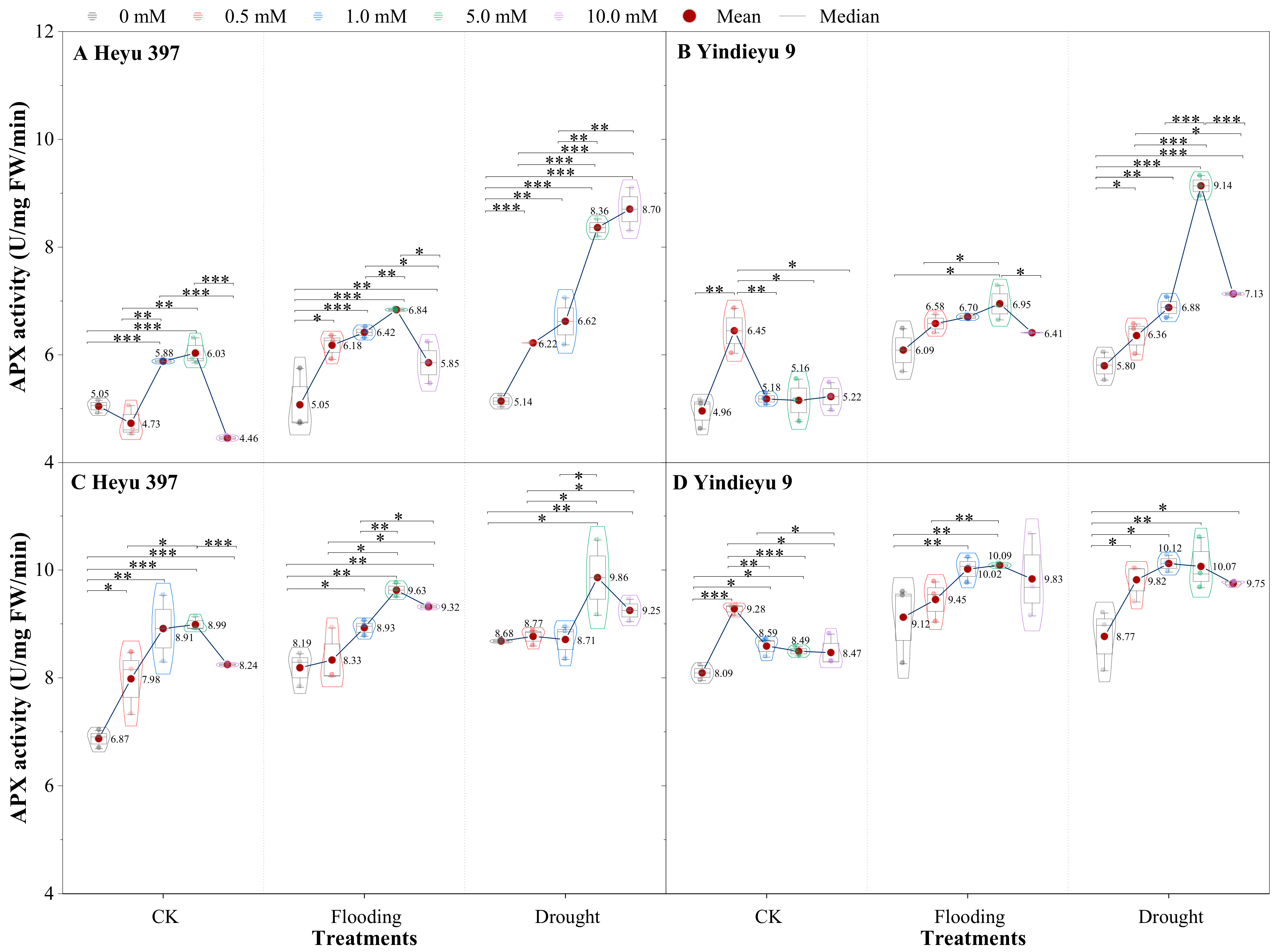
3.4. Glycinebetaine Improved the APX Activity under Water Stress
3.5. Lower Dosage of Glycinebetaine Increased the Proline Content under Water Stress
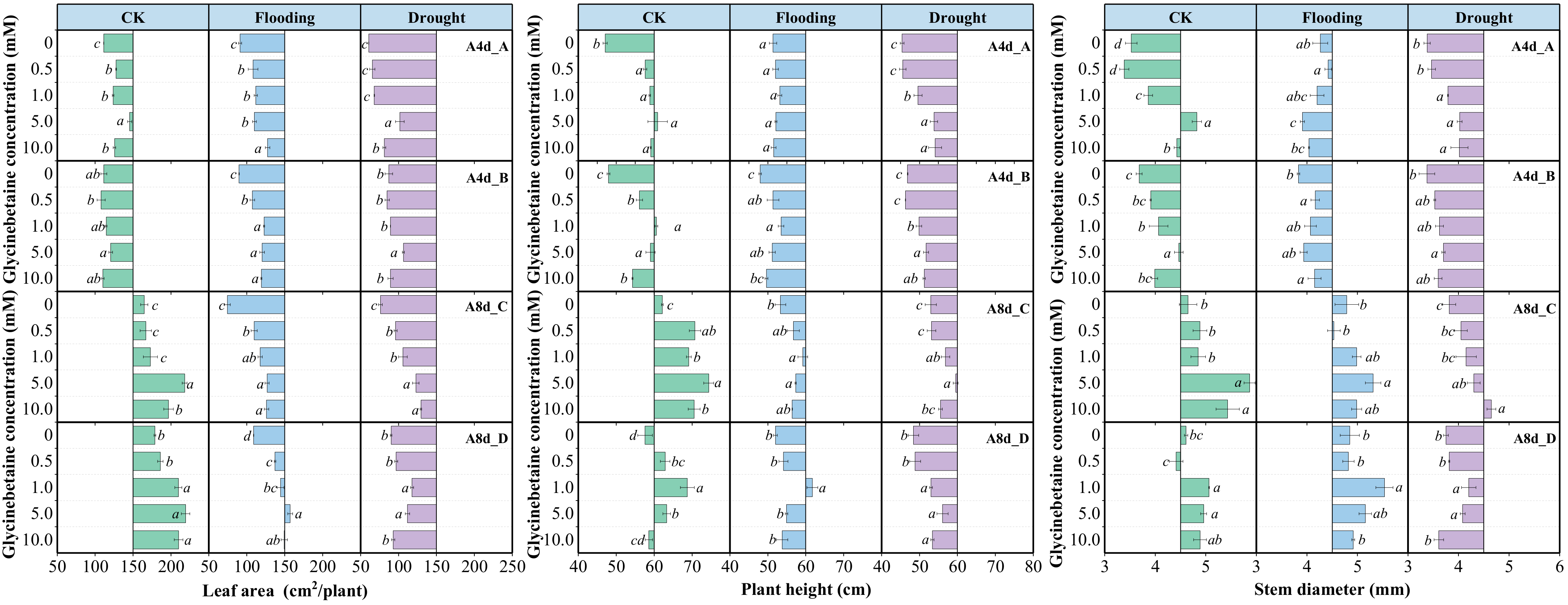
3.6. Glycinebetaine Promoted the Growth of Maize Seedlings under Water Stress
3.7. Glycinebetaine Increased the Biomass Accumulation of Maize Seedlings under Water Stress
3.8. Glycinebetaine Improved Leaf Relative Water Content under Drought Stress
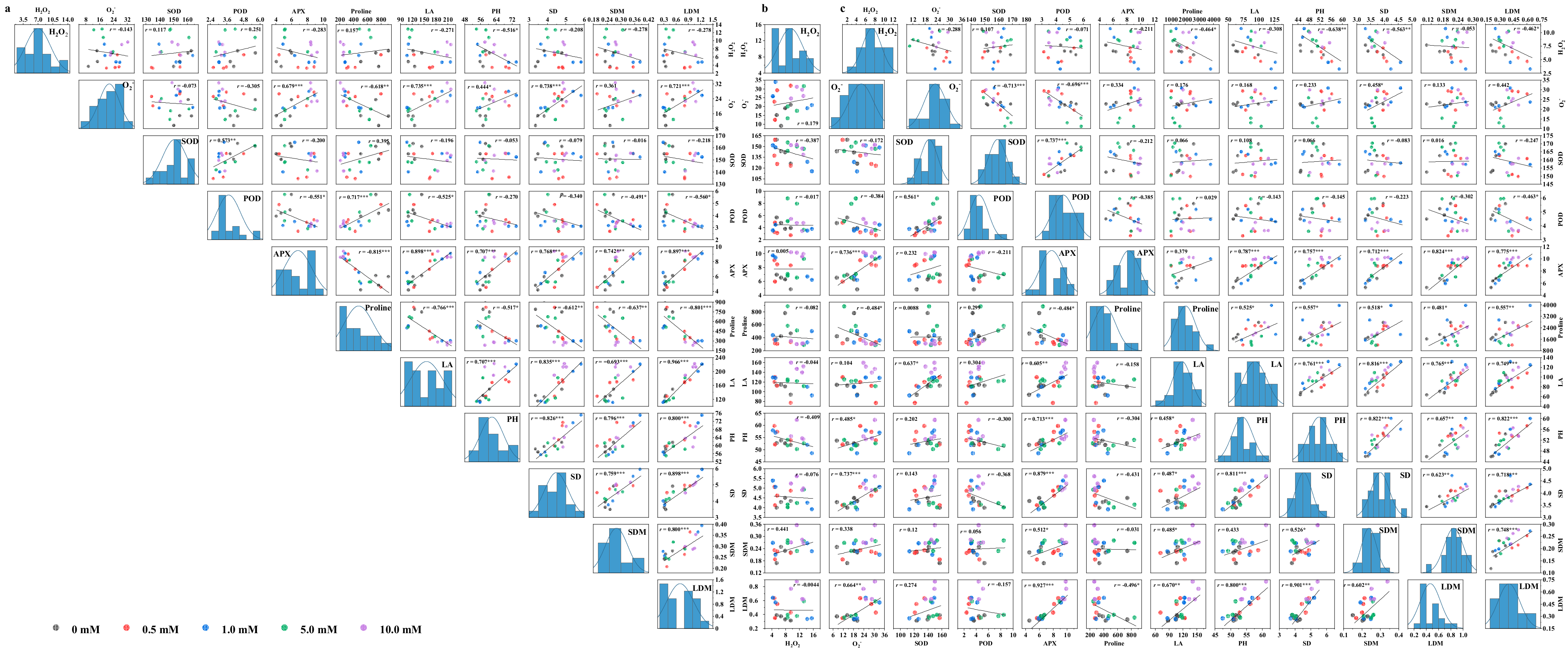
3.9. Analysis for Correlation, Stepwise Regression, and Tolerance
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| GB | Glycinebetaine |
| WS | Water stress |
| ROS | Reactive oxygen species |
| LA | Leaf area |
| SOD | Superoxide dismutase |
| POD | Guaiacol peroxidase |
| APX | Ascorbate peroxidase |
| V3 | Three-leaf stage |
| V5 | Five-leaf stage |
| H2O2 | Hydrogen peroxide |
| O2− | Superoxide anion |
| RWC | Relative water content |
| CK | Maize planted in normal field capacity |
| FC | Field capacity |
| PBS | Phosphate buffer solution |
| OD | Absorbance of the mixture |
| FW | Fresh weight |
| D value | Comprehensive evaluation value |
References
- EM-DAT. 2022. Available online: https://www.emdat.be/publications/ (accessed on 30 April 2023).
- Wang, B.C.H.; Lian, J.; Peng, Y.; Hu, B.; Chen, H. Generalized reference evapotranspiration models with limited climatic data based on random forest and gene expression programming in Guangxi, China. Agric. Water Manag. 2019, 221, 220–230. [Google Scholar] [CrossRef]
- Wang, G.Y.; Ahmad, S.; Wang, Y.; Wang, B.W.; Huang, J.H.; Jahan, M.S.; Zhou, X.B.; Shi, C.Q. Multivariate analysis compares and evaluates drought and flooding tolerances of maize germplasm. Plant Physiol. 2023, 193, 339–355. [Google Scholar] [CrossRef] [PubMed]
- Ren, B.; Zhang, J.; Dong, S.; Zhao, B. Responses of carbon metabolism and antioxidant system of summer maize to waterlogging at different stages. J. Agron. Crop Sci. 2018, 204, 505–514. [Google Scholar] [CrossRef]
- Das, R.R.; Vinayan, M.T.; Seetharam, K.; Patel, M.; Phagna, R.K.; Singh, S.B.; Shahi, J.P.; Sarma, A.; Barua, N.S.; Babu, R.; et al. Genetic gains with genomic versus phenotypic selection for drought and waterlogging tolerance in tropical maize (Zea mays L.). Crop J. 2021, 9, 1438–1448. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Mahmud, J.A.; Fujita, M.; Fotopoulos, V. Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants 2022, 9, 681. [Google Scholar] [CrossRef]
