

Diversity and Evolution of NLR Genes in Citrus Species


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Data Used in This Study
2.2. Identification of NLR Genes
2.3. Phylogenetic Analysis
2.4. Positive Selection Analysis
2.5. Recombination Analysis
3. Results
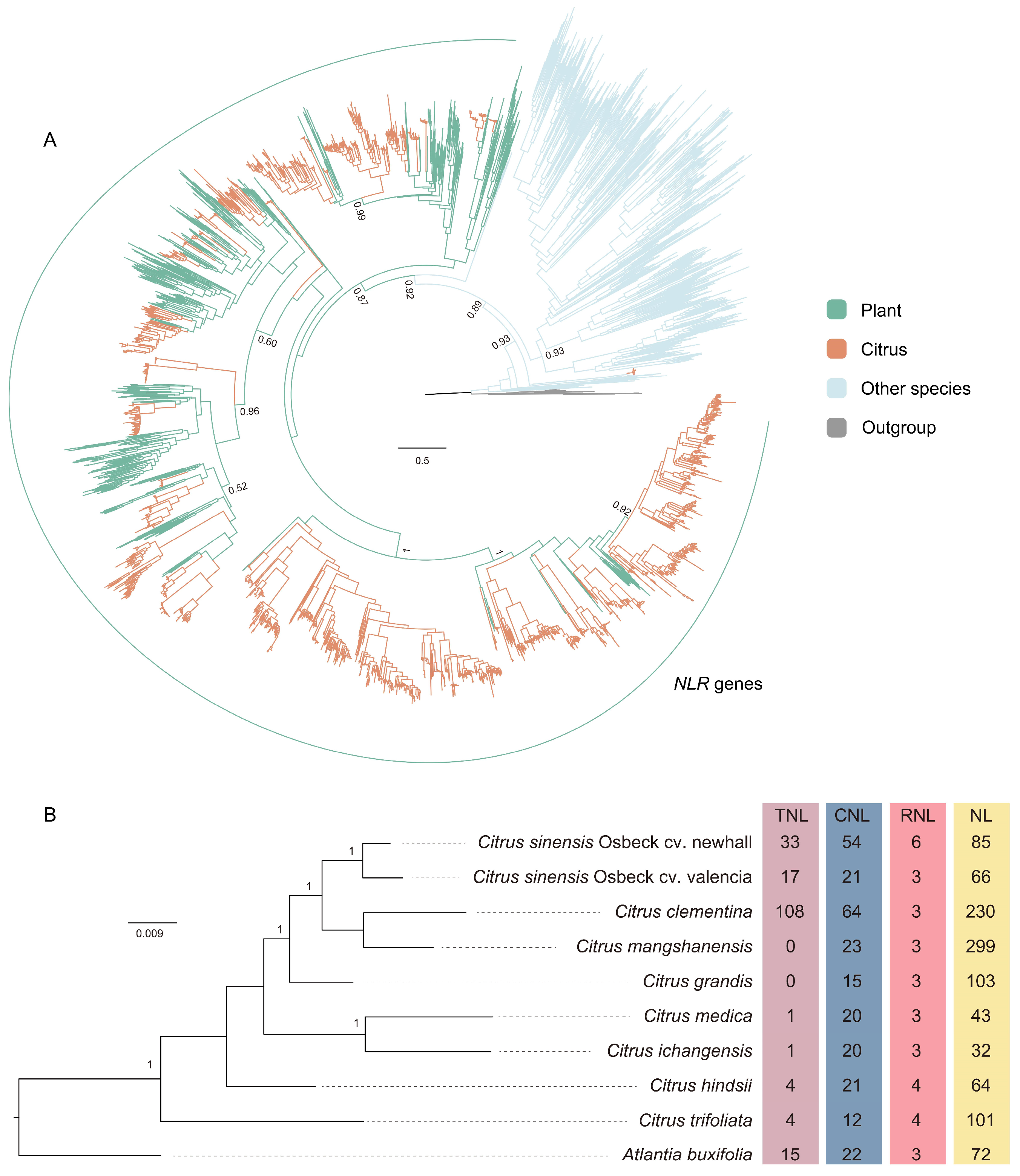
3.1. Identification and Diversity of Citrus NLR Genes
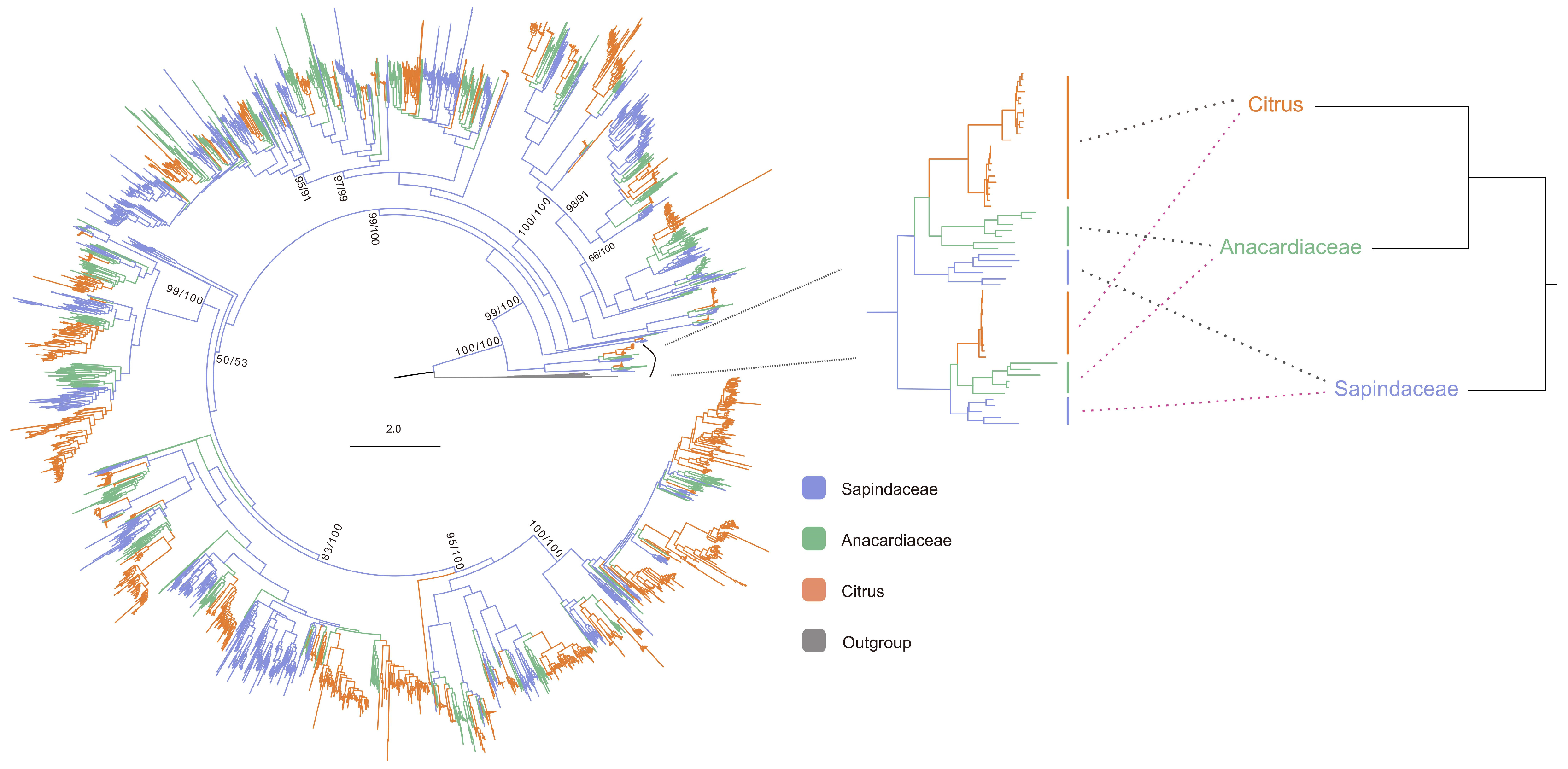
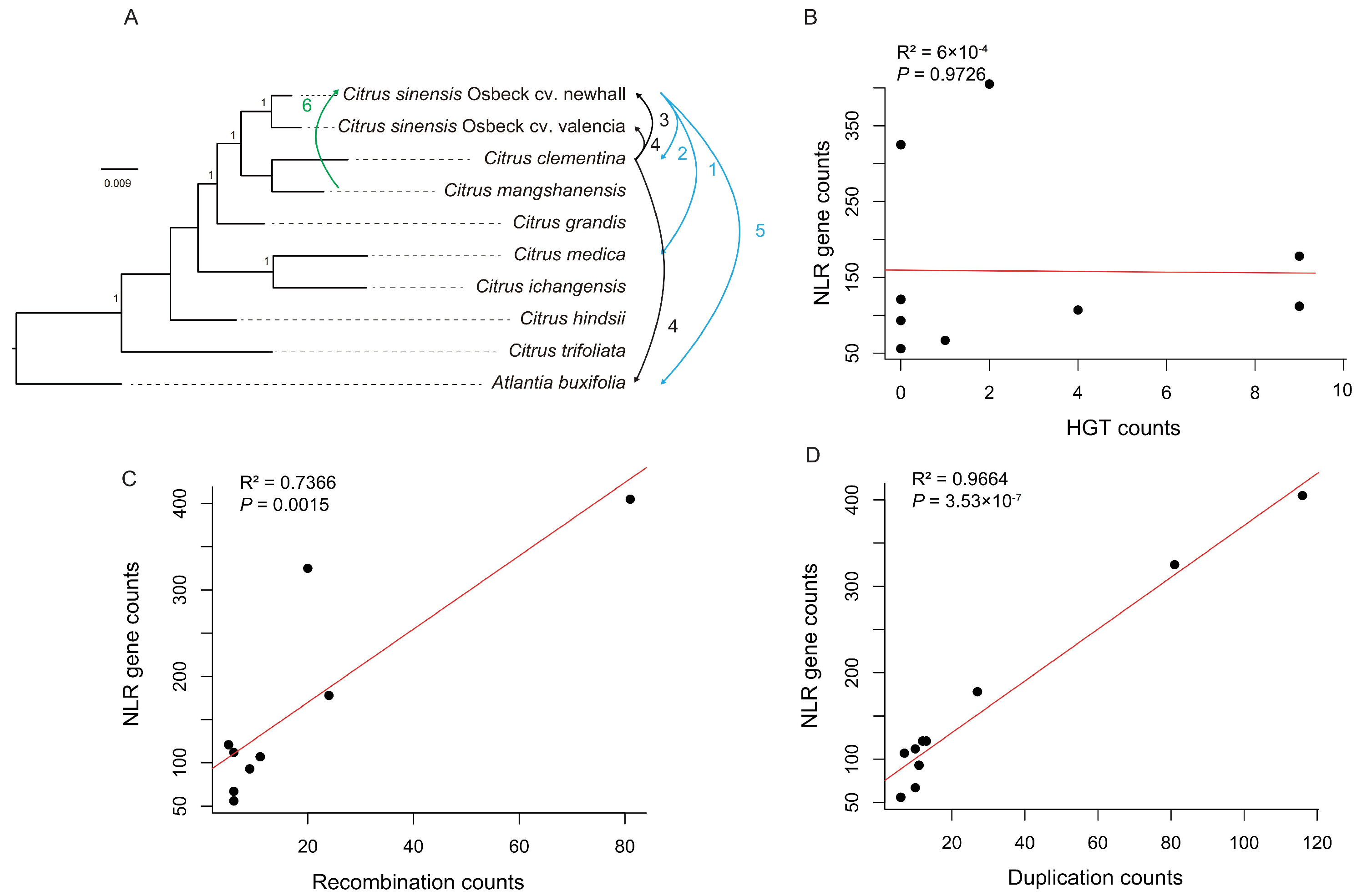
3.2. The Origin of Citrus NLR Genes
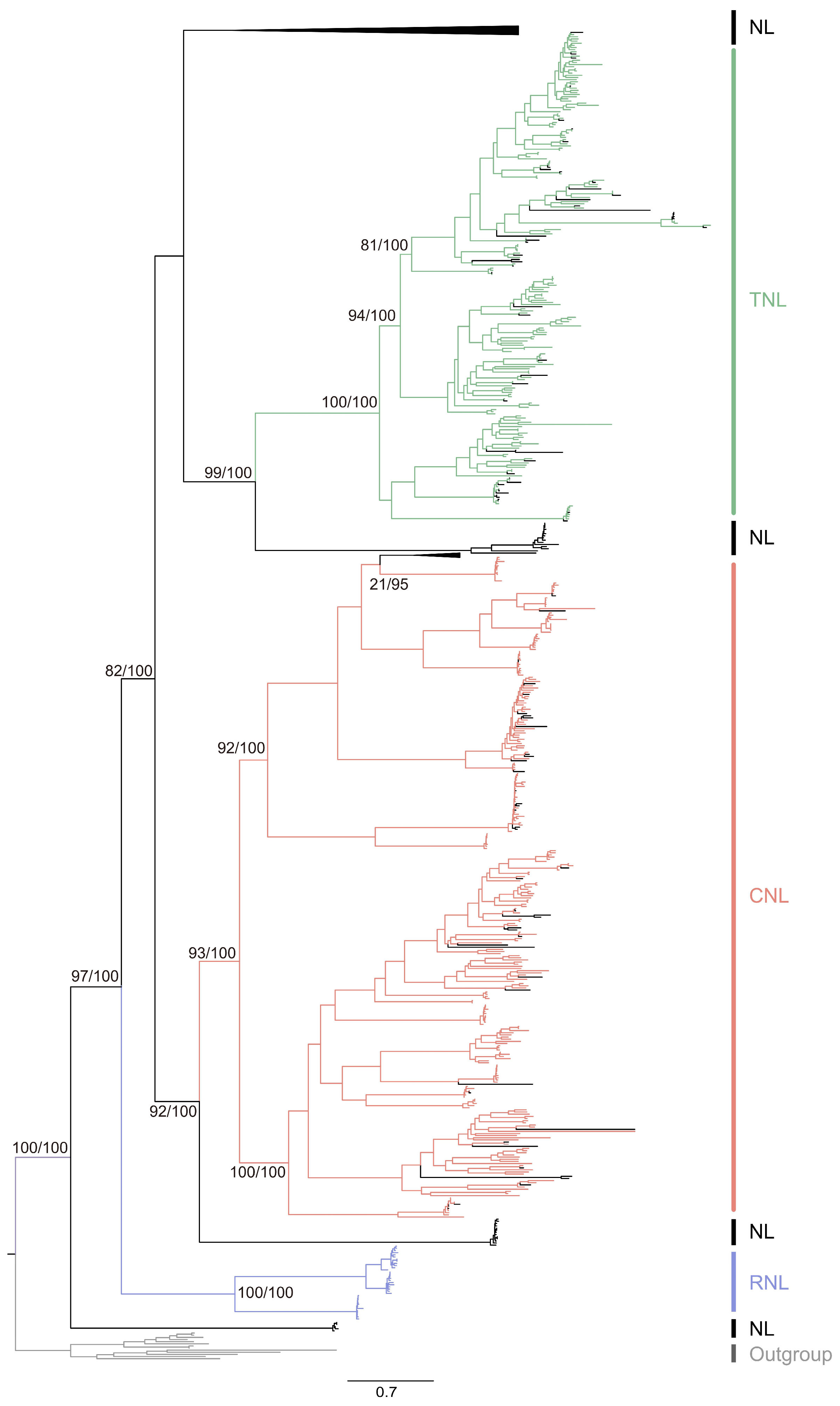
3.3. The Evolution of Citrus NLR Genes
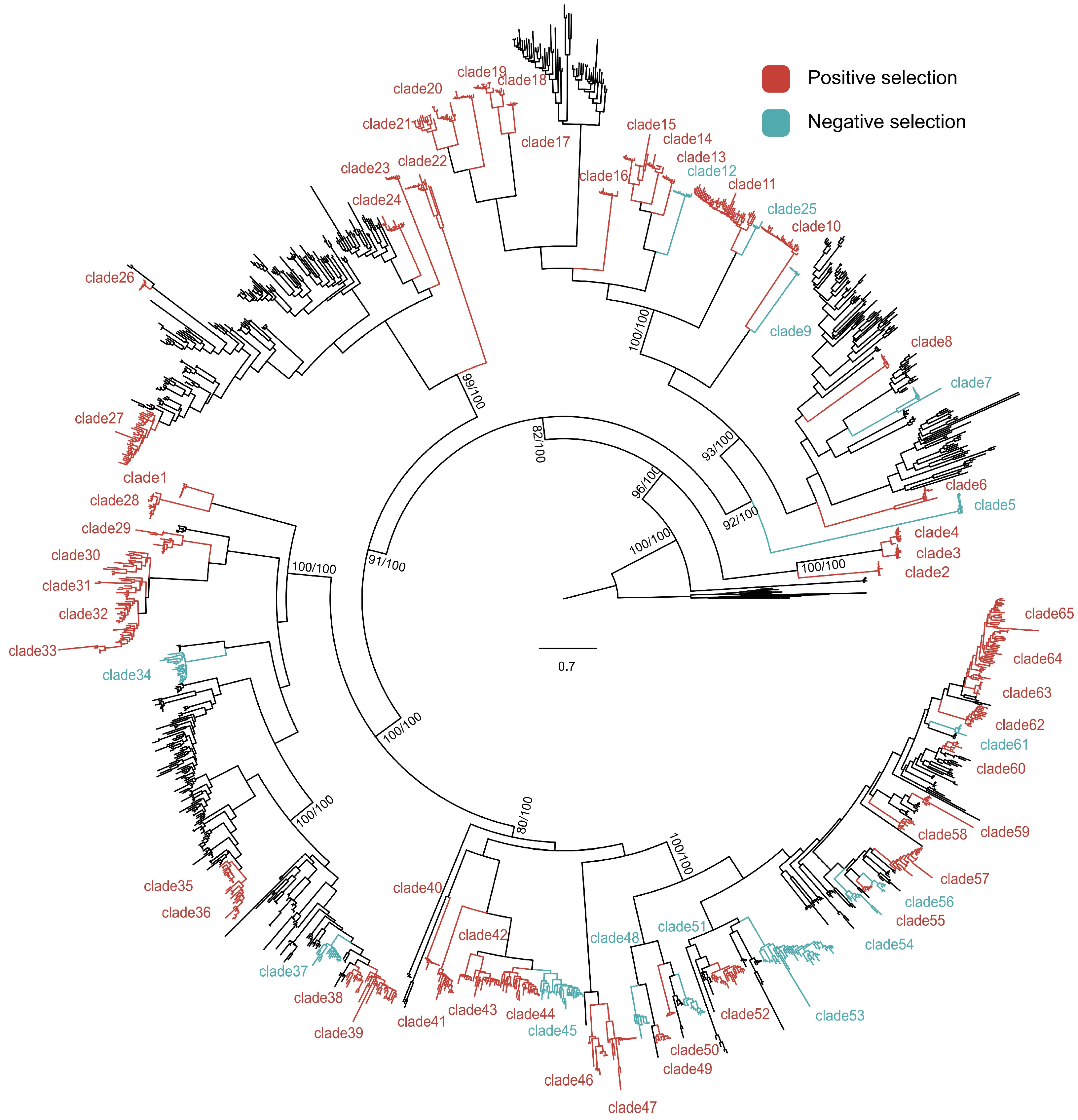
3.4. Mechanism of NLR Gene Diversity Formation in Citrus
