
11-Deoxycorticosterone (DOC)’s Action on the Gill Osmoregulation of Juvenile Rainbow Trout (Oncorhynchus mykiss)

 ,
,  and
and 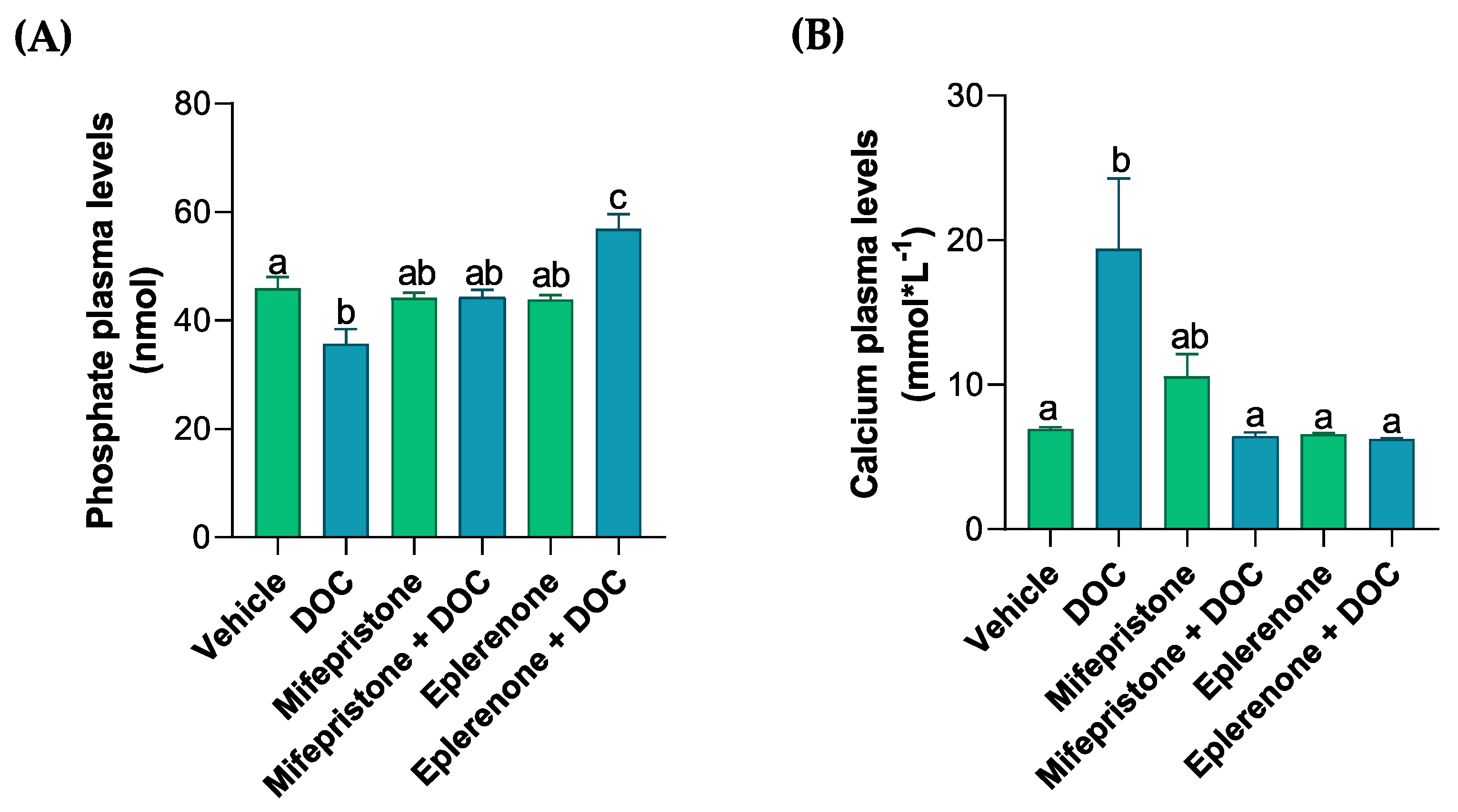{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Protocol
2.2. Measurement of the Solutes in Plasma
2.3. Measurement of the Muscle Water Content
2.4. Library Construction and Sequencing
2.5. Raw Data Processing, RNA-Seq Analysis, and Functional Annotation Analysis
2.6. Real-Time PCR Validation
2.7. Statistical Analysis
3. Results
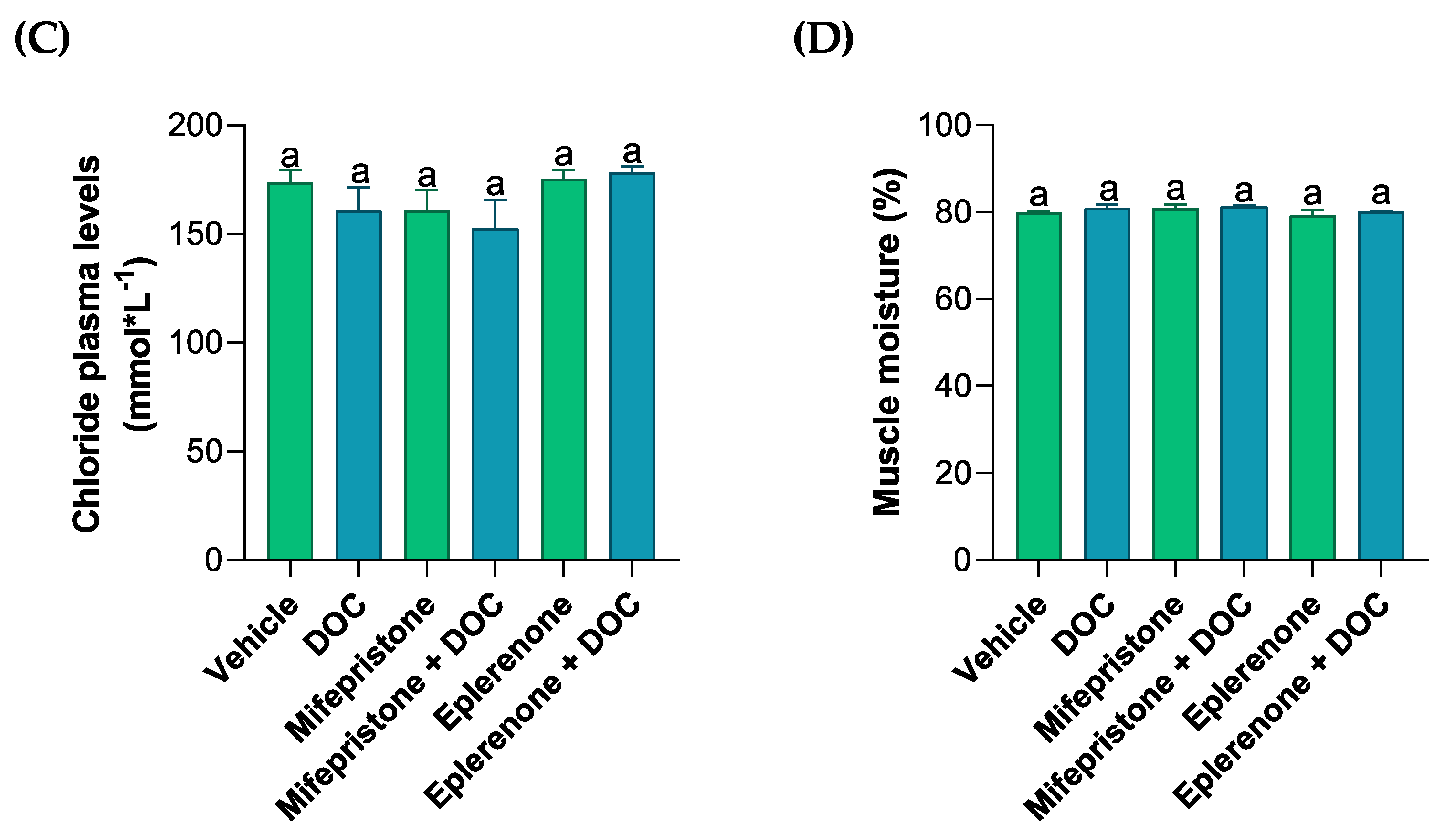
3.1. Evaluation of Solutes in the Plasma, Muscle Water Content, and Glycogen Content in the Gills
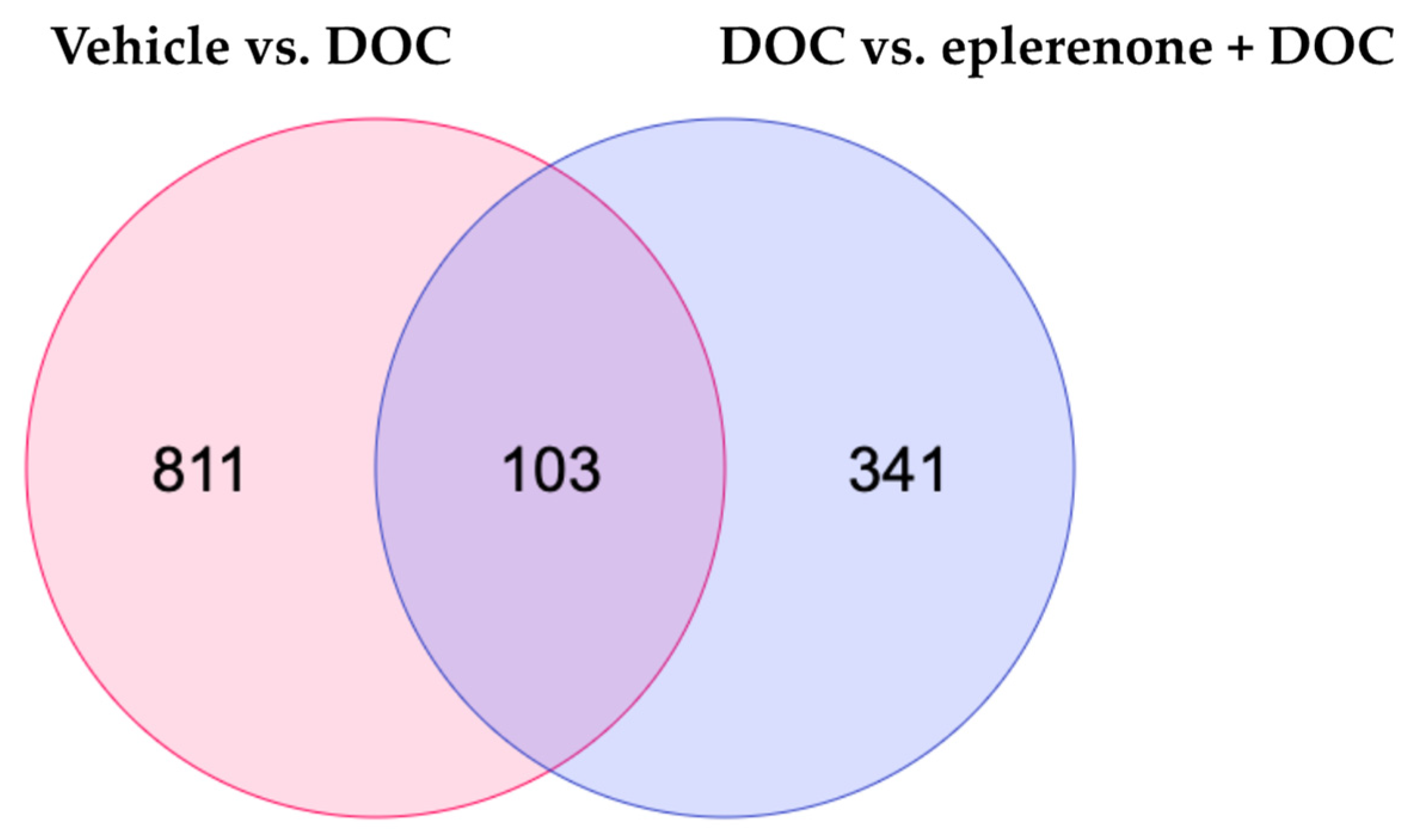
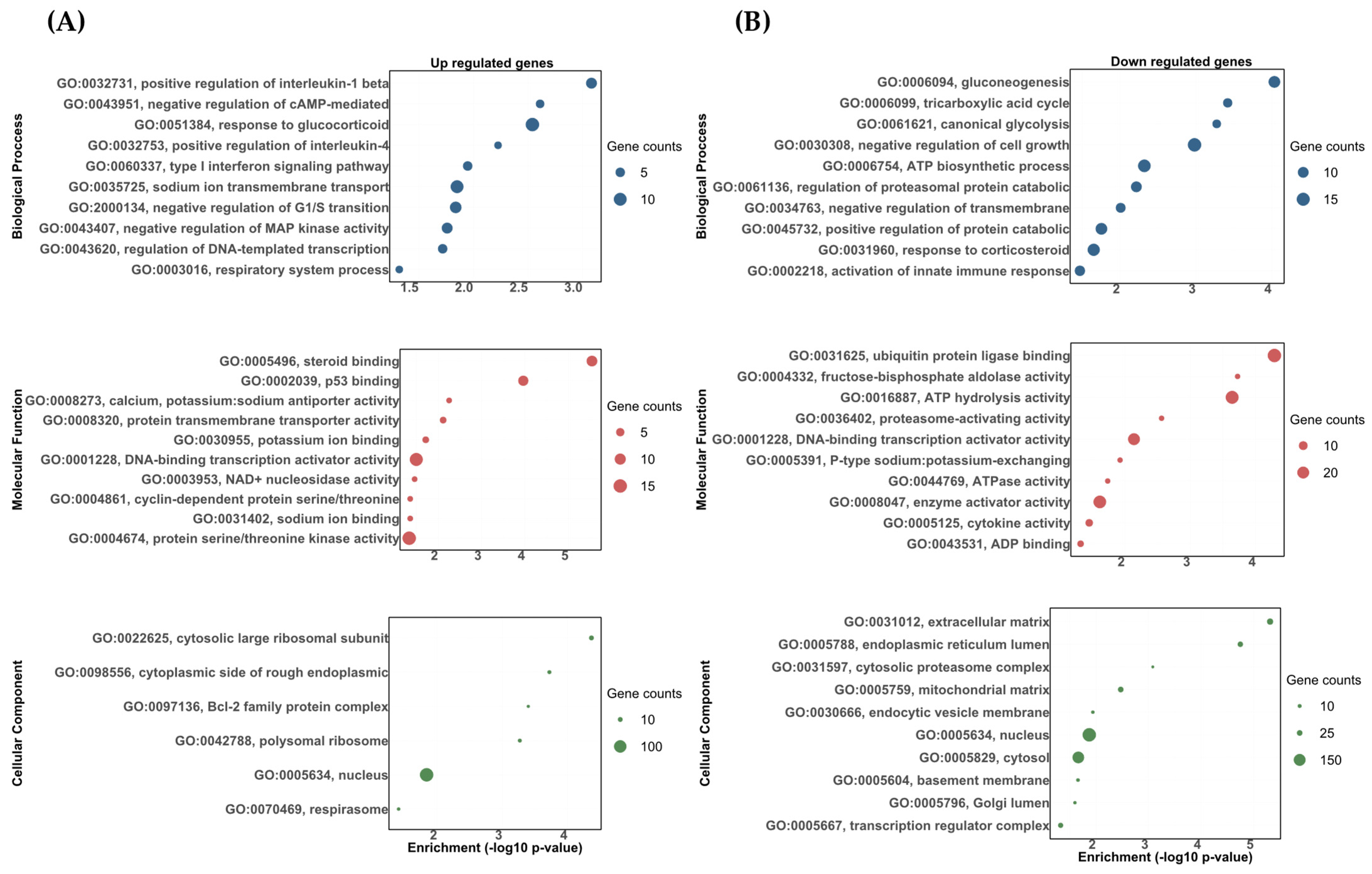
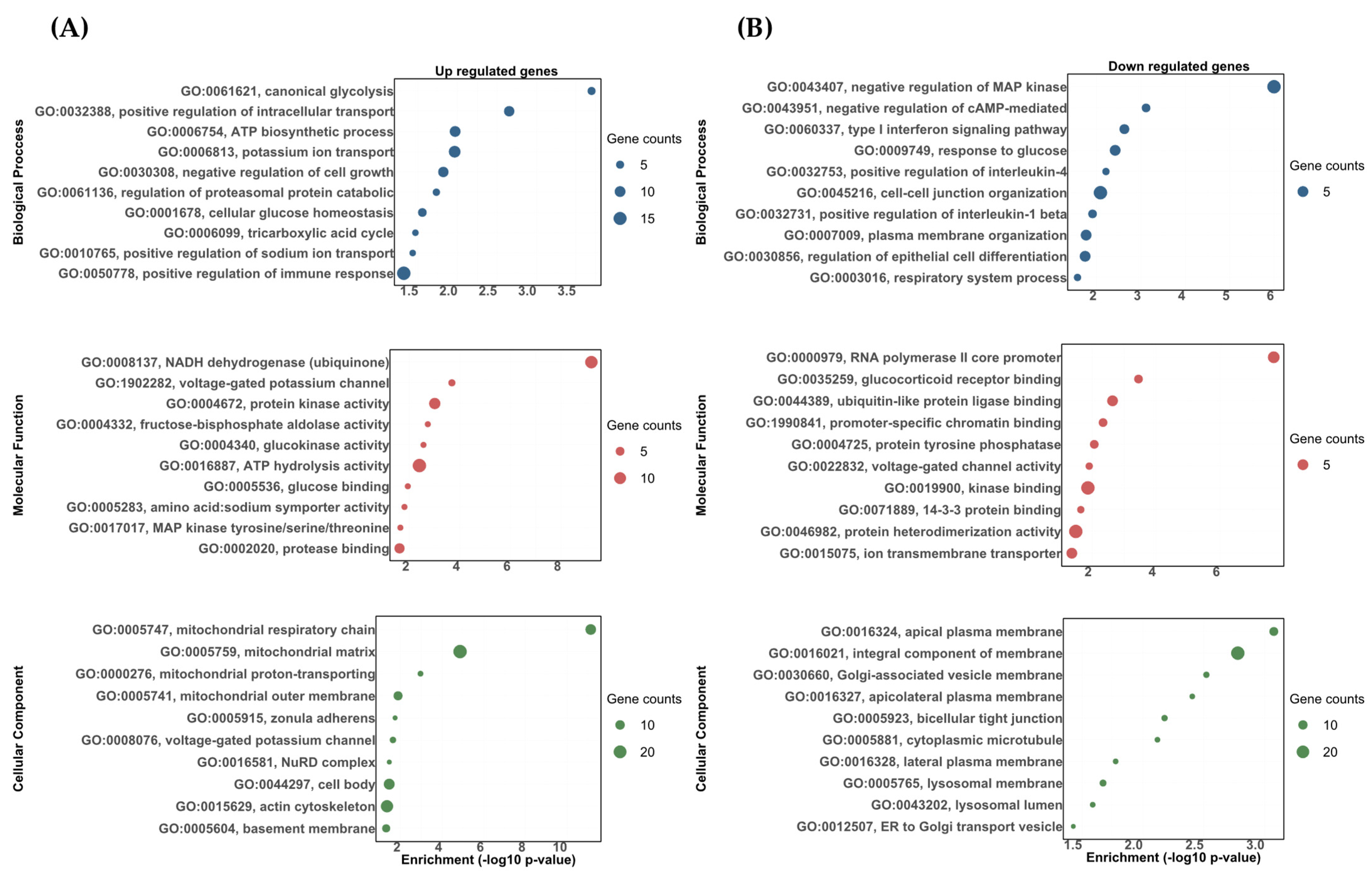
3.2. Transcriptomic Responses of the Rainbow Trout Gill to DOC Mediated by the Mineralocorticoid Receptor (MR)
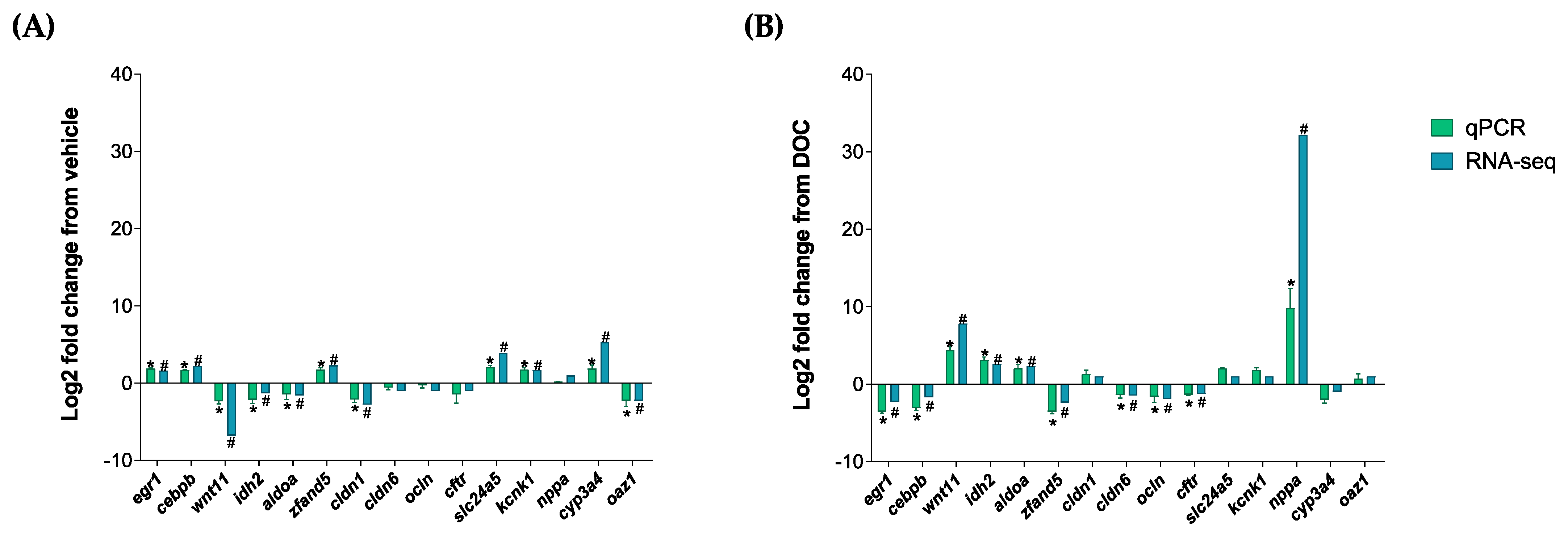
3.3. Validation of the In Silico Data by Real-Time PCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Friedman, K.J.; Bartley, D.M.; Rodríguez-Ezpeleta, N.; Mair, G.C.; Ban, N.; Beveridge, M.; Carolsfeld, J.; Carvalho, G.; Cowx, I.; Dean, G.; et al. Current and Future Genetic Technologies for Fisheries and Aquaculture: Implications for the Work of FAO; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2022; ISBN 9789251367162. [Google Scholar] [CrossRef]
- FAO. The State of World Fisheries and Aquaculture 2022; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2022; ISBN 9789251363645. [Google Scholar] [CrossRef]
- Rehman, S.; Gora, A.H.; Ahmad, I.; Rasool, S.I. Stress in aquaculture hatcheries: Source, impact and mitigation. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 3030–3045. [Google Scholar] [CrossRef]
- Ciji, A.; Akhtar, M.S. Stress management in aquaculture: A review of dietary interventions. Rev. Aquac. 2021, 13, 2190–2247. [Google Scholar] [CrossRef]
- Balasch, J.C.; Tort, L. Netting the Stress Responses in Fish. Front. Endocrinol. 2019, 10, 62. [Google Scholar] [CrossRef]
- Schreck, C.B.; Tort, L. The concept of stress in fish. In Fish Physiology; Academic Press: Cambridge, MA, USA, 2016; Volume 35, pp. 1–34. [Google Scholar]