- Ahmad, S.; Wang, G.Y.; Muhammad, I.; Chi, Y.X.; Zeeshan, M.; Nasar, J.; Zhou, X.B. Interactive effects of melatonin and nitrogen improve drought tolerance of maize seedlings by regulating growth and physiochemical attributes. Antioxidants 2022, 11, 359. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Zhang, W.; Wang, H.; Gao, Y.; Ma, S.; Qin, A.; Liu, Z.; Zhao, B.; Ning, D.; Zheng, H. Effects of waterlogging at different stages on growth and ear quality of waxy maize. Agric. Water Manag. 2022, 261, 107334. [Google Scholar] [CrossRef]
- Barickman, T.C.; Simpson, C.R.; Sams, C.E. Waterlogging causes early modification in the physiological performance, carotenoids, chlorophylls, proline, and soluble sugars of cucumber plants. Plants 2019, 8, 160. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Feng, Y.; Zong, Y.; Zhang, D.; Li, P. Elevated CO2-induced changes in photosynthesis, antioxidant enzymes and signal transduction enzyme of soybean under drought stress. Plant Physiol. Bioch. 2020, 154, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Lukić, N.; Trifković, T.; Kojić, D.; Kukavica, B. Modulations of the antioxidants defence system in two maize hybrids during flooding stress. J. Plant Res. 2021, 134, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Salah, A.; Nwafor, C.C.; Han, Y.; Liu, L.; Rashid, M.; Batool, M.; El-Badri, A.M.; Cao, C.; Zhan, M. Spermidine and brassinosteroid regulate root anatomical structure, photosynthetic traits and antioxidant defense systems to alleviate waterlogging stress in maize seedlings. S. Afr. J. Bot. 2022, 144, 389–402. [Google Scholar] [CrossRef]
- Azahar, I.; Ghosh, S.; Adhikari, A.; Adhikari, S.; Hossain, Z. Comparative analysis of maize root sRNA transcriptome unveils the regulatory roles of miRNAs in submergence stress response mechanism. Environ. Exp. Bot. 2020, 171, 103924. [Google Scholar] [CrossRef]
- Qi, M.; Liu, X.; Li, Y.; Song, H.; Zhou, G. Photosynthetic resistance and resilience under drought, flooding and rewatering in maize plants. Photosynth. Res. 2021, 148, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, X.; Gu, J.; Shi, W.; Zhao, H.; Sun, C.; You, S. Drought and waterlogging status and dominant meteorological factors affecting maize (Zea mays L.) in different growth and development stages in northeast China. Agronomy 2023, 13, 374. [Google Scholar] [CrossRef]
- Tian, L.X.; Bi, W.S.; Ren, X.S.; Li, W.L.; Li, J. Flooding has more adverse effects on the stem structure and yield of spring maize (Zea mays L.) than waterlogging in northeast China. Eur. J. Agron. 2020, 117, 126054. [Google Scholar] [CrossRef]