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Han, G. Origin and Evolution of the Plant Immune System. New Phytol. 2019, 222, 70–83. [Google Scholar] [CrossRef] [PubMed]
- Ngou, B.P.M.; Ahn, H.-K.; Ding, P.; Jones, J.D.G. Mutual Potentiation of Plant Immunity by Cell-Surface and Intracellular Receptors. Nature 2021, 592, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Liu, X.; Dai, L.; Wang, G. Recent Progress in Elucidating the Structure, Function and Evolution of Disease Resistance Genes in Plants. J. Genet. Genom. 2007, 34, 765–776. [Google Scholar] [CrossRef]
- Bigeard, J.; Colcombet, J.; Hirt, H. Signaling Mechanisms in Pattern-Triggered Immunity (PTI). Mol. Plant 2015, 8, 521–539. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.; Chen, H.; Liu, F.; Fu, Z.Q. PTI and ETI: Convergent Pathways with Diverse Elicitors. Trends Plant Sci. 2022, 27, 113–115. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.D.G.; Dangl, J.L. The Plant Immune System. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.; Jiang, Z.; Bi, G.; Nomura, K.; Liu, M.; Wang, Y.; Cai, B.; Zhou, J.-M.; He, S.Y.; Xin, X.-F. Pattern-Recognition Receptors Are Required for NLR-Mediated Plant Immunity. Nature 2021, 592, 105–109. [Google Scholar] [CrossRef]
- Kim, C.-Y.; Song, H.; Lee, Y.-H. Ambivalent Response in Pathogen Defense: A Double-Edged Sword? Plant Commun. 2022, 3, 100415. [Google Scholar] [CrossRef]
- Bent, A.; Kunkel, B.; Dahlbeck, D.; Brown, K.; Schmidt, R.; Giraudat, J.; Leung, J.; Staskawicz, B. RPS2 of Arabidopsis