

Macro, Trace and Toxic Element Composition in Liver and Meat of Broiler Chicken Associated with Cecal Microbiome Community

 , ,
, ,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Data Collection: Animals and Feed
2.3. Determination of Elements: Samples Preparation and Analytical Procedure
2.4. Microbial Community: DNA Extraction, Sequencing, and Data Processing
2.5. Statistical Analysis
3. Results
3.1. Macro, Trace and Toxic Element Content in the Liver and Meat of Broiler Chickens
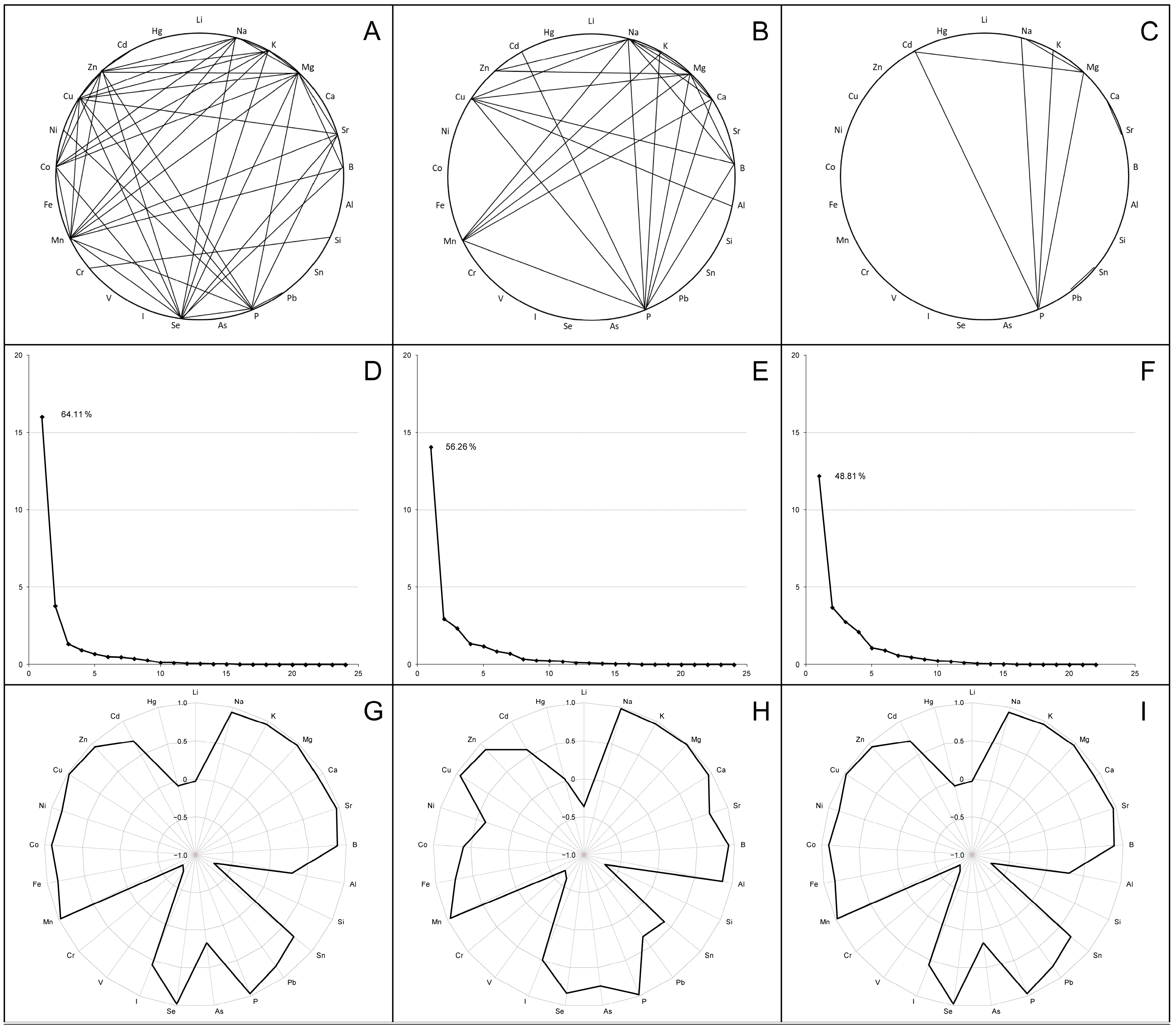
3.2. Elemental Compositions of Broiler Chicken Biosamples