- Tian, L.; Li, J.; Bi, W.; Zuo, S.; Li, L.; Li, W.; Sun, L. Effects of waterlogging stress at different growth stages on the photosynthetic characteristics and grain yield of spring maize (Zea mays L.) under field conditions. Agric. Water Manag. 2019, 218, 250–258. [Google Scholar] [CrossRef]
- Jia, Y.; Xiao, W.; Ye, Y.; Wang, X.; Wang, Y. Response of photosynthetic performance to drought duration and re-watering in maize. Agronomy 2020, 10, 533. [Google Scholar] [CrossRef]
- Shemi, R.; Wang, L.; Zhang, K.; Gheith, E.S.M.S. Effects of salicylic acid, zinc and glycine betaine on morphophysiological growth and yield of maize under drought stress. Sci. Rep. 2021, 11, 3195. [Google Scholar] [CrossRef] [PubMed]
- Sadaghiani, F.M.; Dehaghi, M.A.; Pirzad, A.; Fotokian, M.H. Variation in yield and biochemical factors of German chamomile (Matricaria recutita L.) under foliar application of osmolytes and drought stress conditions. J. Herbmed. Pharmacol. 2019, 8, 90–100. [Google Scholar] [CrossRef]
- Hamani, A.K.M.; Li, S.; Chen, J.; Amin, A.S.; Wang, G.; Xiaojun, S.; Zain, M.; Gao, Y. Linking exogenous foliar application of glycine betaine and stomatal characteristics with salinity stress tolerance in cotton (Gossypium hirsutum L.) seedlings. BMC Plant Biol. 2021, 21, 146. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Leon, S.G.; Valenzuela-Soto, E.M. Glycine betaine is a phytohormone-like plant growth and development regulator under stress conditions. J. Plant Growth Regul. 2022, 42, 5029–5040. [Google Scholar] [CrossRef]
- Nawaz, M.; Wang, Z. Abscisic acid and glycine betaine mediated tolerance mechanisms under drought stress and recovery in Axonopus compressus, a new insight. Sci. Rep. 2020, 10, 6942. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela-Soto, E.M.; Figueroa-Soto, C.G. Biosynthesis and degradation of glycine betaine and its potential to control plant growth and development. In Osmoprotectant-Mediated Abiotic Stress Tolerance in Plants: Recent Advances and Future Perspectives; Kumar, V., Burritt, D.J., Fujita, M., Mäkela, P., Hossain, M.A., Eds.; Springer Nature: Cham, Switzerland, 2019; pp. 123–140. [Google Scholar]
- Elhakem, A.H. Mitigation of the salinity influences on maize (Zea mays L.) productivity by exogenous applications of glycine betaine. J. Stress Physiol. Biochem. 2019, 15, 21–28. [Google Scholar]