- Baker, M.E. Evolution of Glucocorticoid and Mineralocorticoid Responses: Go Fish. Endocrinology 2003, 144, 4223–4225. [Google Scholar] [CrossRef][Green Version]
- Baker, M.E.; Katsu, Y. Chapter Two—Evolution of the Mineralocorticoid Receptor. Vitamins and Hormones; Gerald, L., Ed.; Elsevier: Amsterdam, The Netherlands, 2019; Volume 109, pp. 17–36. [Google Scholar] [CrossRef]
- Barton, B.A. Stress in fishes: A diversity of responses with particular reference to changes in circulating corticosteroids. Integr. Comp. Biol. 2002, 42, 517–525. [Google Scholar] [CrossRef]
- Greenwood, A.K.; Butler, P.C.; White, R.B.; DeMarco, U.; Pearce, D.; Fernald, R.D. Multiple corticosteroid receptors in a teleost fish: Distinct sequences, expression patterns, and transcriptional activities. Endocrinology 2003, 144, 4226–4236. [Google Scholar] [CrossRef]
- Arterbery, A.S.; Fergus, D.J.; A Fogarty, E.; Mayberry, J.; Deitcher, D.L.; Kraus, W.L.; Bass, A.H. Evolution of ligand specificity in vertebrate corticosteroid receptors. BMC Evol. Biol. 2011, 11, 14. [Google Scholar] [CrossRef] [PubMed]
- Prunet, P.; Sturm, A.; Milla, S. Multiple corticosteroid receptors in fish: From old ideas to new concepts. Gen. Comp. Endocrinol. 2006, 147, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Sturm, A.; Bury, N.; Dengreville, L.; Fagart, J.; Flouriot, G.; Rafestin-Oblin, M.E.; Prunet, P. 11-deoxycorticosterone is a potent agonist of the rainbow trout (Oncorhynchus mykiss) mineralocorticoid receptor. Endocrinology 2005, 146, 47–55. [Google Scholar] [CrossRef]
- Milla, S.; Terrien, X.; Sturm, A.; Ibrahim, F.; Giton, F.; Fiet, J.; Prunet, P.; Le Gac, F. Plasma 11-Deoxycorticosterone (DOC) and Mineralocorticoid Receptor Testicular Expression during Rainbow Trout Oncorhynchus Mykiss Spermiation: Implication with 17alpha, 20beta-Dihydroxyprogesterone on the Milt Fluidity? Reprod. Biol. Endocrinol. 2008, 6, 19. [Google Scholar] [CrossRef] [PubMed]
- Gilmour, K.M. Mineralocorticoid Receptors and Hormones: Fishing for Answers. Endocrinology 2005, 146, 44–46. [Google Scholar] [CrossRef]
- Milla, S.; Wang, N.; Mandiki, S.; Kestemont, P. Corticosteroids: Friends or Foes of Teleost Fish Reproduction? Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2009, 153, 242–251. [Google Scholar] [CrossRef]