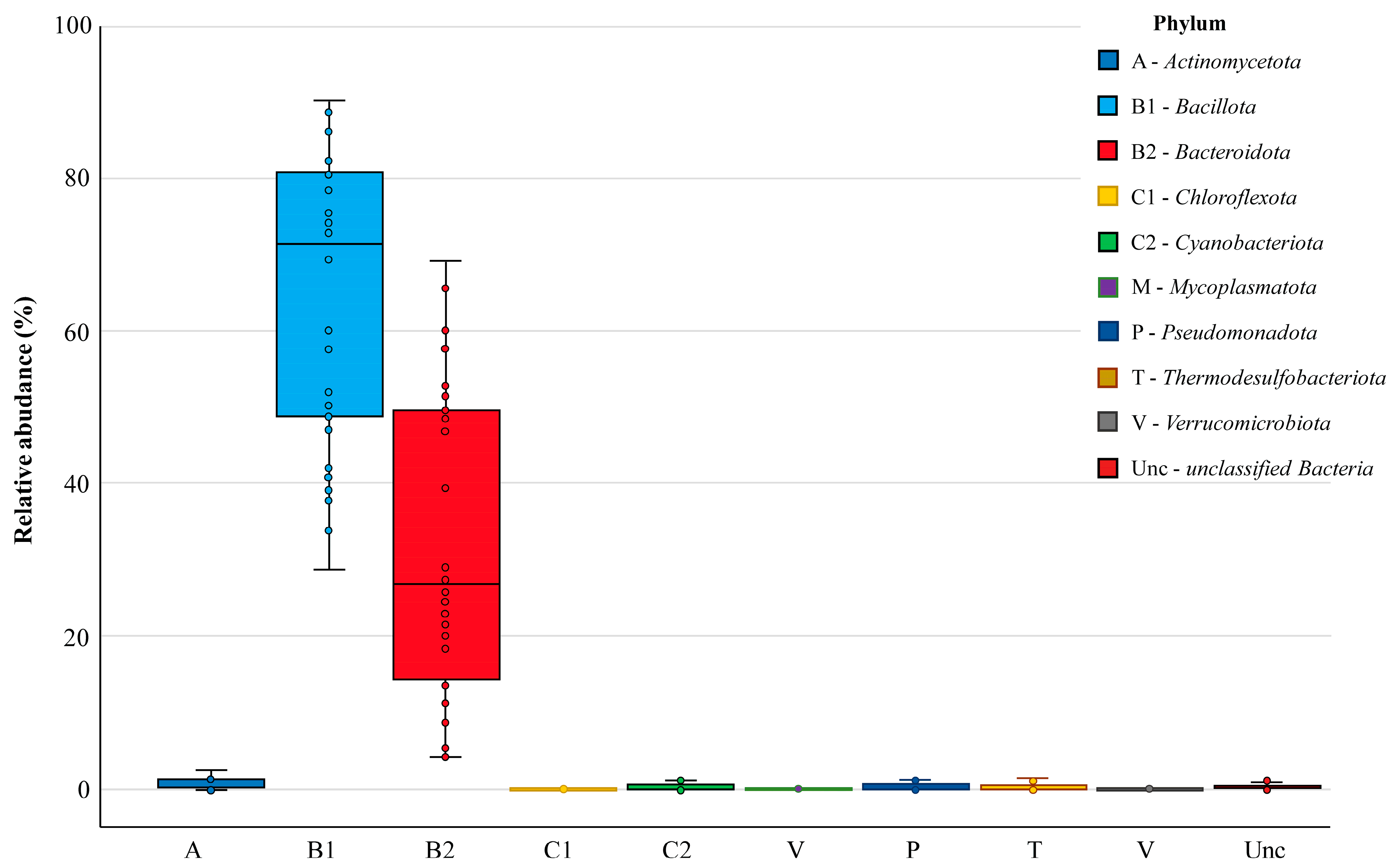
3.3. Bacterial Community in Chicken Cecal Microbiomes
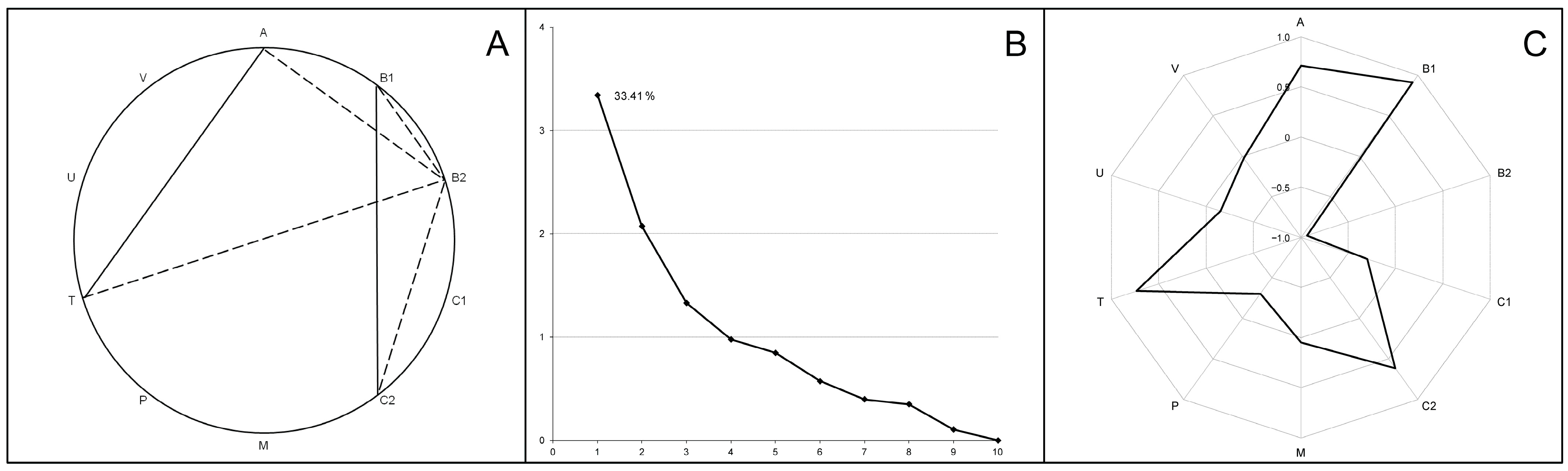
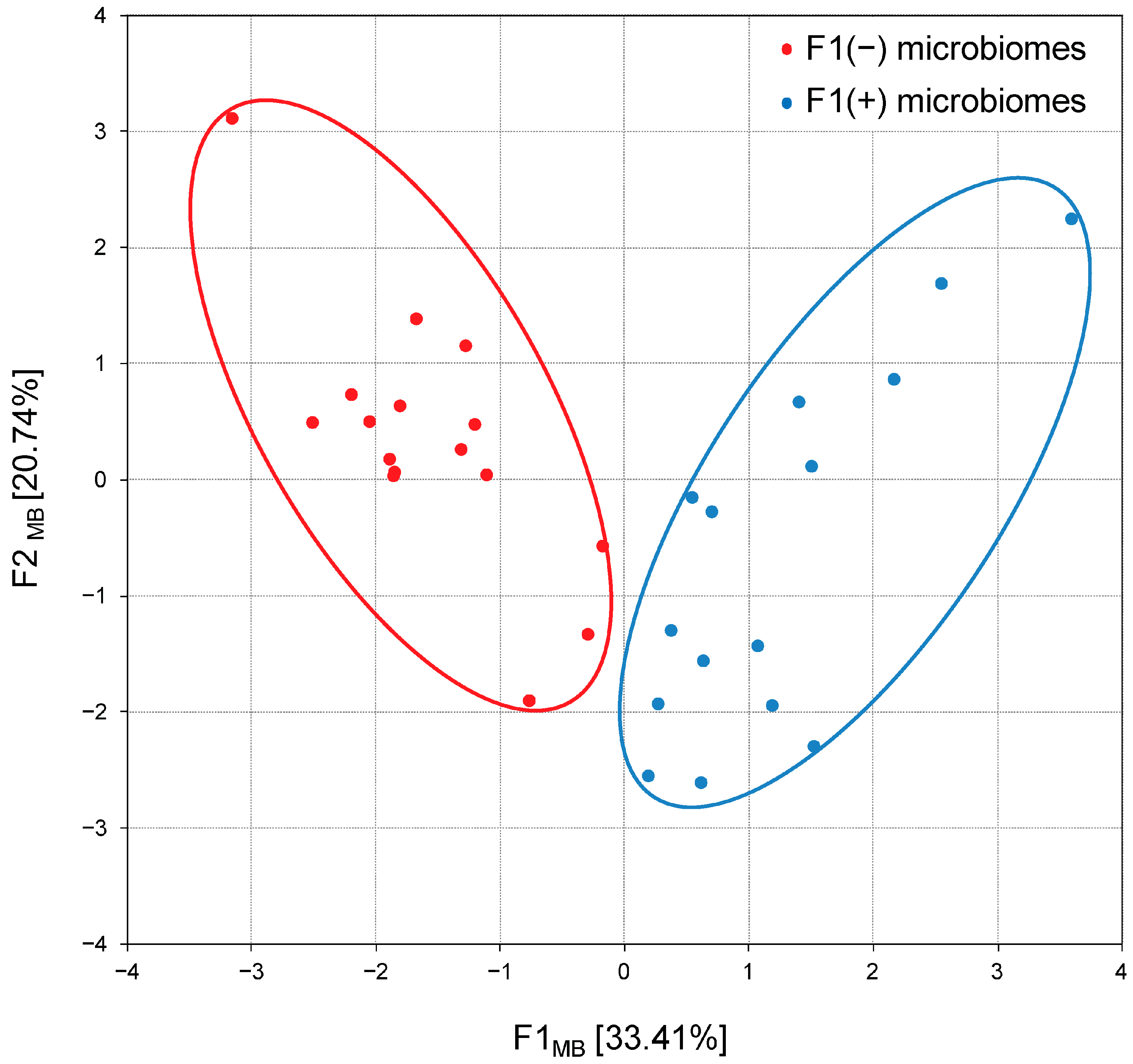
3.4. Cecal Microbiome Communities (Microbiome Patterns)
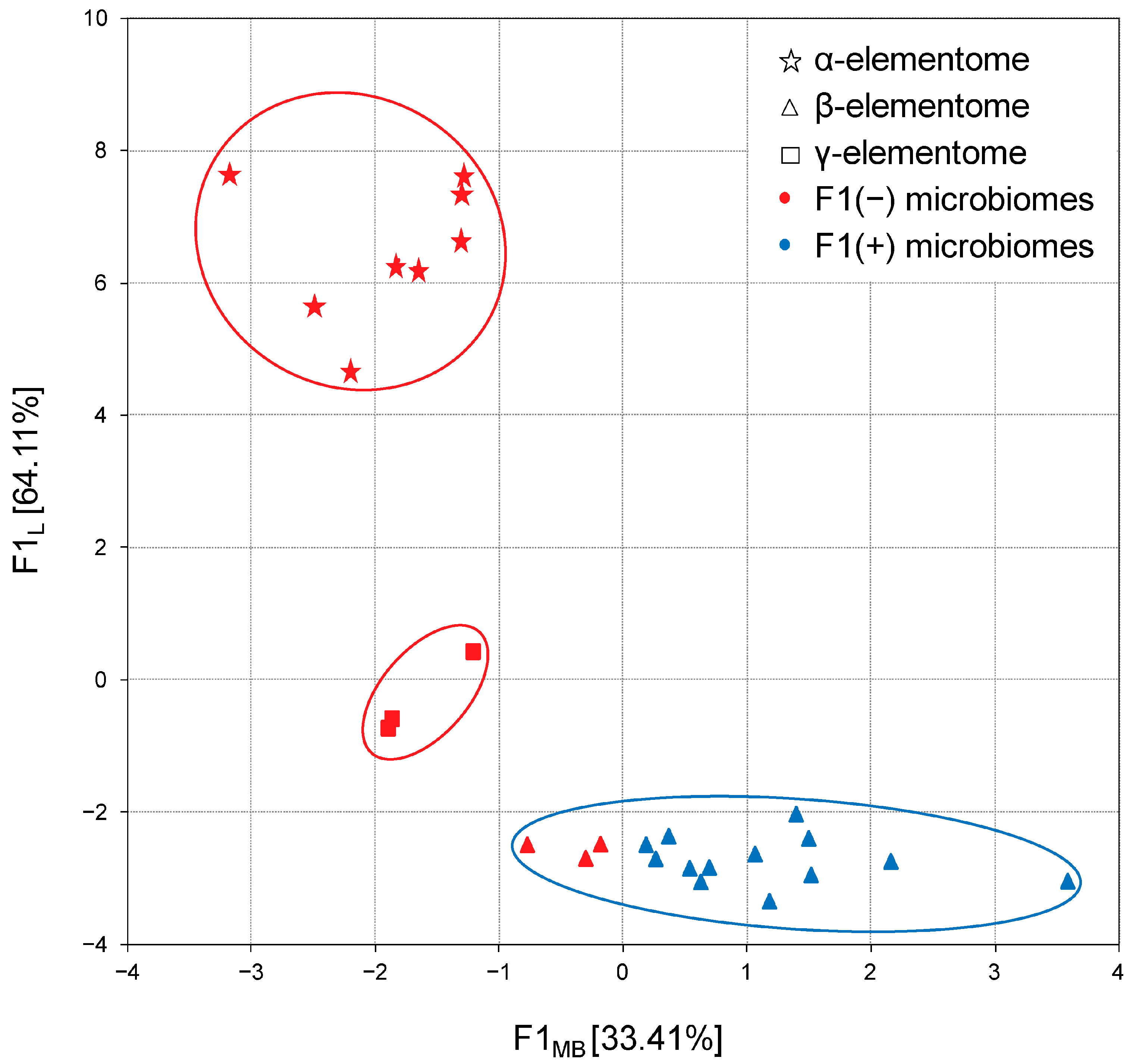
3.5. Microbiome–Elementome Associations
3.6. Bacterial Taxa Influenced the Elemental Compositions in Broiler Chicken Biosamples
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wahyono, N.D.; Utami, M.M.D. A Review of the Poultry Meat Production Industry for Food Safety in Indonesia. J. Phys. Conf. Ser. 2018, 953, 012125. [Google Scholar] [CrossRef]
- Santos, A.N.A.; Viana, A.L.; Guimaraes, C.C.; Gomes, M.F.S.; Barai, A.A.; Nobrega, T.C.; Ribeiro, M.W.S.; Silva, A.J.I.; Chaves, F.A.L.; Mendonca, M.A.F.; et al. Paullinia cupana peel meal on the growth performance, meat quality, and haematological and serum biochemical parameters of slow-growing broilers. Anim. Prod. Sci. 2024, 64, AN24137. [Google Scholar] [CrossRef]
- Statista. Available online: https://www.statista.com/statistics/237644/global-meat-production-since-1990/ (accessed on 24 November 2024).