Thaliana: A Leucine-Rich Repeat Class of Plant Disease Resistance Genes. Science 1994, 265, 1856–1860. [Google Scholar] [CrossRef]
- Jia, Y.; Wang, Z.; Fjellstrom, R.G.; Moldenhauer, K.A.K.; Azam, M.A.; Correll, J.; Lee, F.N.; Xia, Y.; Rutger, J.N. Rice Pi-Ta Gene Confers Resistance to the Major Pathotypes of the Rice Blast Fungus in the United States. Phytopathology 2004, 94, 296–301. [Google Scholar] [CrossRef]
- Chen, J.; Li, M.; Liu, L.; Chen, G.; Fu, Z.Q. ZAR1 Resistosome and Helper NLRs: Bringing in Calcium and Inducing Cell Death. Mol. Plant 2021, 14, 1234–1236. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zeng, Z.; Zhang, Y.; Li, Q.; Jiang, X.-M.; Jiang, Z.; Tang, J.; Chen, D.; Wang, Q.; Chen, J.-Q.; et al. An Angiosperm NLR Atlas Reveals That NLR Gene Reduction Is Associated with Ecological Specialization and Signal Transduction Component Deletion. Mol. Plant 2021, 14, 2015–2031. [Google Scholar] [CrossRef]
- Gao, Y.; Wang, W.; Zhang, T.; Gong, Z.; Zhao, H.; Han, G.-Z. Out of Water: The Origin and Early Diversification of Plant R-Genes. Plant Physiol. 2018, 177, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Xiao, S. Broad-Spectrum Mildew Resistance in Arabidopsis Thaliana Mediated by RPW8. Science 2001, 291, 118–120. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.; Wang, Y.; Wu, P.; Wang, Q.; Yang, L.-T.; Pan, X.-H.; Wang, B.; Chen, J.-Q. A Primary Survey on Bryophyte Species Reveals Two Novel Classes of Nucleotide-Binding Site (NBS) Genes. PLoS ONE 2012, 7, e36700. [Google Scholar] [CrossRef] [PubMed]
- Shao, Z.-Q.; Xue, J.-Y.; Wang, Q.; Wang, B.; Chen, J.-Q. Revisiting the Origin of Plant NBS-LRR Genes. Trends Plant Sci. 2019, 24, 9–12. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zhang, Y.; Zhang, P.; Trivedi, P.; Riera, N.; Wang, Y.; Liu, X.; Fan, G.; Tang, J.; Coletta-Filho, H.D.; et al. The Structure and Function of the Global Citrus Rhizosphere Microbiome. Nat. Commun. 2018, 9, 4894. [Google Scholar] [CrossRef]
- Falaki, F. Citrus Virus and Viroid Diseases. Citrus Research—Horticultural and Human Health Aspects. IntechOpen. 2023. Available online: https://www.intechopen.com/books/11629 (accessed on 8 October 2024).