- Ahmed, N.; Zhang, Y.; Li, K.; Zhou, Y.; Zhang, M.; Li, Z. Exogenous application of glycine betaine improved water use efficiency in winter wheat (Triticum aestivum L.) via modulating photosynthetic efficiency and antioxidative capacity under conventional and limited irrigation conditions. Crop J. 2019, 7, 65–80. [Google Scholar] [CrossRef]
- Rasheed, R.; Iqbal, M.; Ashraf, M.A.; Hussain, I.; Shafiq, F.; Yousaf, A.; Zaheer, A. Glycine betaine counteracts the inhibitory effects of waterlogging on growth, photosynthetic pigments, oxidative defence system, nutrient composition, and fruit quality in tomato. J. Hortic. Sci. Biotech. 2018, 93, 385–391. [Google Scholar] [CrossRef]
- Mäkelä, P.; Peltonen-Sainio, P.; Jokinen, K.; Pehu, E.; Setala, H.; Hinkkanen, R.; Somersalo, S. Uptake and translocation of foliar-applied glycinebetaine in crop plants. Plant Sci. 1996, 1, 221–230. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Banerjee, A.; Bhuyan, M.H.M.B.; Roychoudhury, A.; Mahmud, J.A.; Fujita, M. Targeting glycine betaine for abiotic stress tolerance in crop plants, physiological mechanism, molecular interaction and signaling. Phyton Int. J. Exp. Bot. 2019, 88, 185–221. [Google Scholar] [CrossRef]
- Chen, F.; Fang, P.; Zeng, W.; Ding, Y.; Peng, Y. Comparing transcriptome expression profiles to reveal the mechanisms of salt tolerance and exogenous glycine betaine mitigation in maize seedlings. PLoS ONE 2020, 15, e0233616. [Google Scholar] [CrossRef] [PubMed]
- Sharma, J.; Kumar, S.; Kumar, N.; Yadav, N.; Khyalia, P.; Sharma, A. Evaluation of yield and quality attributes of barley cultivars with foliar spray of glycine betaine under lead toxicity. Cereal Res. Commun. 2024. [Google Scholar] [CrossRef]
- Yang, X.H.; Lu, C.M. Effects of exogenous glycinebetaine on growth, CO2 assimilation, and photosystem II photochemistry of maize plants. Physiol. Plantarum 2006, 127, 593–602. [Google Scholar] [CrossRef]
- Huang, Y.; Li, J.; Duan, L.; Li, Z. Drought resistance of maize seedlings induced by betaine. J. Maize Sci. 2011, 19, 95–100, (In Chinese with English abstract). [Google Scholar]
- Yang, X.; Song, T.; Liu, H.; Wang, H. Effects of exogenous glycine betaine on the growth and chlorophyll content of maize seedlings under NaCl stress. Hubei Agr. Sci. 2017, 56, 830–833+875, (In Chinese with English abstract). [Google Scholar]
