
Ontogenetic Shifts in Body Morphology of Demersal Sharks’ Species (Order: Squaliformes) Inhabiting the Western-Central Mediterranean Sea, with Implications for Their Bio-Ecological Role

 ,
,  , ,
, ,  ,
,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling
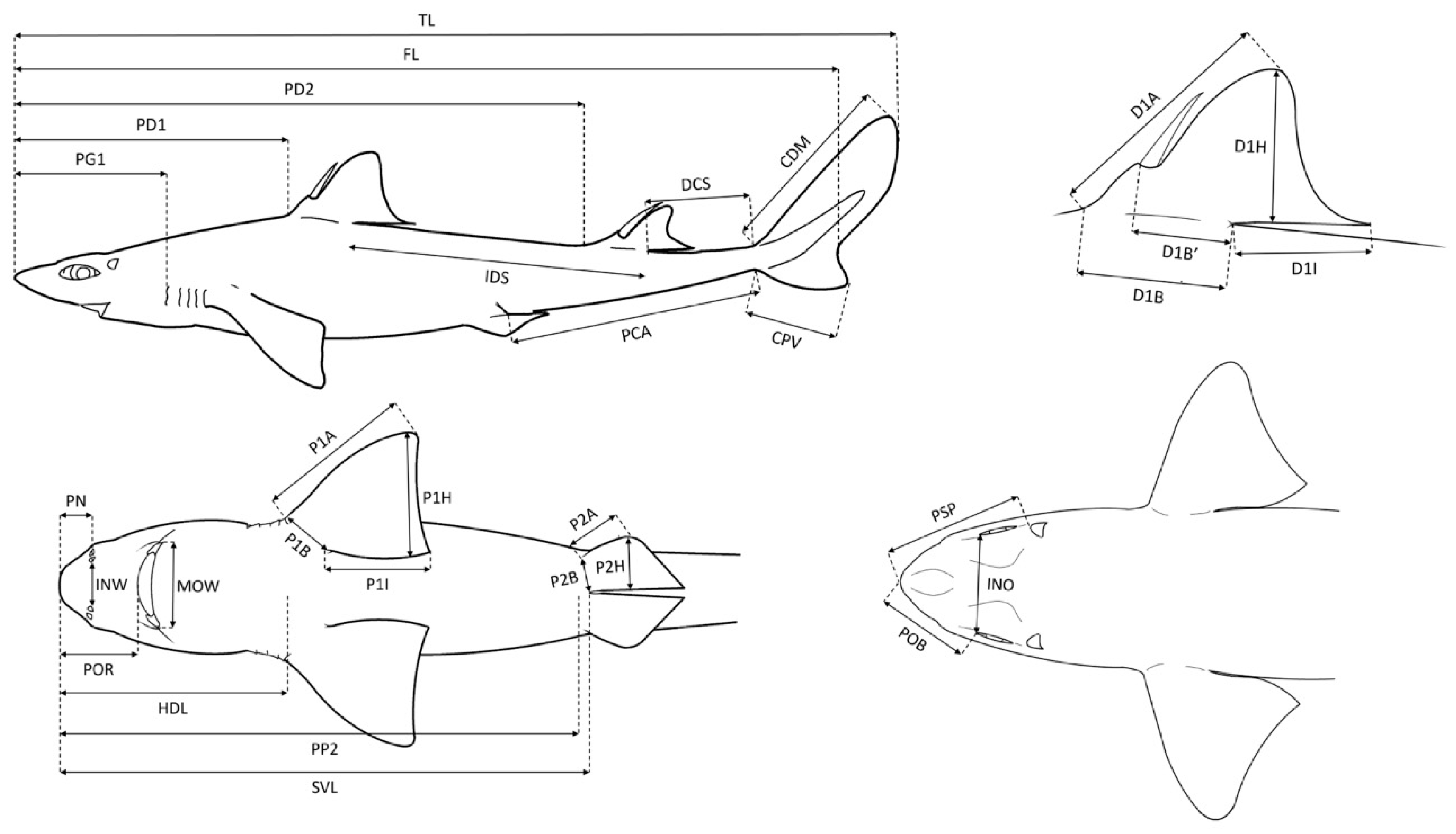
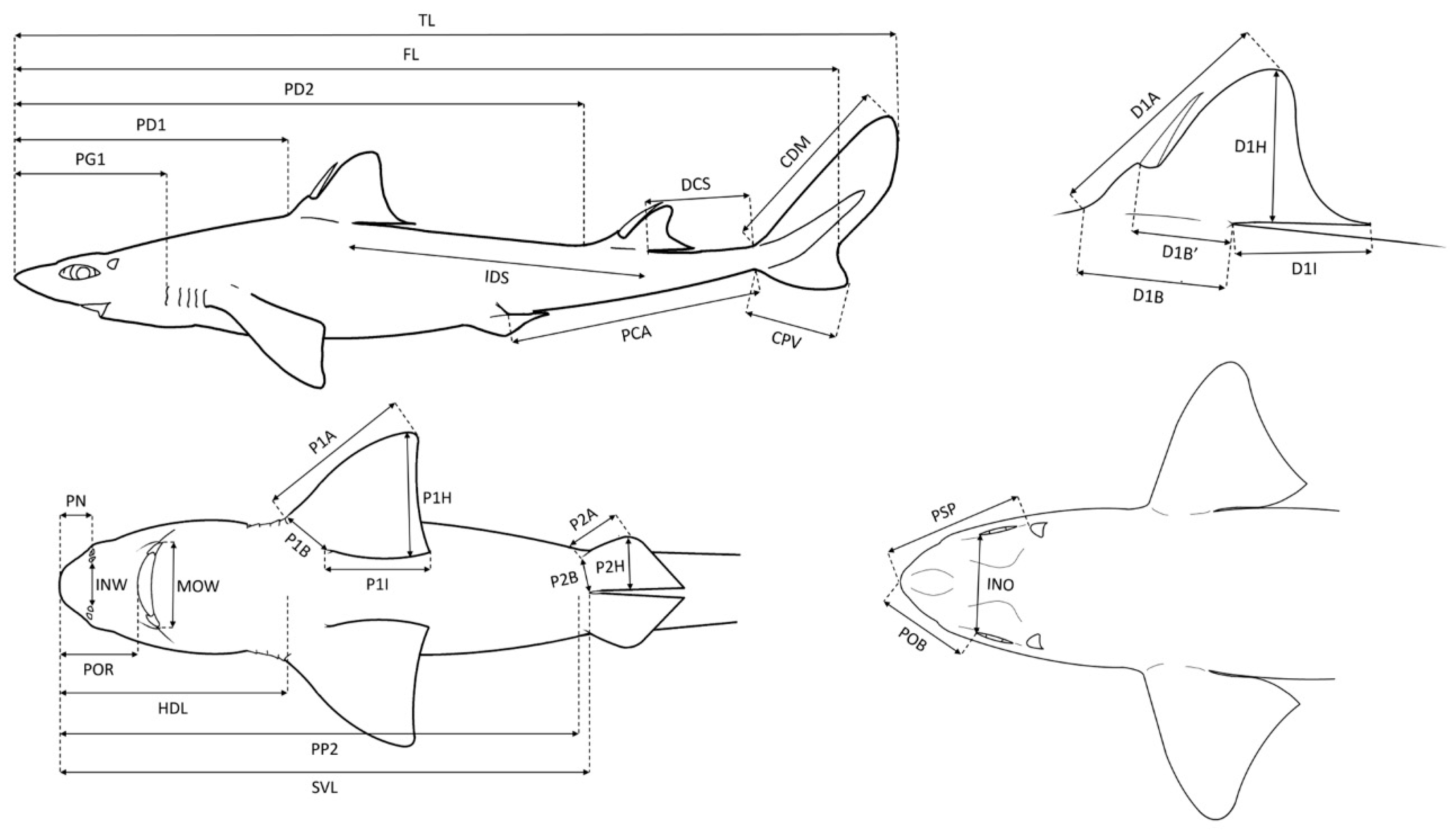