- Korish, M.A.; Attia, Y.A. Evaluation of heavy metal content in feed, litter, meat, meat products, liver, and table eggs of chickens. Animals 2020, 10, 727. [Google Scholar] [CrossRef] [PubMed]
- Aljohani, A.S.M. Heavy metal toxicity in poultry: A comprehensive review. Front. Vet. Sci. 2023, 10, 1161354. [Google Scholar] [CrossRef] [PubMed]
- Nisianakis, P.; Giannenas, I.; Gavriil, A.; Kontopidis, G.; Kyriazakis, I. Variation in trace element contents among chicken, Turkey, duck, goose, and pigeon eggs analyzed by inductively coupled plasma mass spectrometry (ICP-MS). Biol. Trace Element Res. 2008, 128, 62–71. [Google Scholar] [CrossRef]
- Chen, S.-S.; Lin, Y.-W.; Kao, Y.-M.; Shih, Y.-C. Trace elements and heavy metals in poultry and livestock meat in Taiwan. Food Addit. Contam. Part B 2013, 6, 231–236. [Google Scholar] [CrossRef]
- Kamaly, H.F.; Sharkawy, A.A. Health risk assessment of metals in chicken meat and liver in Egypt. Environ. Monit. Assess. 2023, 195, 802. [Google Scholar] [CrossRef]
- Yang, L.; He, T.; Xiong, F.; Chen, X.; Fan, X.; Jin, S.; Geng, Z. Identification of key genes and pathways associated with feed efficiency of native chickens based on transcriptome data via bioinformatics analysis. BMC Genom. 2020, 21, 292. [Google Scholar] [CrossRef]
- Mondal, S.; Haldar, S.; Saha, P.; Ghosh, T.K. Metabolism and tissue distribution of trace elements in broiler chickens’ fed diets containing deficient and plethoric levels of copper, manganese, and zinc. Biol. Trace Elem. Res. 2009, 137, 190–205. [Google Scholar] [CrossRef]
- Lebedev, S.; Kazakova, T.; Marshinskaia, O.; Grechkina, V. The assessment of serum trace element levels as the diagnostic biomarkers of functional state of broiler chickens. Vet. World 2023, 16, 1512–1519. [Google Scholar] [CrossRef]
- Lebedev, S.; Nikitin, A.Y.; Fisinin, V.I.; Egorov, I.A.; Miroshnikov, I.S.; Ryazanov, V.A.; Grechkina, V.V.; Kvan, O.V. Formation of element status at chickens when using enzyme, probiotic and antibiotic agents in food. IOP Conf. Ser. Earth Environ. Sci. 2019, 341, 012077. [Google Scholar] [CrossRef]
- Collins, J.F.; Flores, S.R.L.; Wang, X.; Anderson, G.J. Mechanisms and Regulation of Intestinal Iron Transport. In Physiology of the Gastrointestinal Tract, 6th ed.; Academic Press: Cambridge, MA, USA, 2018; pp. 1451–1483. [Google Scholar] [CrossRef]
- Gommers, L.M.M.; Hoenderop, J.G.J.; de Baaij, J.H.F. Mechanisms of proton pump inhibitor-induced hypomagnesemia. Acta Physiol. 2022, 235, e13846. [Google Scholar] [CrossRef] [PubMed]
- Kasaikina, M.V.; Kravtsova, M.A.; Lee, B.C.; Seravalli, J.; Peterson, D.A.; Walter, J.; Legge, R.; Benson, A.K.; Hatfield, D.L.; Gladyshev, V.N. Dietary selenium affects host selenoproteome expression by influencing the gut microbiota. FASEB J. 2011, 25, 2492–2499. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, F.; Li, H.; Yang, S.; Chen, X.; Long, S.; Yang, S.; Yang, Y.; Wang, Z. Metabolic and inflammatory linkage of the chicken cecal microbiome to growth performance. Front. Microbiol. 2023, 14, 1060458. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Yang, Y.; Long, X.; Li, H.; Zhang, F.; Wang, Z. Integrated Analysis of the Effects of Cecal Microbiota and Serum Metabolome on Market Weights of Chinese Native Chickens. Animals 2023, 13, 3034. [Google Scholar] [CrossRef]
- Qu, A.; Brulc, J.M.; Wilson, M.K.; Law, B.F.; Theoret, J.R.; Joens, L.A.; Konkel, M.E.; Angly, F.; Dinsdale, E.A.; Edwards, R.A.; et al. Comparative metagenomics reveals host specific metavirulomes and horizontal gene transfer elements in the chicken cecum microbiome. PLoS ONE 2008, 3, e2945. [Google Scholar] [CrossRef]
- Pajarillo, E.A.B.; Lee, E.; Kang, D.-K. Trace metals and animal health: Interplay of the gut microbiota with iron, manganese, zinc, and copper. Anim. Nutr. 2021, 7, 750–761. [Google Scholar] [CrossRef]
- Arumugam, M.; Raes, J.; Pelletier, E.; Le Paslier, D.; Yamada, T.; Mende, D.R.; Fernandes, G.R.; Tap, J.; Bruls, T.; Batto, J.M.; et al. Enterotypes of the human gut microbiome. Nature 2011, 473, 174–180. [Google Scholar] [CrossRef]
- Deryabin, D.; Lazebnik, C.; Vlasenko, L.; Karimov, I.; Kosyan, D.; Zatevalov, A.; Duskaev, G. Broiler Chicken Cecal Microbiome and Poultry Farming Productivity: A Meta-Analysis. Microorganisms 2024, 12, 747. [Google Scholar] [CrossRef]
- Yao, X.; Zuo, N.; Guan, W.; Fu, L.; Jiang, S.; Jiao, J.; Wang, X. Association of Gut Microbiota Enterotypes with Blood Trace Elements in Women with Infertility. Nutrients 2022, 14, 3195. [Google Scholar] [CrossRef]
- Rakhmatullin, S.G.; Duskaev, G.K.; Galiev, B.H. Plant extract and probiotcs change elemental status of muscle tissue of broilers. IOP Conf. Series Earth Environ. Sci. 2020, 548, 082015. [Google Scholar] [CrossRef]
- Duskaev, G.; Rakhmatullin, S.; Kvan, O. Effects of Bacillus cereus and coumarin on growth performance, blood biochemical parameters, and meat quality in broilers. Veter. World 2020, 13, 2484–2492. [Google Scholar] [CrossRef] [PubMed]