- Kiilerich, P.; Servili, A.; Péron, S.; Valotaire, C.; Goardon, L.; Leguen, I.; Prunet, P. Regulation of the Corticosteroid Signalling System in Rainbow Trout HPI Axis during Confinement Stress. Gen. Comp. Endocrinol. 2018, 258, 184–193. [Google Scholar] [CrossRef] [PubMed]
- Soengas, J.L.; Sangiao-Alvarellos, S.; Laiz-Carrion, R.; Mancera, J.M. Energy metabolism and osmotic acclimation in teleost fish. In Fish Osmoregulation, 1st ed.; CRC Press: Boca Raton, FL, USA, 2007; pp. 277–307. [Google Scholar] [CrossRef]
- Bourque, C.W. Central mechanisms of osmosensation and systemic osmoregulation. Nat. Rev. Neurosci. 2008, 9, 519–531. [Google Scholar] [CrossRef] [PubMed]
- Fridman, S. Ontogeny of the Osmoregulatory Capacity of Teleosts and the Role of Ionocytes. Front. Mar. Sci. 2020, 7, 709. [Google Scholar] [CrossRef]
- Richards, J.G.; Semple, J.W.; Bystriansky, J.S.; Schulte, P.M. Na+/K+-ATPase α-isoform switching in gills of rainbow trout (Oncorhynchus mykiss) during salinity transfer. J. Exp. Biol. 2003, 206, 4475–4486. [Google Scholar] [CrossRef] [PubMed]
- Gamba, G. Molecular Physiology and pathophysiology of electroneutral cation-chloride cotransporters. Physiol. Rev. 2005, 85, 423–493. [Google Scholar] [CrossRef] [PubMed]
- Chasiotis, H.; Kolosov, D.; Bui, P.; Kelly, S.P. Tight junctions, tight junction proteins and paracellular permeability across the gill epithelium of fishes: A review. Respir. Physiol. Neurobiol. 2012, 184, 269–281. [Google Scholar] [CrossRef] [PubMed]
- Trubitt, R.T.; Rabeneck, D.B.; Bujak, J.K.; Bossus, M.C.; Madsen, S.S.; Tipsmark, C.K. Transepithelial resistance and claudin expression in trout RTgill-W1 cell line: Effects of osmoregulatory hormones. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2015, 182, 45–52. [Google Scholar] [CrossRef] [PubMed]
- McCormick, S.D.; Regish, A.; O’dea, M.F.; Shrimpton, J.M. Are we missing a mineralocorticoid in teleost fish? Effects of cortisol, deoxycorticosterone and aldosterone on osmoregulation, gill Na+, K+-ATPase activity and isoform mRNA levels in Atlantic salmon. Gen. Comp. Endocrinol. 2008, 157, 35–40. [Google Scholar] [CrossRef]