- Lukatkin, A.S. Contribution of oxidative stress to the development of cold-induced damage to leaves of chilling-sensitive plants: Reactive oxygen species formation during plant chilling. Russ. J. Plant Physiol. 2002, 49, 622–627. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutases: I. occurrence in higher plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Cakmak, I.; Marschner, H. Magnesium deficiency and high light intensity enhance activities of superoxide dismutase ascorbate peroxidase and glutathione reductase in bean leaves. Plant Physiol. 1992, 98, 1222–1227. [Google Scholar] [CrossRef] [PubMed]
- Nakano, Y.; Asada, K. Purification of ascorbate peroxidase in spinach chloroplasts; its inactivation in ascorbate-depleted medium and reactivation by monodehydroascorbate radical. Plant Cell Physiol. 1987, 28, 131–140. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Ahmad, S.; Wang, G.Y.; Muhammad, I.; Farooq, S.; Kamran, M.; Ahmad, I.; Zeeshan, M.; Huang, J.H.; Zhou, X.B. Application of melatonin-mediated modulation of drought tolerance by regulating photosynthetic efficiency, chloroplast ultrastructure, and endogenous hormones in maize. Chem. Biol. Technol. Agric. 2022, 9, 5. [Google Scholar] [CrossRef]
- Zou, J.; Hu, W.; Li, Y.; He, J.; Zhu, H.; Zhou, Z. Screening of drought resistance indices and evaluation of drought resistance in cotton (Gossypium hirsutum L.). J. Integr. Agric. 2020, 19, 495–508. [Google Scholar] [CrossRef]
- Fang, W.; Huang, S.; Huang, G.; Huang, Q.; Wang, H.; Wang, L.; Zhang, Y.; Li, P.; Ma, L. Copulas-based risk analysis for inter-seasonal combinations of wet and dry conditions under a changing climate. Int. J. Climatol. 2019, 39, 2005–2021. [Google Scholar] [CrossRef]
- Bi, W.; Weng, B.; Yan, D.; Wang, H.; Wang, M.; Yan, S.; Jing, L.; Liu, T.; Chang, W. Responses of phosphate-solubilizing microorganisms mediated phosphorus cycling to drought-flood abrupt alternation in summer maize field soil. Front. Microbiol. 2022, 12, 768921. [Google Scholar] [CrossRef] [PubMed]
- Prazeres, C.S.; Coelho, C.M.M.; Souza, C.A. Biochemical compounds and enzymatic systems related to tolerance to water deficit of maize seedlings. Plant Physiol. Rep. 2021, 26, 402–411. [Google Scholar] [CrossRef]
- Salah, A.; Ming, Z.; Cao, C.G.; Han, Y.L.; Ling, L.; Liu, Z.H.; Li, P.; Ye, M.; Jiang, Y. γ-aminobutyric acid promotes chloroplast ultrastructure antioxidant capacity and growth of waterlogged maize seedlings. Sci. Rep. 2019, 9, 484. [Google Scholar] [CrossRef]