- Gao, Y.; Xu, J.; Li, Z.; Zhang, Y.; Riera, N.; Xiong, Z.; Ouyang, Z.; Liu, X.; Lu, Z.; Seymour, D.; et al. Citrus Genomic Resources Unravel Putative Genetic Determinants of Huanglongbing Pathogenicity. iScience 2023, 26, 106024. [Google Scholar] [CrossRef]
- Liu, H.; Wang, X.; Liu, S.; Huang, Y.; Guo, Y.-X.; Xie, W.-Z.; Liu, H.; Muhammad; Xu, Q.; Chen, L.-L. Citrus Pan-Genome to Breeding Database (CPBD): A Comprehensive Genome Database for Citrus Breeding. Mol. Plant 2022, 15, 1503–1505. [Google Scholar] [CrossRef]
- Eddy, S.R. Accelerated Profile HMM Searches. PLoS Comput. Biol. 2011, 7, e1002195. [Google Scholar] [CrossRef]
- Urbach, J.M.; Ausubel, F.M. The NBS-LRR Architectures of Plant R-Proteins and Metazoan NLRs Evolved in Independent Events. Proc. Natl. Acad. Sci. USA 2017, 114, 1063–1068. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast Model Selection for Accurate Phylogenetic Estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed]
- Marchler-Bauer, A.; Bo, Y.; Han, L.; He, J.; Lanczycki, C.J.; Lu, S.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; et al. CDD/SPARCLE: Functional Classification of Proteins via Subfamily Domain Architectures. Nucleic Acids Res. 2017, 45, D200–D203. [Google Scholar] [CrossRef]
- Zhang, W. NLR-Annotator: A Tool for de Novo Annotation of Intracellular Immune Receptor Repertoire. Plant Physiol. 2020, 183, 418–420. [Google Scholar] [CrossRef]
- Emms, D.M.; Kelly, S. OrthoFinder: Phylogenetic Orthology Inference for Comparative Genomics. Genome Biol. 2019, 20, 238. [Google Scholar] [CrossRef]
- Yang, Z. PAML 4: Phylogenetic Analysis by Maximum Likelihood. Mol. Biol. Evol. 2007, 24, 1586–1591. [Google Scholar] [CrossRef]
- Yang, Z.; Nielsen, R.; Goldman, N.; Pedersen, A.-M.K. Codon-Substitution Models for Heterogeneous Selection Pressure at Amino Acid Sites. Genetics 2000, 155, 431–449. [Google Scholar] [CrossRef]
- Martin, D.P.; Varsani, A.; Roumagnac, P.; Botha, G.; Maslamoney, S.; Schwab, T.; Kelz, Z.; Kumar, V.; Murrell, B. RDP5: A Computer Program for Analyzing Recombination In, and Removing Signals of Recombination From, Nucleotide Sequence Datasets. Virus Evol. 2020, 7, veaa087. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Zhang, S.; Mei, S.; Zhou, Y.; Wang, J.; Han, G.-Z.; Chen, L.; Zhou, C.; Cao, M. Viromics Unveils Extraordinary Genetic Diversity of the Family Closteroviridae in Wild Citrus. PLoS Pathog. 2021, 17, e1009751. [Google Scholar] [CrossRef] [PubMed]
- Hao, W.; Collier, S.A.; Moffett, P.; Chai, J. Structural Basis for the Interaction between the Potato Virus X Resistance Protein (Rx) and Its Cofactor Ran GTPase-Activating Protein 2 (RanGAP2). J. Biol. Chem. 2013, 288, 35868–35876. [Google Scholar] [CrossRef] [PubMed]
- Qian, L.-H.; Wang, Y.; Chen, M.; Liu, J.; Lu, R.-S.; Zou, X.; Sun, X.-Q.; Zhang, Y.-M. Genome-Wide Identification and Evolutionary Analysis of NBS-LRR Genes from Secale Cereale. Front. Genet. 2021, 12, 771814. [Google Scholar] [CrossRef] [PubMed]