- Kiilerich, P.; Pedersen, S.H.; Kristiansen, K.; Madsen, S.S. Corticosteroid regulation of Na+, K+-ATPase α1-isoform expression in Atlantic salmon gill during smolt development. Gen. Comp. Endocrinol. 2011, 170, 283–289. [Google Scholar] [CrossRef]
- Zuloaga, R.; Aravena-Canales, D.; Aedo, J.E.; Osorio-Fuentealba, C.; Molina, A.; Valdés, J.A. Effect of 11-Deoxycorticosterone in the Transcriptomic Response to Stress in Rainbow Trout Skeletal Muscle. Genes 2023, 14, 512. [Google Scholar] [CrossRef]
- Villarreal, F.D.; Kültz, D. Direct ionic regulation of the activity of myo-inositol biosynthesis enzymes in Mozambique tilapia. PLoS ONE 2015, 10, e0123212. [Google Scholar] [CrossRef]
- Srivastav, S.; Mishra, D.; Srivastav, S.K.; Suzuki, N.; Srivastav, A.K. Estradiol Affects Prolactin Producing Cells and Calcium levels in a Teleost, Heteropneustesfossilis, Kept in Different Calcium Concentrations. Iran. J. Toxicol. 2017, 11, 45–51. [Google Scholar] [CrossRef]
- Tavares-Dias, M.; Moraes, F.R.D.; Imoto, M.E. Hematological parameters in two neotropical freshwater teleost, Leporinus macrocephalus (Anostomidae) and Prochilodus lineatus (Prochilodontidae). Biosci. J. 2008, 24, 96–101. [Google Scholar]
- Al-Jandal, N.J.; Wilson, R.W. A comparison of osmoregulatory responses in plasma and tissues of rainbow trout (Oncorhynchus mykiss) following acute salinity challenges. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2011, 159, 175–181. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for clustering the next-generation sequencing data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Danecek, P.; Bonfield, J.K.; Liddle, J.; Marshall, J.; Ohan, V.; Pollard, M.O.; Whitwham, A.; Keane, T.; McCarthy, S.A.; Davies, R.M.; et al. Twelve years of SAMtools and BCFtools. GigaScience 2021, 10, giab008. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Smyth, G.K.; Shi, W. The R package Rsubread is easier, faster, cheaper and better for alignment and quantification of RNA sequencing reads. Nucleic Acids Res. 2019, 47, e47. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Cantalapiedra, C.P.; Hernández-Plaza, A.; Letunic, I.; Bork, P.; Huerta-Cepas, J. eggNOG-mapper v2: Functional annotation, orthology assignments, and domain prediction at the metagenomic scale. Mol. Biol. Evol. 2021, 38, 5825–5829. [Google Scholar] [CrossRef] [PubMed]