- Sarkar, B.; Bandyopadhyay, P.; Das, A.; Pal, S.; Hasanuzzaman, M.; Adak, M.K. Abscisic acid priming confers salt tolerance in maize seedlings by modulating osmotic adjustment, bond energies, ROS homeostasis, and organic acid metabolism. Plant Physiol. Bioch. 2023, 202, 107980. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, G.; Dong, H.; Li, C. Waterlogging stress in cotton: Damage, adaptability, alleviation strategies, and mechanisms. Crop J. 2020, 9, 257–270. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Ahmed, N.; Saha, T.; Rahman, M.; Rahman, K.; Alam, M.M.; Rohman, M.M. Exogenous salicylic acid and kinetin modulate reactive oxygen species metabolism and glyoxalase system to confer waterlogging stress tolerance in soybean (Glycine max L.). Plant Stress 2022, 3, 100057. [Google Scholar] [CrossRef]
- Ren, B.; Yu, W.; Liu, P.; Zhao, B.; Zhang, J. Responses of photosynthetic characteristics and leaf senescence in summer maize to simultaneous stresses of waterlogging and shading. Crop J. 2022, 11, 9. [Google Scholar] [CrossRef]
- Xiong, Q.; Cao, C.; Shen, T.; Zhong, L.; He, H.H.; Chen, X. Comprehensive metabolomic and proteomic analysis in biochemical metabolic pathways of rice spikes under drought and submergence stress. Biochim.Biophys. Acta Proteins Proteom. 2019, 1867, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Jinsheng, Y.; Wenjie, G.; Jiwang, Z.; Baizhao, R.; Lichun, W. Responses of the lodging resistance of summer maize with different gene types to plant density. Agronomy 2022, 12, 10. [Google Scholar] [CrossRef]
- Li, H.; Li, Z.; Shen, Z.J.; Luo, M.R.; Zheng, H.L. Physiological and proteomic responses of mangrove plant Avicennia marina seedlings to simulated periodical inundation. Plant Soil 2020, 450, 231–254. [Google Scholar] [CrossRef]
- Mcgee, T.; Shahid, M.A.; Beckman, T.; Chaparro, J.X.; Sarkhosh, A. Physiological and biochemical characterization of six prunus rootstocks in response to flooding. Environ. Exp. Bot. 2021, 183, 104368. [Google Scholar] [CrossRef]
- Anjum, S.A.; Ashraf, U.; Tanveer, M.; Khan, I.; Hussain, S.; Shahzad, B.; Zohaib, A.; Abbas, F.; Saleem, M.F.; Ali, I.; et al. Drought induced changes in growth, osmolyte accumulation and antioxidant metabolism of three maize hybrids. Front. Plant Sci. 2017, 8, 69. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.L.; Chen, Q.; Chen, Q.J.; Jiang, M.; Qu, Y.Y. Screening of key drought tolerance indices for cotton at the flowering and boll setting stage using the dimension reduction method. Front. Plant Sci. 2021, 12, 619926. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Lyu, W.; Gao, Y.; Zhang, X.; Sun, Y.; Huang, B. Choline-mediated lipid reprogramming as a dominant salt tolerance mechanism in grass species lacking glycine betaine. Plant Cell Physiol. 2021, 61, 2018–2030. [Google Scholar] [CrossRef]