- Zavyalov, O.; Galimzhan, D.; Marina, K. Effect of feeding bioactive compounds identified from plant extracts (4-hexylresorcinol, 7-hydroxycoumarin, and gamma-octalactone) on the productivity and quality of broiler meat. Veter. World 2022, 15, 2986–2996. [Google Scholar] [CrossRef] [PubMed]
- Duskaev, G.K.; Vlasenko, L.V.; Kosyan, D.B.; Kurilkina, M.Y. The effect of small molecules of plant origin on the cecal microbial diversity in broilers. Ptitsevodstvo 2023, 4, 46–51. [Google Scholar] [CrossRef]
- Duskaev, G.; Kvan, O.; Kosyan, D.; Rakhmatullin, S.; Levakhin, G. Coumarin derivative and Bacillus cereus change live weight and cecal ecology in broilers. AIMS Agric. Food 2021, 6, 360–380. [Google Scholar] [CrossRef]
- Shajahmetova, E.; Deryabin, D.; Duskaev, G. Gamma Lactone and vanillic acid change the microbiome of broiler blind appendage. Biotechnol. Biotechnol. Equip. 2021, 35, S62–S128. [Google Scholar] [CrossRef]
- Duskaev, G.K.; Lazebnik, K.S.; Klimova, T.A. Microbial diversity in the cecum of broiler chickens after introduction of coumarin and feed antibiotic into the diet. RUDN J. Agron. Anim. Ind. 2022, 17, 555–566. [Google Scholar] [CrossRef]
- Duskaev, G.K.; Rakhmatullin, S.G.; Kosyan, D.B.; Rusakova, E.A.; Kvan, O.V.; Levakhin, G.I. The effect of the combined use of gammaoctalactone and chlortetracycline in the broiler diet: Live weight, feed efficiency and the microbiome of the caecum. Agrar. Sci. 2022, 9, 47–53. [Google Scholar] [CrossRef]
- Duskaev, G.K.; Rusakova, E.A.; Deryabin, D.G. The using of gamma-octanolactone in the broiler diet: Intestinal microbiome. Int. J. Veter. Med. 2022, 4, 152–158. [Google Scholar] [CrossRef]
- Duskaev, G.K.; Klimova, T.A. Influence of coumarin on productivity, immunity and antioxidant status of healthy broilers. Sib. J. Life Sci. Agric. 2023, 15, 197–218. [Google Scholar] [CrossRef]
- Deryabin, D.G.; Kosyan, D.B.; Inchagova, K.S.; Duskaev, G.K. Plant-Derived Quorum Sensing Inhibitors (Quercetin, Vanillin and Umbelliferon) Modulate Cecal Microbiome, Reduces Inflammation and Affect Production Efficiency in Broiler Chickens. Microorganisms 2023, 11, 1326. [Google Scholar] [CrossRef] [PubMed]
- Bagirov, V.A.; Ushakov, A.S.; Duskaev, G.K.; Kvan, O.V.; Rakhmatullin, S.G.; Yausheva, E.V.; Vershinina, I.A. Metagenomic analysis of intestinal microbiome and biochemical composition of broiler meat upon use of quercus cortex extract dietary additive. Sel’skokhozyaistvennaya Biol. 2020, 55, 682–696. [Google Scholar] [CrossRef]
- Ryazanov, V.; Duskaev, G.; Rakhmatullin, S.; Nurzhanov, B.; Rysaev, A. Effects of joint and separate uses of the Quercus cortex extract and antibiotics on the microbiome of the broiler intestine. FEBS Open Bio 2021, 11, 385–386. [Google Scholar] [CrossRef]
- Lebedev, S.V.; Zav’yalov, O.A.; Frolov, A.N.; Kurilkina, M.Y.; Grechkina, V.V. Age-dependent accumulation of essential and toxic chemical elements in feather of arbor acres broilers (Gallus gallus L.) reared in the South Ural biogeochemical province of Russia. Sel’skokhozyaistvennaya Biol. 2023, 58, 386–398. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef]
- Schoch, C.L.; Ciufo, S.; Domrachev, M.; Hotton, C.L.; Kannan, S.; Khovanskaya, R.; Leipe, D.; Mcveigh, R.; O’neill, K.; Robbertse, B.; et al. NCBI Taxonomy: A comprehensive update on curation, resources and tools. Database 2020, 2020, baaa062. [Google Scholar] [CrossRef]
- Peñuelas, J.; Fernández-Martínez, M.; Ciais, P.; Jou, D.; Piao, S.; Obersteiner, M.; Vicca, S.; Janssens, I.A.; Sardans, J. The bioelements, the elementome, and the biogeochemical niche. Ecology 2019, 100, e02652. [Google Scholar] [CrossRef]
- Uluozlu, O.D.; Tuzen, M.; Mendil, D.; Soylak, M. Assessment of trace element contents of chicken products from turkey. J. Hazard. Mater. 2009, 163, 982–987. [Google Scholar] [CrossRef]
- Hay, M.C.; Hinsu, A.T.; Koringa, P.G.; Pandit, R.J.; Liu, P.-Y.; Parekh, M.J.; Jakhesara, S.J.; Dai, X.; Crotta, M.; Fosso, B.; et al. Chicken caecal enterotypes in indigenous Kadaknath and commercial Cobb chicken lines are associated with Campylobacter abundance and influenced by farming practices. Front. Microbiomes 2023, 2, 1301609. [Google Scholar] [CrossRef]
- Marcolla, C.S.; Marcolla, C.S.; Ju, T.; Ju, T.; Lantz, H.L.; Lantz, H.L.; Willing, B.P.; Willing, B.P. Investigating the cecal microbiota of broilers raised in extensive and intensive production systems. Microbiol. Spectr. 2023, 11, e0235223. [Google Scholar] [CrossRef]
- Voit, E.O.; Olivença, D.V. Discrete Biochemical Systems Theory. Front. Mol. Biosci. 2022, 9, 874669. [Google Scholar] [CrossRef] [PubMed]
- Huynh, U.; Zastrow, M.L. Metallobiology of Lactobacillaceae in the gut microbiome. J. Inorg. Biochem. 2023, 238, 112023. [Google Scholar] [CrossRef] [PubMed]