- Alexa, A.; Rahnenführer, J. Gene set enrichment analysis with topGO. Bioconductor Improv. 2009, 27, 1–26. [Google Scholar]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis, 2nd ed.; Springer International Publishing: Cham, Switzerland, 2016; p. 260, (Use R!). [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Baker, M.E.; Funder, J.W.; Kattoula, S.R. Evolution of hormone selectivity in glucocorticoid and mineralocorticoid receptors. J. Steroid Biochem. Mol. Biol. 2013, 137, 57–70. [Google Scholar] [CrossRef] [PubMed]
- Viengchareun, S.; Le Menuet, D.; Martinerie, L.; Munier, M.; Tallec, L.P.-L.; Lombès, M. The Mineralocorticoid Receptor: Insights into its Molecular and (Patho)Physiological Biology. Nucl. Recept. Signal. 2007, 5, e012. [Google Scholar] [CrossRef] [PubMed]
- Gorissen, M.; Flik, G. The endocrinology of the stress response in fish: An adaptation-physiological view. In Fish Physiology; Carl, B., Schreck, L.T., Anthony, P., Farrell, C., Brauner, J., Eds.; Academic Press: Cambridge, MA, USA, 2016; Volume 35, pp. 75–111. [Google Scholar] [CrossRef]
- Lin, C.-H.; Tsai, I.-L.; Su, C.-H.; Tseng, D.-Y.; Hwang, P.-P. Reverse Effect of Mammalian Hypocalcemic Cortisol in Fish: Cortisol Stimulates Ca2+ Uptake via Glucocorticoid Receptor-Mediated Vitamin D3 Metabolism. PLoS ONE 2011, 6, e23689. [Google Scholar] [CrossRef] [PubMed]
- Das, C.; Rout, M.K.; Wildering, W.C.; Vijayan, M.M. Cortisol modulates calcium release-activated calcium channel gating in fish hepatocytes. Sci. Rep. 2021, 11, 9621. [Google Scholar] [CrossRef]
- Michea, L.; Delpiano, A.M.; Hitschfeld, C.; Lobos, L.; Lavandero, S.; Marusic, E.T. Eplerenone blocks nongenomic effects of aldosterone on the Na+/H+ exchanger, intracellular Ca2+ levels, and vasoconstriction in mesenteric resistance vessels. Endocrinology 2005, 146, 973–980. [Google Scholar] [CrossRef]