- Qi, X.; Li, Q.; Ma, X.; Qian, C.; Wang, H.; Ren, N.; Shen, C.; Huang, S.; Xu, X.; Xu, Q.; et al. Waterlogging-induced adventitious root formation in cucumber is regulated by ethylene and auxin through reactive oxygen species signalling. Plant Cell Environ. 2019, 42, 1458–1470. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Water Treatments | GB (mM) | After 4 d | After 8 d | ||
|---|---|---|---|---|---|
| Heyu 397 | Yindieyu 9 | Heyu 397 | Yindieyu 9 | ||
| CK | 0 | 514.03 ± 53.28 d | 516.97 ± 32.46 b | 433.06 ± 48.68 a | 351.74 ± 59.51 a |
| 0.5 | 773.46 ± 13.20 bc | 484.00 ± 29.75 b | 312.15 ± 19.12 bc | 264.64 ± 39.26 b | |
| 1.0 | 914.96 ± 5.76 a | 658.77 ± 22.34 a | 333.71 ± 10.26 b | 257.60 ± 31.10 b | |
| 5.0 | 731.70 ± 20.56 c | 636.56 ± 49.11 a | 286.30 ± 2.85 c | 263.65 ± 35.82 b | |
| 10.0 | 805.06 ± 52.51 b | 496.00 ± 15.93 b | 285.09 ± 11.78 c | 218.50 ± 22.50 b | |
| Flooding | 0 | 450.08 ± 51.47 b | 483.93 ± 2.55 c | 315.87 ± 44.52 bc | 271.27 ± 1.07 b |
| 0.5 | 765.04 ± 90.03 a | 310.40 ± 24.91 d | 289.94 ± 14.43 c | 290.44 ± 28.11 b | |
| 1.0 | 408.71 ± 5.24 b | 868.75 ± 49.61 a | 353.37 ± 17.67 b | 285.66 ± 2.24 b | |
| 5.0 | 359.11 ± 23.79 bc | 567.70 ± 44.73 b | 424.42 ± 3.39 a | 333.84 ± 8.23 a | |
| 10.0 | 296.97 ± 51.38 c | 495.31 ± 49.33 c | 344.03 ± 46.18 bc | 295.52 ± 15.18 b | |
| Drought | 0 | 1545.92 ± 175.38 d | 1472.65 ± 13.69 c | 1128.34 ± 42.66 d | 1109.00 ± 28.02 e |
| 0.5 | 1661.98 ± 37.12 cd | 2875.10 ± 65.66 a | 2477.36 ± 225.42 b | 1963.93 ± 35.93 c | |
| 1.0 | 1863.65 ± 13.49 c | 1367.66 ± 20.64 d | 2450.63 ± 41.92 b | 2378.21 ± 72.96 b | |
| 5.0 | 2708.21 ± 108.50 a | 1729.74 ± 74.63 b | 3904.41 ± 33.75 a | 2694.72 ± 58.82 a | |
| 10.0 | 2240.05 ± 195.17 b | 1244.03 ± 30.90 e | 1861.99 ± 12.24 c | 1821.16 ± 70.37 d | |
| Source of Variation | p > F | ||||
| Variety | <0.0001 | <0.0001 | |||
| Water | <0.0001 | <0.0001 | |||
| GB | <0.0001 | <0.0001 | |||
| Variety × Water | <0.0001 | <0.0001 | |||
| Variety × GB | <0.0001 | <0.0001 | |||
| Water × GB | <0.0001 | <0.0001 | |||
| Variety × Water × GB | <0.0001 | <0.0001 | |||
| Water Treatments | GB (mM) | Heyu 397 | Yindieyu 9 |
|---|---|---|---|
| CK | 0 | 95.39 ± 1.98 a | 97.12 ± 0.35 ab |
| 0.5 | 97.48 ± 0.21 a | 96.59 ± 1.60 ab | |
| 1.0 | 96.80 ± 1.25 a | 98.35 ± 1.45 a | |
| 5.0 | 96.72 ± 1.59 a | 96.16 ± 0.79 b | |