- Joyce, E.M.; Appelhans, M.S.; Buerki, S.; Cheek, M.; de Vos, J.M.; Pirani, J.R.; Zuntini, A.R.; Bachelier, J.B.; Bayly, M.J.; Callmander, M.W.; et al. Phylogenomic Analyses of Sapindales Support New Family Relationships, Rapid Mid-Cretaceous Hothouse Diversification, and Heterogeneous Histories of Gene Duplication. Front. Plant Sci. 2023, 14, 1063174. [Google Scholar] [CrossRef]
- Huang, Y.; He, J.; Xu, Y.; Zheng, W.; Wang, S.; Chen, P.; Zeng, B.; Yang, S.; Jiang, X.; Liu, Z.X.; et al. Pangenome Analysis Provides Insight into the Evolution of the Orange Subfamily and a Key Gene for Citric Acid Accumulation in Citrus Fruits. Nat. Genet. 2023, 55, 1964–1975. [Google Scholar] [CrossRef]
- Märkle, H.; Saur, I.M.L.; Stam, R. Evolution of Resistance (R) Gene Specificity. Essays Biochem. 2022, 66, 551–560. [Google Scholar] [CrossRef]
- Xu, Q.; Chen, L.-L.; Ruan, X.; Chen, D.; Zhu, A.; Chen, C.; Bertrand, D.; Jiao, W.-B.; Hao, B.-H.; Lyon, M.P.; et al. The Draft Genome of Sweet Orange (Citrus Sinensis). Nat. Genet. 2012, 45, 59–66. [Google Scholar] [CrossRef]
- Wang, L.; Huang, Y.; Liu, Z.; He, J.; Jiang, X.; He, F.; Lu, Z.; Yang, S.; Chen, P.; Yu, H.; et al. Somatic Variations Led to the Selection of Acidic and Acidless Orange Cultivars. Nat. Plants 2021, 7, 954–965. [Google Scholar] [CrossRef]
- Zhong, Y.; Yin, H.; Sargent, D.J.; Malnoy, M.; Cheng, Z. Species-Specific Duplications Driving the Recent Expansion of NBS-LRR Genes in Five Rosaceae Species. BMC Genom. 2015, 16, 77. [Google Scholar] [CrossRef]
- Tarr, D.E.K.; Alexander, H.M. TIR-NBS-LRR Genes Are Rare in Monocots: Evidence from Diverse Monocot Orders. BMC Res. Notes 2009, 2, 197. [Google Scholar] [CrossRef] [PubMed]
- Rensing, S.A.; Lang, D.; Zimmer, A.D.; Terry, A.; Salamov, A.; Shapiro, H.; Nishiyama, T.; Perroud, P.-F.; Lindquist, E.A.; Kamisugi, Y.; et al. The Physcomitrella Genome Reveals Evolutionary Insights into the Conquest of Land by Plants. Science 2007, 319, 64–69. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiong, Z.; Zhang, W.; Yin, H.; Wan, J.; Wu, Z.; Gao, Y. Diversity and Evolution of NLR Genes in Citrus Species. Biology 2024, 13, 822. https://doi.org/10.3390/biology13100822
Xiong Z, Zhang W, Yin H, Wan J, Wu Z, Gao Y. Diversity and Evolution of NLR Genes in Citrus Species. Biology. 2024; 13(10):822. https://doi.org/10.3390/biology13100822
Chicago/Turabian StyleXiong, Zhiwei, Wanshan Zhang, Hui Yin, Jiaxing Wan, Zhuozhuo Wu, and Yuxia Gao. 2024. "Diversity and Evolution of NLR Genes in Citrus Species" Biology 13, no. 10: 822. https://doi.org/10.3390/biology13100822
APA StyleXiong, Z., Zhang, W., Yin, H., Wan, J., Wu, Z., & Gao, Y. (2024). Diversity and Evolution of NLR Genes in Citrus Species. Biology, 13(10), 822. https://doi.org/10.3390/biology13100822