- Abbink, W.; Bevelander, G.S.; Rotllant, J.; Canario, A.V.M.; Flik, G. Calcium handling in Sparus auratus: Effects of water and dietary calcium levels on mineral composition, cortisol and PTHrP levels. J. Exp. Biol. 2004, 207, 4077–4084. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Suarez-Bregua, P.; Guerreiro, P.M.; Rotllant, J. Stress, Glucocorticoids and Bone: A Review from Mammals and Fish. Front. Endocrinol. 2018, 9, 526. [Google Scholar] [CrossRef] [PubMed]
- Sbaihi, M.; Rousseau, K.; Baloche, S.; Meunier, F.; Fouchereau-Peron, M.; Dufour, S. Cortisol mobilizes mineral stores from vertebral skeleton in the European eel: An ancestral origin for glucocorticoid-induced osteoporosis? J. Endocrinol. 2009, 201, 241–252. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sakamoto, T.; Kozaka, T.; Takahashi, A.; Kawauchi, H.; Ando, M. Medaka (Oryzias latipes) as a model for hypoosmoregulation of euryhaline fishes. Aquaculture 2001, 193, 347–354. [Google Scholar] [CrossRef]
- McCormick, S.D.; Taylor, M.L.; Regish, A.M. Cortisol is an osmoregulatory and glucose-regulating hormone in Atlantic sturgeon, a basal ray-finned fish. J. Exp. Biol. 2020, 223, jeb220251. [Google Scholar] [CrossRef] [PubMed]
- Marshall, W.; Cozzi, R.; Pelis, R.; Mccormick, S. Cortisol receptor blockade and seawater adaptation in the euryhaline teleostFundulus heteroclitus. J. Exp. Zool. A Comp. Exp. Biol. 2005, 303A, 132–142. [Google Scholar] [CrossRef]
- Jerez-Cepa, I.; Gorissen, M.; Mancera, J.; Ruiz-Jarabo, I. What can we learn from glucocorticoid administration in fish? Effects of cortisol and dexamethasone on intermediary metabolism of gilthead seabream (Sparus aurata L.). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2019, 231, 1–10. [Google Scholar] [CrossRef]
- Chasiotis, H.; Kelly, S.P. Effects of elevated circulating cortisol levels on hydromineral status and gill tight junction protein abundance in the stenohaline goldfish. Gen. Comp. Endocrinol. 2012, 175, 277–283. [Google Scholar] [CrossRef]
- Kiilerich, P.; Milla, S.; Sturm, A.; Valotaire, C.; Chevolleau, S.; Giton, F.; Terrien, X.; Fiet, J.; Fostier, A.; Debrauwer, L.; et al. Implication of the mineralocorticoid axis in rainbow trout osmoregulation during salinity acclimation. J. Endocrinol. 2011, 209, 221–235. [Google Scholar] [CrossRef]