- Huynh, U.; Nguyen, H.N.; Trinh, B.K.; Elhaj, J.; Zastrow, M.L. A bioinformatic analysis of zinc transporters in intestinal Lactobacillaceae. Metallomics 2023, 15, mfad044. [Google Scholar] [CrossRef] [PubMed]
- González, A.; Gálvez, N.; Martín, J.; Reyes, F.; Pérez-Victoria, I.; Dominguez-Vera, J.M. Identification of the key excreted molecule by Lactobacillus fermentum related to host iron absorption. Food Chem. 2017, 228, 374–380. [Google Scholar] [CrossRef] [PubMed]
- Ohlsson, C.; Engdahl, C.; Fåk, F.; Andersson, A.; Windahl, S.H.; Farman, H.H.; Movérare-Skrtic, S.; Islander, U.; Sjögren, K. Probiotics protect mice from ovariectomy-induced cortical bone loss. PLoS ONE 2014, 9, e92368. [Google Scholar] [CrossRef]
- Asemi, Z.; Esmaillzadeh, A. Effect of daily consumption of probiotic yoghurt on serum levels of calcium, Iron and liver enzymes in pregnant women. Int. J. Prev. Med. 2013, 4, 949–955. [Google Scholar]
- Chamniansawat, S.; Suksridechacin, N.; Thongon, N. Current opinion on the regulation of small intestinal magnesium absorption. World J. Gastroenterol. 2023, 29, 332–342. [Google Scholar] [CrossRef]
- Ma, Y.; Fei, Y.; Ding, S.; Jiang, H.; Fang, J.; Liu, G. Trace metal elements: A bridge between host and intestinal microorganisms. Sci. China Life Sci. 2023, 66, 1976–1993. [Google Scholar] [CrossRef]
- Jurburg, S.D.; Brouwer, M.S.M.; Ceccarelli, D.; van der Goot, J.; Jansman, A.J.M.; Bossers, A. Patterns of community assembly in the developing chicken microbiome reveal rapid primary succession. Microbiologyopen 2019, 8, e00821. [Google Scholar] [CrossRef]
- Ramos-Vidales, D.; Gómez-Verduzco, G.; Cortes-Cuevas, A.; del Río-García, J.C.; Fernández-Tinoco, S.; Chárraga-Aguilar, S.; Ávila-González, E. Organic trace minerals on productive performance, egg quality and immune response in Bovans White laying hens. J. Anim. Physiol. Anim. Nutr. 2019, 103, 1484–1491. [Google Scholar] [CrossRef]
- Hassan, R.I.M.; Refaie, M.S.; El-Shoukary, R.D.; Rehan, I.F.; Zigo, F.; Karaffová, V.; Amer, H.Y. Effect of Dietary Microalgae (Spirulina platensis) on Growth Performance, Ingestive Behavior, Hemato-Biochemical Parameters, and Economic Efficiency of Fayoumi Broilers. Life 2022, 12, 1892. [Google Scholar] [CrossRef] [PubMed]
- Khalid, A.; Huang, Z.; Khan, I.M.; Khalid, F.; Nassar, N.; Jiang, X.; Cheng, M.; Zhan, K.; Wang, Z. Improving broiler health through cecal microbiota transplantation: A comprehensive study on growth, immunity, and microbial diversity. J. Anim. Sci. 2024, 102, skae131. [Google Scholar] [CrossRef] [PubMed]
- Kogut, M.H. Role of diet-microbiota interactions in precision nutrition of the chicken: Facts, gaps, and new concepts. Poult. Sci. 2022, 101, 101673. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.J.; Zhou, Y.M.; Wu, Y.N.; Wang, T. Intestinal development and function of broiler chickens on diets supplemented with clinoptilolite. Asian-Australas. J. Anim. Sci. 2013, 26, 987–994. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.B.; Ferket, P.; Park, I.; Malheiros, R.D.; Kim, S.W. Effects of feed grade L-methionine on intestinal redox status, intestinal development, and growth performance of young chickens compared with conventional DL-methionine. J. Anim. Sci. 2015, 93, 2977–2986. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Element | Liver | Thigh | Breast | Maximum Permissible Level (FAO/WHO) | |
|---|---|---|---|---|---|
| Liver | Meat | ||||
| Macro elements (‰) | |||||
| Na | 1.11 [0.95–3.16] | 0.79 [0.69–2.82] * | 0.58 [0.49–1.33] * | NA | NA |
| K | 3.04 [2.72–7.17] | 4.27 [4.01–9.34] * | 4.40 [4.00–7.37] * | NA | NA |
| Mg | 0.24 [0.21–0.58] | 0.31 [0.29–1.08] * | 0.34 [0.32–0.97] * | NA | NA |
| Ca | 0.08 [0.07–0.22] | 0.06 [0.05–0.18] * | 0.07 [0.05–0.16] | NA | NA |
| P | 3.42 [3.09–7.70] | 2.19 [2.09–6.27] * | 2.48 [2.31–7.72] * | NA | NA |
| Essential trace elements (ppm) | |||||
| Li | 0.01 [0.00–0.01] | 0.01 [0.00–0.01] | 0.01 [0.00–0.01] | NA | NA |
| Sr | 0.09 [0.06–0.27] | 0.05 [0.04–0.16] | 0.07 [0.05–0.28] | NA | NA |
| Si | 16.72 [6.24–33.97] | 24.72 [6.40–29.70] | 22.16 [10.40–26.99] | NA | NA |
| Se | 0.77 [0.67–1.98] | 0.19 [0.18–0.48] * | 0.18 [0.15–0.40] * | NA | 0.5 |
| I | 0.12 [0.05–0.20] | 0.08 [0.02–0.13] | 0.12 [0.03–0.29] | NA | NA |
| V | 0.04 [0.01–0.07] | 0.01 [0.01–0.02] * | 0.01 [0.01–0.02] * | NA | NA |
| Cr | 0.27 [0.17–0.39] | 0.26 [0.12–0.31] | 0.29 [0.15–0.32] | 0.05 | 1.00 |
| Mn | 3.40 [2.98–8.59] | 0.21 [0.18–0.57] * | 0.21 [0.17–0.57] * | 0.50 | 0.50 |
| Fe | 291.83 [232.20–656.22] | 19.72 [13.02–36.76] * | 25.84 [14.28–34.98] * | NA | NA |
| Co | 0.02 [0.02–0.04] | 0.00 [0.00–0.01] * | 0.00 [0.00–0.01] * | NA | NA |
| Cu | 4.49 [3.92–11.25] | 0.66 [0.49–2.11] * | 0.46 [0.42–1.08] * | 1.00 | 1.00 |
| Zn | 36.41 [29.68–92.26] | 19.24 [16.95–53.47] * | 19.53 [9.02–24.57] * | 20.00 | 20.00 |
| Toxic elements (ppm) | |||||
| B | 0.21 [0.19–0.46] | 0.16 [0.11–0.27] | 0.17 [0.13–0.33] | NA | 10.00 |
| Al | 1.09 [0.86–1.37] | 1.35 [0.90–3.17] | 1.91 [1.28–3.61] * | NA | 1.00 |