2.2. Body Morphometric Analysis
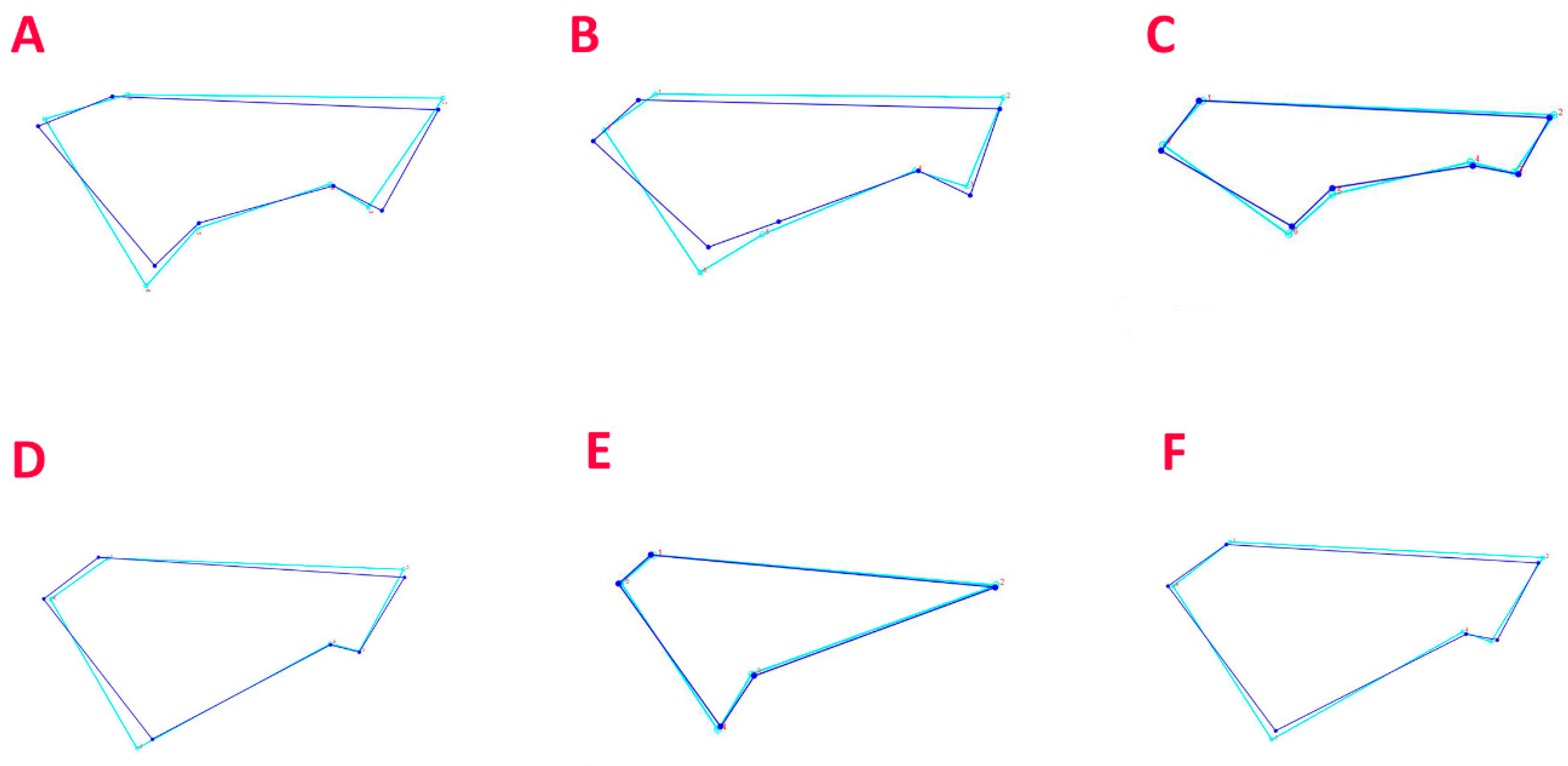
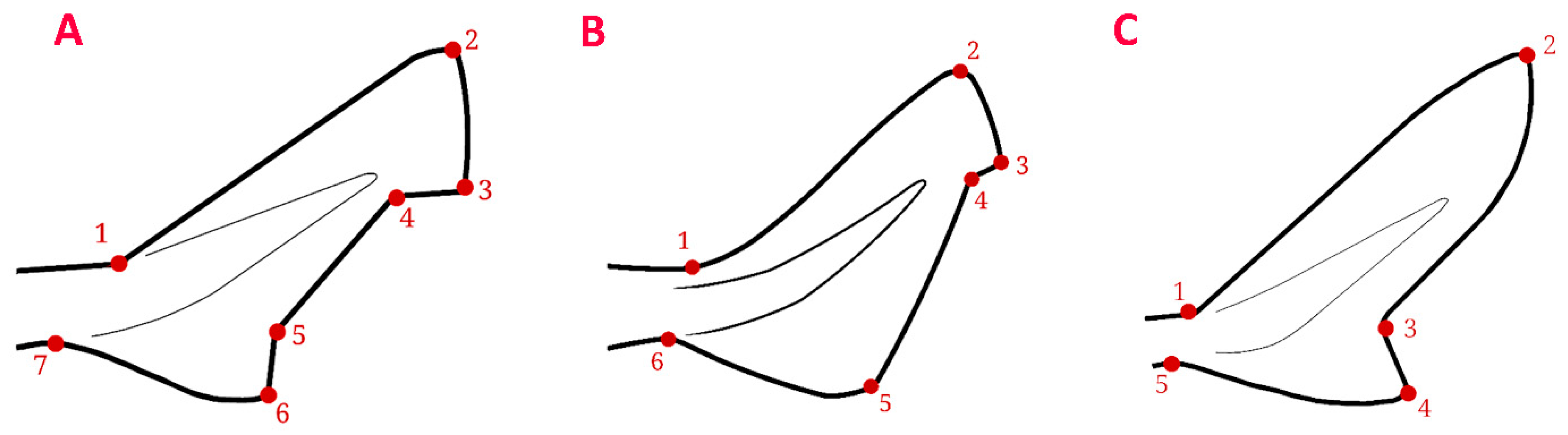
2.3. Geometric Morphometric Analysis of the Caudal Fin
3. Results
3.1. Linear Morphometric Analysis
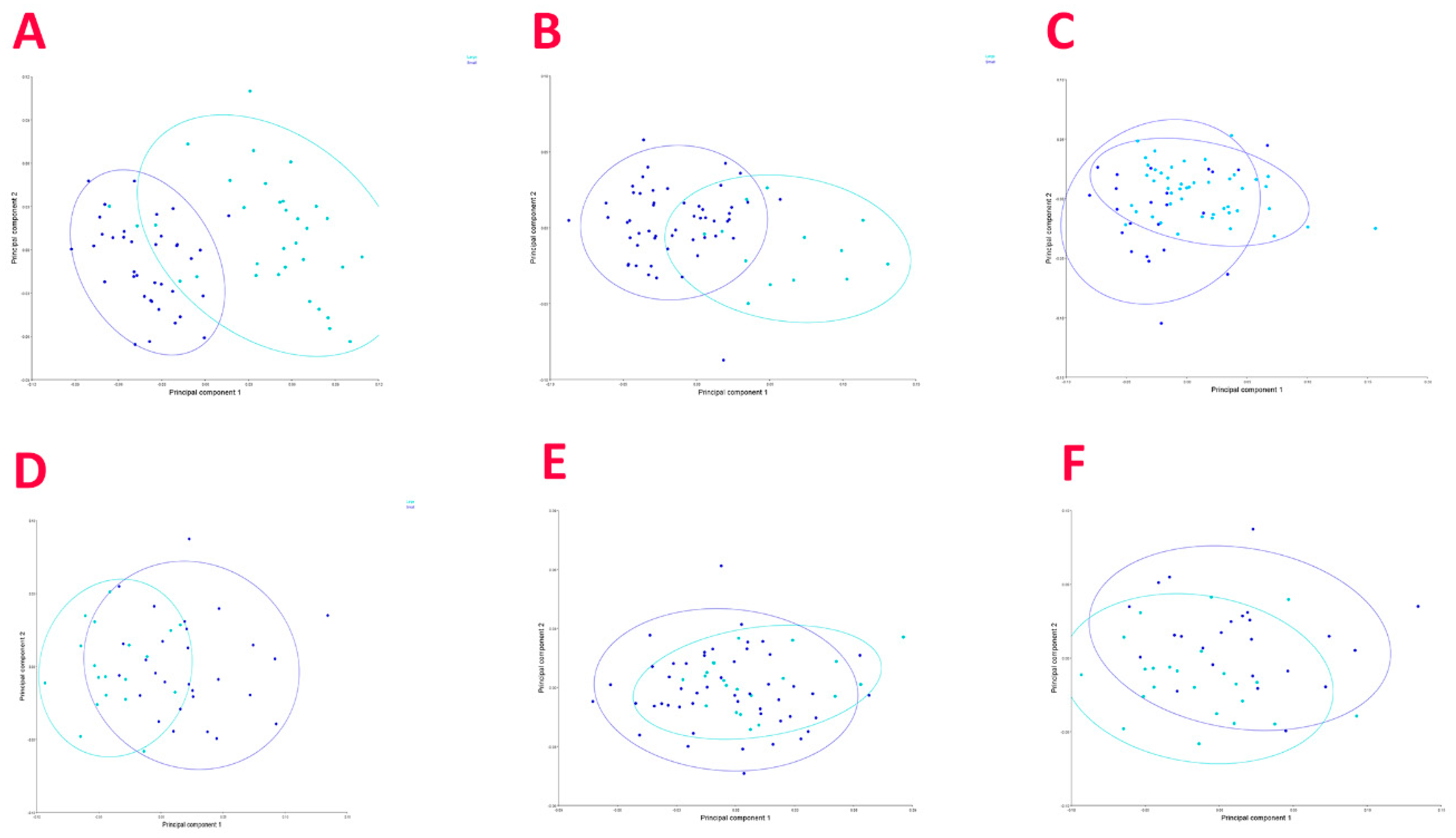
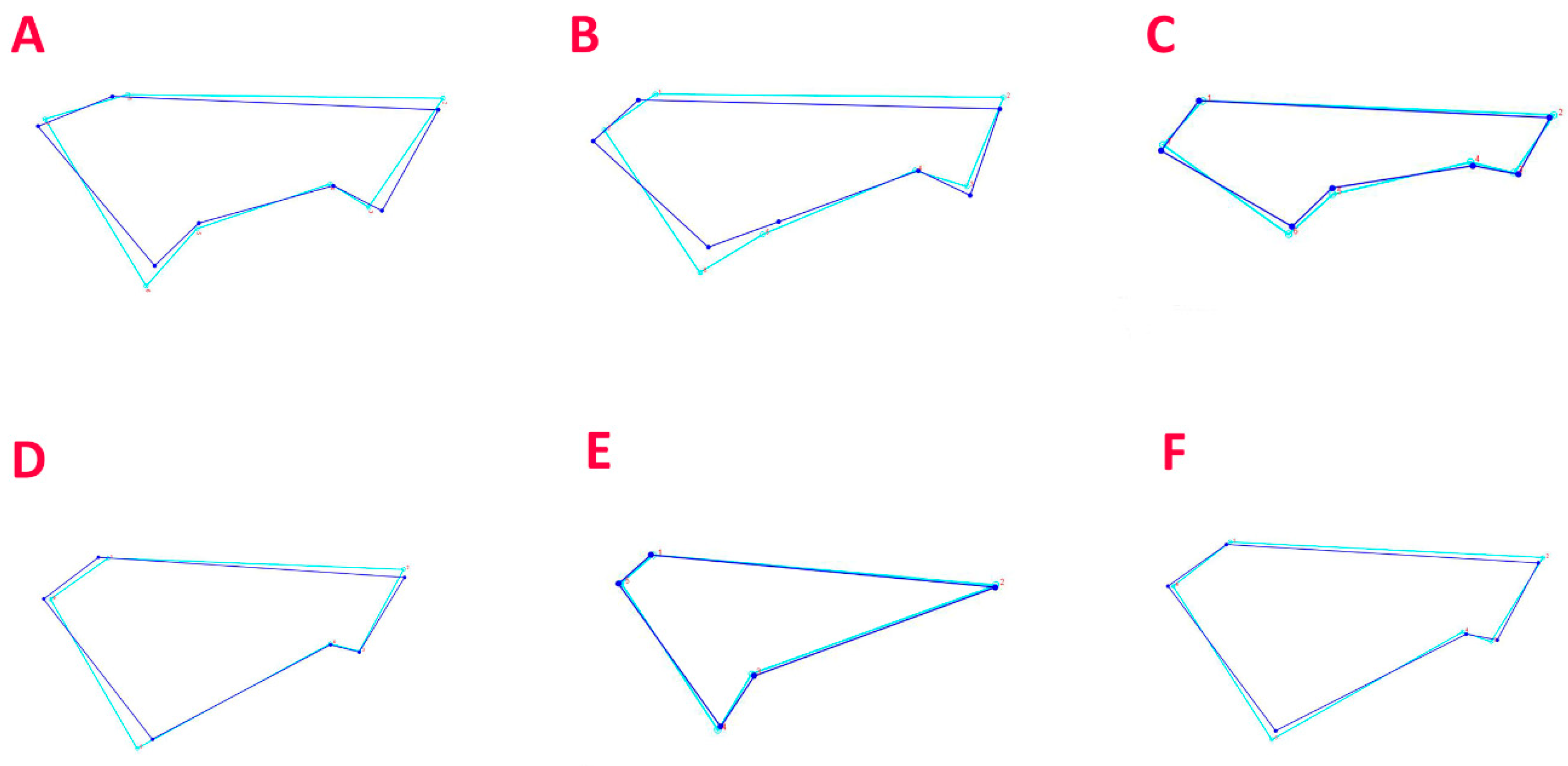
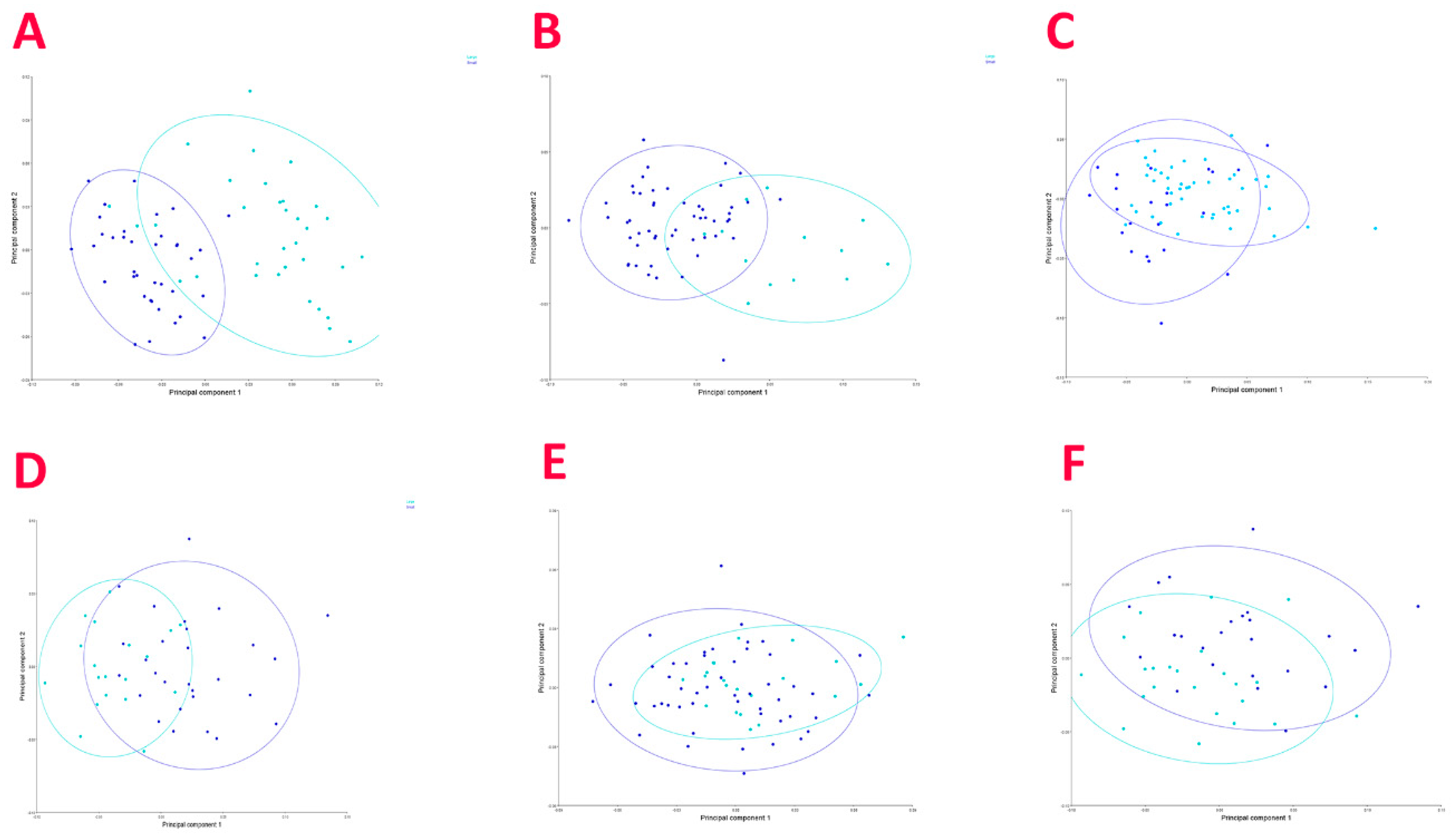