- Takahashi, H.; Sakamoto, T. The role of ‘mineralocorticoids’ in teleost fish: Relative importance of glucocorticoid signaling in the osmoregulation and ‘central’ actions of mineralocorticoid receptor. Gen. Comp. Endocrinol. 2013, 181, 223–228. [Google Scholar] [CrossRef]
- Zhang, J.; Xie, S.; Ma, W.; Teng, Y.; Tian, Y.; Huang, X.; Zhang, Y. A newly identified microRNA, mmu-miR-7578, functions as a negative regulator on inflammatory cytokines tumor necrosis factor-α and interleukin-6 via targeting Egr1 in vivo. J. Biol. Chem. 2013, 288, 4310–4320. [Google Scholar] [CrossRef]
- Cheng, C.; Liu, H.; Tan, C.; Tong, D.; Zhao, Y.; Liu, X.; Si, W.; Wang, L.; Liang, L.; Li, J.; et al. Mutation in NPPA causes atrial fibrillation by activating inflammation and cardiac fibrosis in a knock-in rat model. FASEB J. 2019, 33, 8878–8891. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, C.; Milla, S.; Mandiki, S.; Douxfils, J.; Douny, C.; Scippo, M.-L.; De Pauw, E.; Kestemont, P. First evidence of the possible implication of the 11-deoxycorticosterone (DOC) in immune activity of Eurasian perch (Perca fluviatilis, L.): Comparison with cortisol. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2013, 165, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, C.; Milla, S.; Mandiki, S.; Douxfils, J.; Kestemont, P. In vivo response of some immune and endocrine variables to LPS in Eurasian perch (Perca fluviatilis, L.) and modulation of this response by two corticosteroids, cortisol and 11-deoxycorticosterone. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2014, 167, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Naderi, M.; Keyvanshokooh, S.; Ghaedi, A.; Salati, A.P. Effect of acute crowding stress on rainbow trout (Oncorhynchus mykiss): A proteomics study. Aquaculture 2018, 495, 106–114. [Google Scholar] [CrossRef]
- Mu, Y.; Li, W.; Wei, Z.; He, L.; Zhang, W.; Chen, X. Transcriptome analysis reveals molecular strategies in gills and heart of large yellow croaker (Larimichthys crocea) under hypoxia stress. Fish Shellfish. Immunol. 2020, 104, 304–313. [Google Scholar] [CrossRef] [PubMed]