| Sn | 0.01 [0.01–0.02] | 0.00 [0.00–0.01] | 0.01 [0.01–0.02] | NA | NA |
| Pb | 0.02 [0.01–0.03] | 0.02 [0.01–0.02] | 0.02 [0.01–0.03] | 0.10 | 0.10 |
| As | 0.01 [0.00–0.01] | 0.00 [0.00–0.01] | 0.00 [0.00–0.01] | NA | 0.10 |
| Ni | 0.04 [0.02–0.21] | 0.03 [0.01–0.06] | 0.04 [0.02–0.06] | NA | 0.50 |
| Cd | 0.03 [0.02–0.19] | 0.00 [0.00–0.00] * | 0.00 [0.00–0.04] * | 0.50 | 0.05 |
| Hg | 0.00 [0.00–0.01] | 0.00 [0.00–0.01] | 0.01 [0.00–0.01] | NA | NA |
| Genera | Occurrence (%) | Abundance (%) * |
|---|---|---|
| Belonging to the phylum Bacteroidota | ||
| Bacteroides | 100 | 8.04 [0.22–24.75] |
| Alistipes | 72.22 | 6.77 [0.0–27.69] |
| Belonging to the phylum Bacillota | ||
| Butyricicoccus | 100 | 0.93 [0.34–1.39] |
| Faecalibacterium | 100 | 4.93 [3.02–7.16] |
| Lactobacillus | 100 | 2.79 [0.93–4.32] |
| Ruminococcus | 100 | 0.58 [0.28–1.47] |
| uncassified Lachnospiraceae | 100 | 3.59 [3.14–4.35] |
| unclassified Oscillospiraceae | 100 | 17.40 [11.87–21.44] |
| Christensenella | 97.22 | 1.44 [0.12–3.49] |
| Fusicatenibacter | 97.22 | 0.40 [0.12–0.55] |
| Romboutsia | 97.22 | 0.45 [0.15–0.90] |
| Subdoligranulum | 97.22 | 1.02 [0.38–1.78] |
| Eisenbergiella | 94.44 | 1.17 [0.15–2.98] |
| Turicibacter | 88.89 | 0.35 [0.04–2.35] |
| Flavonifractor | 77.78 | 0.42 [0.03–0.64] |
| Intestinimonas | 72.22 | 0.39 [0.0–1.69] |
| Anaerotignum | 63.89 | 0.59 [0.0–1.04] |
| Dysosmobacter | 61.11 | 0.54 [0.0–1.10] |
| Mediterraneibacter | 61.11 | 0.99 [0.0–2.16] |
| Neglectibacter | 61.11 | 0.34 [0.0–0.59] |
| Genera | Abundance in F1(−) Microbiomes, % | Abundance in F1(+) Microbiomes, % | p Value |
|---|---|---|---|
| Bacteroides | 13.72 [4.61–35.68] | 0.48 [0.20–22.87] | 2.7 × 10−2 |
| Alistipes | 35.95 [12.66–44.73] | 2.94 [0–8.08] | 1.2 × 10−4 |
| Butyricicoccus | 0.26 [0.18–0.53] | 1.28 [0.93–1.54] | 4.6 × 10−5 |
| Faecalibacterium | 5.02 [4.65–7.96] | 4.26 [2.64–5.45] | >0.05 |
| Lactobacillus | 4.24 [3.04–6.72] | 1.57 [0.57–2.99] | 3.1 × 10−3 |
| Ruminococcus | 0.93 [0.35–1.53] | 0.38 [0.19–0.99] | >0.05 |
| uncassified Lachnospiraceae | 3.31 [1.68–3.89] | 3.80 [3.22–4.53] | >0.05 |
| unclassified Oscillospiraceae | 11.54 [7.97–14.67] | 20.10 [17.45–21.44] | 2.9 × 10−4 |
| Christensenella | 0.06 [0.02–0.20] | 3.09 [1.44–4.62] | 9.4 × 10−6 |
| Fusicatenibacter | 0.60 [0.05–0.87] | 0.32 [0.14–0.50] | >0.05 |
| Romboutsia | 0.07 [0.01–0.32] | 0.62 [0.44–1.39] | 2.2 × 10−4 |
| Subdoligranulum | 0.37 [0.21–0.65] | 1.71 [0.94–2.38] | 1.5 × 10−3 |
| Eisenbergiella | 0.09 [0.01–0.43] | 2.69 [1.69–4.94] | 4.4 × 10−6 |
| Turicibacter | 0.03 [0–0.08] | 1.22 [0.41–6.20] | 2.7 × 10−6 |
| Flavonifractor | 0.09 [0.01–0.42] | 0.51 [0.36–0.70] | 4.9 × 10−2 |
| Intestinimonas | 0.01 [0.0–0.04] | 1.24 [0.88–2.45] | 2.7 × 10−3 |
| Anaerotignum | 0.0 [0.0–0.0] | 0.68 [0.59–1.44] | 1.4 × 10−4 |
| Dysosmobacter | 0.0 [0.0–0.0] | 0.99 [0.89–1.68] | 4.1 × 10−7 |
| Mediterraneibacter | 0.0 [0.0–0.0] | 2.12 [1.19–2.92] | 9.8 × 10−7 |
| Neglectibacter | 0.0 [0.0–0.0] | 0.49 [0.37–0.71] | 1.6 × 10−6 |
| Element | Liver | Thigh | Breast | ||||||
|---|---|---|---|---|---|---|---|---|---|
| F1(−) Microbiome Group | F1(+) Microbiome Group | p Value | F1(−) Microbiome Group | F1(+) Microbiome Group | p Value | F1(−) Microbiome Group | F1(+) Microbiome Group | p Value | |
| Macro elements (‰) | |||||||||
| Na | 3.36 [1.56–3.81] | 1.02 [0.87–1.02] | 3.3 × 10−5 | 2.85 [0.84–4.04] | 0.71 [0.66–0.71] | 1.6 × 10−4 | 1.38 [0.83–1.58] | 0.50 [0.48–0.50] | 4.0 × 10−6 |
| K | 7.17 [4.01–8.53] | 2.73 [2.67–2.73] | 1.6 × 10−5 | 9.42 [6.11–11.05] | 4.10 [3.81–4.10] | 1.0 × 10−4 | 7.37 [6.22–7.58] | 4.10 [3.79–4.10] | 1.6 × 10−4 |
| Mg | 0.59 [0.25–0.75] | 0.22 [0.21–0.22] | 3.8 × 10−5 | 1.10 [0.36–1.17] | 0.30 [0.29–0.30] | 6.0 × 10−6 | 0.98 [0.40–1.05] | 0.33 [0.30–0.33] | 8.4 × 10−5 |
| Ca | 0.22 [0.11–0.28] | 0.08 [0.07–0.08] | 5.0 × 10−6 | 0.19 [0.07–0.22] | 0.05 [0.04–0.05] | 1.0 × 10−4 | 0.16 [0.07–0.21] | 0.06 [0.05–0.06] | 6.6 × 10−4 |
| P | 8.03 [3.94–11.66] | 3.24 [3.00–3.24] | 1,1 × 10−4 | 6.38 [2.91–7.20] | 2.10 [2.04–2.10] | 1.1 × 10−5 | 7.72 [3.17–8.24] | 2.35 [2.01–2.35] | 4.3 × 10−5 |
| Essential trace elements (ppm) | |||||||||
| Li | 0.01 [0.00–0.05] | 0.01 [0.00–0.01] | >0.05 | 0.00 [0.00–0.02] | 0.01 [0.01–0.01] | >0.05 | 0.00 [0.00–0.01] | 0.01 [0.00–0.01] | >0.05 |
| Sr | 0.28 [0.12–0.38] | 0.06 [0.05–0.06] | 1.2 × 10−4 | 0.20 [0.05–0.32] | 0.05 [0.05–0.05] | >0.01 | 0.26 [0.06–0.29] | 0.06 [0.05–0.06] | >0.05 |
| Si | 6.15 [4.42–7.03] | 29.32 [17.53–29.32] | 1.4 × 10−4 | 5.70 [4.62–10.04] | 28.53 [24.79–28.53] | 8.5 × 10−5 | 10.40 [7.59–13.70] | 24.60 [22.63–24.60] | 2.1 × 10−4 |
| Se | 2.24 [0.80–2.47] | 0.70 [0.67–0.70] | 3.0 × 10−5 | 0.51 [0.22–0.52] | 0.19 [0.17–0.19] | >0.001 | 0.40 [0.24–0.45] | 0.16 [0.14–0.16] | >0.001 |
| I | 0.20 [0.18–0.29] | 0.05 [0.04–0.05] | 4.0 × 10−6 | 0.13 [0.12–0.16] | 0.02 [0.02–0.02] | 8.0 × 10−5 | 0.30 [0.16–0.66] | 0.04 [0.01–0.04] | 1.6 × 10−5 |