| 10.0 | 96.71 ± 1.89 a | 95.65 ± 0.34 b | |
| Flooding | 0 | 97.13 ± 0.67 ab | 95.84 ± 1.94 a |
| 0.5 | 95.18 ± 1.80 b | 95.79 ± 0.59 a | |
| 1.0 | 97.44 ± 0.52 a | 96.35 ± 0.58 a | |
| 5.0 | 95.75 ± 0.41 ab | 97.23 ± 0.72 a | |
| 10.0 | 95.44 ± 1.43 ab | 97.19 ± 0.63 a | |
| Drought | 0 | 68.04 ± 1.73 c | 71.50 ± 0.53 c |
| 0.5 | 80.12 ± 1.64 b | 78.26 ± 2.66 bc | |
| 1.0 | 87.54 ± 0.69 a | 80.94 ± 0.57 ab | |
| 5.0 | 87.10 ± 4.45 a | 82.61 ± 2.78 a | |
| 10.0 | 79.76 ± 1.86 b | 82.04 ± 3.62 ab | |
| Source of Variation | p > F | ||
| Variety | 0.3565 | ||
| Water | <0.0001 | ||
| GB | <0.0001 | ||
| Variety × Water | 0.0987 | ||
| Variety × GB | 0.0188 | ||
| Water × GB | <0.0001 | ||
| Variety × Water × GB | 0.0005 | ||
| Water Stress | Models | R2 | F | p > F |
|---|---|---|---|---|
| Flooding | LDM1 = −0.858 + 0.071XAPX + 0.014XPH | 0.91 | 89.03 | <0.001 |
| Drought | LDM2 = −0.504 − 0.051XPOD + 0.022XPH | 0.80 | 32.35 | <0.001 |
| Varieties | Water Treatments | D Value | Tolerance Coefficient |
|---|---|---|---|
| Heyu 397 | CK | 0.53 | - |
| Flooding | 0.10 | 0.82 | |
| Drought | 0.48 | 1.07 | |
| Yindieyu 9 | CK | 0.59 | - |
| Flooding | 0.40 | 0.93 | |
| Drought | 0.50 | 1.12 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, G.-Y.; Ahmad, S.; Wang, B.-W.; Shi, L.-B.; Wang, Y.; Shi, C.-Q.; Zhou, X.-B. Exogenous Glycinebetaine Regulates the Contrasting Responses in Leaf Physiochemical Attributes and Growth of Maize under Drought and Flooding Stresses. Biology 2024, 13, 360. https://doi.org/10.3390/biology13060360
Wang G-Y, Ahmad S, Wang B-W, Shi L-B, Wang Y, Shi C-Q, Zhou X-B. Exogenous Glycinebetaine Regulates the Contrasting Responses in Leaf Physiochemical Attributes and Growth of Maize under Drought and Flooding Stresses. Biology. 2024; 13(6):360. https://doi.org/10.3390/biology13060360
Chicago/Turabian StyleWang, Guo-Yun, Shakeel Ahmad, Bing-Wei Wang, Li-Bo Shi, Yong Wang, Cheng-Qiao Shi, and Xun-Bo Zhou. 2024. "Exogenous Glycinebetaine Regulates the Contrasting Responses in Leaf Physiochemical Attributes and Growth of Maize under Drought and Flooding Stresses" Biology 13, no. 6: 360. https://doi.org/10.3390/biology13060360
APA StyleWang, G.-Y., Ahmad, S., Wang, B.-W., Shi, L.-B., Wang, Y., Shi, C.-Q., & Zhou, X.-B. (2024). Exogenous Glycinebetaine Regulates the Contrasting Responses in Leaf Physiochemical Attributes and Growth of Maize under Drought and Flooding Stresses. Biology, 13(6), 360. https://doi.org/10.3390/biology13060360