- Milla, S.; Massart, S.; Mathieu, C.; Wang, N.; Douny, C.; Douxfils, J.; Scippo, M.-L.; De Pauw, E.; Dieu, M.; Silvestre, F.; et al. Physiological and proteomic responses to corticosteroid treatments in Eurasian perch, Perca fluviatilis: Investigation of immune-related parameters. Comp. Biochem. Physiol. Part D Genom. Proteom. 2018, 25, 86–98. [Google Scholar] [CrossRef] [PubMed]
- Tseng, Y.-C.; Hwang, P.-P. Some insights into energy metabolism for osmoregulation in fish. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2008, 148, 419–429. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.-Y.; Lee, T.-H.; Tseng, D.-Y. Glucocorticoid Receptor Mediates Cortisol Regulation of Glycogen Metabolism in Gills of the Euryhaline Tilapia (Oreochromis mossambicus). Fishes 2023, 8, 267. [Google Scholar] [CrossRef]
- Faught, E.; Vijayan, M.M. The mineralocorticoid receptor functions as a key glucose regulator in the skeletal muscle of zebrafish. Endocrinology 2022, 163, bqac149. [Google Scholar] [CrossRef]
- Wu, C.-Y.; Lee, T.-H.; Tseng, D.-Y. Mineralocorticoid Receptor Mediates Cortisol Regulation of Ionocyte Development in Tilapia (Oreochromis mossambicus). Fishes 2023, 8, 283. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zuloaga, R.; Ahumada-Langer, L.; Aedo, J.E.; Molina, A.; Valdés, J.A. 11-Deoxycorticosterone (DOC)’s Action on the Gill Osmoregulation of Juvenile Rainbow Trout (Oncorhynchus mykiss). Biology 2024, 13, 107. https://doi.org/10.3390/biology13020107
Zuloaga R, Ahumada-Langer L, Aedo JE, Molina A, Valdés JA. 11-Deoxycorticosterone (DOC)’s Action on the Gill Osmoregulation of Juvenile Rainbow Trout (Oncorhynchus mykiss). Biology. 2024; 13(2):107. https://doi.org/10.3390/biology13020107
Chicago/Turabian StyleZuloaga, Rodrigo, Luciano Ahumada-Langer, Jorge Eduardo Aedo, Alfredo Molina, and Juan Antonio Valdés. 2024. "11-Deoxycorticosterone (DOC)’s Action on the Gill Osmoregulation of Juvenile Rainbow Trout (Oncorhynchus mykiss)" Biology 13, no. 2: 107. https://doi.org/10.3390/biology13020107
APA StyleZuloaga, R., Ahumada-Langer, L., Aedo, J. E., Molina, A., & Valdés, J. A. (2024). 11-Deoxycorticosterone (DOC)’s Action on the Gill Osmoregulation of Juvenile Rainbow Trout (Oncorhynchus mykiss). Biology, 13(2), 107. https://doi.org/10.3390/biology13020107