| V | 0.01 [0.01–0.04] | 0.05 [0.05–0.05] | 1.7 × 10−4 | 0.01 [0.01–0.01] | 0.02 [0.02–0.02] | >0.001 | 0.01 [0.01–0.02] | 0.01 [0.01–0.01] | >0.05 |
| Cr | 0.17 [0.09–0.37] | 0.32 [0.27–0.32] | >0.01 | 0.12 [0.10–0.16] | 0.31 [0.29–0.31] | 6.0 × 10−6 | 0.14 [0.11–0.27] | 0.32 [0.30–0.32] | 4.3 × 10−4 |
| Mn | 10.17 [3.90–13.18] | 3.13 [2.70–3.13] | 1.6 × 10−4 | 0.59 [0.33–0.68] | 0.20 [0.18–0.20] | 4.7 × 10−4 | 0.57 [0.28–0.61] | 0.19 [0.16–0.19] | 6.8 × 10−4 |
| Fe | 833.00 [416.00–1342.00] | 234.50 [213.25–234.50] | 1.2 × 10−5 | 33.94 [14.25–42.83] | 17.27 [12.84–17.27] | >0.01 | 24.20 [17.20–27.98] | 26.34 [13.80–26.34] | >0.05 |
| Co | 0.05 [0.02–0.05] | 0.02 [0.01–0.02] | 2.5 × 10−4 | 0.01 [0.00–0.01] | 0.00 [0.00–0.00] | >0.001 | 0.01 [0.01–0.01] | 0.00 [0.00–0.00] | >0.001 |
| Cu | 12.01 [4.72–13.55] | 3.98 [3.83–3.98] | 2.9 × 10−5 | 2.16 [0.72–2.57] | 0.57 [0.49–0.57] | 5.4 × 10−4 | 1.08 [0.49–1.39] | 0.43 [0.36–0.43] | 1.5 × 10−4 |
| Zn | 94.24 [41.40–114.00] | 30.88 [25.68–30.88] | 3.9 × 10−4 | 54.30 [14.90–57.87] | 18.31 [17.36–18.31] | >0.01 | 24.49 [9.44–26.99] | 14.20 [8.84–14.20] | >0.01 |
| Toxic elements (ppm) | |||||||||
| B | 0.48 [0.20–0.49] | 0.20 [0.19–0.20] | >0.01 | 0.50 [0.17–0.66] | 0.15 [0.12–0.15] | >0.001 | 0.41 [0.14–0.55] | 0.17 [0.13–0.17] | >0.01 |
| Al | 1.32 [1.14–1.66] | 0.86 [0.77–0.86] | 2.2 × 10−4 | 3.55 [1.14–5.41] | 1.21 [0.91–1.21] | >0.01 | 3.71 [1.24–5.13] | 1.71 [1.32–1.71] | >0.05 |
| Sn | 0.02 [0.01–0.04] | 0.01 [0.01–0.01] | >0.001 | 0.01 [0.01–0.02] | 0.00 [0.00–0.00] | 8.1 × 10−4 | 0.02 [0.01–0.03] | 0.01 [0.01–0.01] | >0.01 |
| Pb | 0.03 [0.02–0.05] | 0.01 [0.01–0.01] | 6.7 × 10−5 | 0.02 [0.02–0.04] | 0.02 [0.02–0.02] | >0.05 | 0.03 [0.02–0.04] | 0.02 [0.02–0.02] | >0.05 |
| As | 0.01 [0.01–0.02] | 0.01 [0.00–0.01] | >0.001 | 0.01 [0.01–0.01] | 0.00 [0.00–0.00] | 6.0 × 10−6 | 0.01 [0.01–0.01] | 0.00 [0.00–0.00] | 8.0 × 10−6 |
| Ni | 0.20 [0.06–0.34] | 0.04 [0.01–0.04] | 5.8 × 10−4 | 0.05 [0.02–0.07] | 0.03 [0.01–0.03] | >0.01 | 0.04 [0.02–0.10] | 0.04 [0.02–0.04] | >0.05 |
| Cd | 0.26 [0.03–1.34] | 0.03 [0.02–0.03] | >0.01 | 0.00 [0.00–0.00] | 0.00 [0.00–0.00] | >0.05 | 0.06 [0.00–0.10] | 0.00 [0.00–0.00] | 9.7 × 10−4 |
| Hg | 0.01 [0.00–0.01] | 0.00 [0.00–0.00] | >0.05 | 0.01 [0.00–0.02] | 0.00 [0.00–0.00] | >0.01 | 0.00 [0.00–0.03] | 0.01 [0.01–0.01] | >0.05 |
| Element | Bacterial Phyla in Chicken’s Cecal Microbiomes *** | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| A | B1 | B2 | C1 | C2 | M | P | T | V | Unc | |
| Macro elements | ||||||||||
| Na | 0.43(L)/0.46(B) | 0.43(L)/0.46(B) | ||||||||
| K | 0.47(L)/0.47(T)/0.47(B) | 0.47(L)/0.44(T)/0.45(B) | 0.39(B) | |||||||
| Mg | 0.40(L)/0.39(T) | |||||||||
| Ca | 0.41(L) | 0.40(L) | 0.52(B) | 0.43(T) | ||||||
| P | 0.41(T)/0.39(B) | 0.40(T)/0.39(B) | 0.50(L) | |||||||
| Essential trace elements | ||||||||||
| Li | 0.70(L)/0.42(T)/0.76(B) | |||||||||
| Sr | ||||||||||
| Si | 0.42(L) | 0.44(L) | ||||||||
| Se | 0.46(L) | |||||||||
| I | 0.51(L) | 0.52(L) | 0.54(B) | |||||||
| V | 0.41(L) | 0.64(L)/0.40(T) | 0.69(L) | |||||||
| Cr | 0.74(T) | 0.72(T) | 0.46(L) | |||||||
| Mn | 0.39(T)/0.43(B) | 0.42(B) | 0.52(L) | |||||||
| Fe | 0.43(B) | 0.41(B) | 0.48(L) | |||||||
| Co | ||||||||||
| Cu | 0.57(T) | |||||||||
| Zn | 0.48(T)/0.40(B) | |||||||||
| Toxic elements | ||||||||||
| B | 0.52(L) | |||||||||
| Al | 0.71(T)/0.82(B) | |||||||||
| Sn | 0.80(L) | |||||||||
| Pb | 0.43(L) | 0.42(L) | ||||||||
| As | 0.55(T)/0.71(B) | 0.53(T)/0.66(B) | 0.44(T)/0.40(B) | 0.39(L) | ||||||
| Ni | 0.39(T) | 0.40(L)/0.59(T) | ||||||||
| Cd | 1.00(T) | 0.57(L)/0.52(B) | ||||||||
| Hg | 0.40(T) | |||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deryabin, D.; Kosyan, D.; Vlasenko, L.; Lazebnik, C.; Zatevalov, A.; Karimov, I.; Duskaev, G. Macro, Trace and Toxic Element Composition in Liver and Meat of Broiler Chicken Associated with Cecal Microbiome Community. Biology 2024, 13, 975. https://doi.org/10.3390/biology13120975
Deryabin D, Kosyan D, Vlasenko L, Lazebnik C, Zatevalov A, Karimov I, Duskaev G. Macro, Trace and Toxic Element Composition in Liver and Meat of Broiler Chicken Associated with Cecal Microbiome Community. Biology. 2024; 13(12):975. https://doi.org/10.3390/biology13120975
Chicago/Turabian StyleDeryabin, Dmitry, Dianna Kosyan, Ludmila Vlasenko, Christina Lazebnik, Alexander Zatevalov, Ilshat Karimov, and Galimzhan Duskaev. 2024. "Macro, Trace and Toxic Element Composition in Liver and Meat of Broiler Chicken Associated with Cecal Microbiome Community" Biology 13, no. 12: 975. https://doi.org/10.3390/biology13120975
APA StyleDeryabin, D., Kosyan, D., Vlasenko, L., Lazebnik, C., Zatevalov, A., Karimov, I., & Duskaev, G. (2024). Macro, Trace and Toxic Element Composition in Liver and Meat of Broiler Chicken Associated with Cecal Microbiome Community. Biology, 13(12), 975. https://doi.org/10.3390/biology13120975