3.2. Geometric Morphometric Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Stevens, J.D.; Bonfil, R.; Dulvy, N.K.; Walker, P. The effects of fishing on sharks, rays and chimaeras (Chondrichthyans), and the implications for marine ecosystems. ICES J. Mar. Sci. 2000, 57, 476–494. [Google Scholar] [CrossRef]
- Mulas, A.; Bellodi, A.; Cannas, R.; Carbonara, P.; Cau, A.; Marongiu, M.F.; Pesci, P.; Porcu, C.; Follesa, M.C. Resource partitioning among sympatric elasmobranchs in the central-western Mediterranean continental shelf. Mar. Biol. 2019, 166, 153. [Google Scholar] [CrossRef]
- Bellodi, A.; Mulas, A.; Carbonara, P.; Cau, A.; Cuccu, D.; Marongiu, M.F.; Mura, V.; Pesci, P.; Zupa, W.; Porcu, C.; et al. New insights into life–history traits of Mediterranean Electric rays (Torpediniformes: Torpedinidae) as a contribution to their conservation. Zoology 2021, 146, 125922. [Google Scholar] [CrossRef] [PubMed]
- Ferretti, F.; Worm, B.; Britten, G.L.; Heithaus, M.R.; Lotze, H.K. Patterns and ecosystem consequences of shark declines in the ocean. Ecol. Lett. 2010, 13, 1055–1071. [Google Scholar] [CrossRef]
- Heithaus, M.R.; Wirsing, A.J.; Dill, L.M. The ecological importance of intact top-predator populations: A synthesis of 15 years of research in a seagrass ecosystem. Mar. Freshw. Res. 2012, 63, 1039–1050. [Google Scholar] [CrossRef]
- Irschick, D.J.; Hammerschlag, N. Morphological scaling of body form in four shark species differing in ecology and life history. Biol. J. Linn. Soc. 2014, 114, 126–135. [Google Scholar] [CrossRef]
- Fu, A.L.; Hammerschlag, N.; Lauder, G.V.; Wilga, C.D.; Kuo, C.Y.; Irschick, D.J. Ontogeny of head and caudal fin shape of an apex marine predator: The tiger shark (Galeocerdo cuvier). J. Morphol. 2016, 277, 556–564. [Google Scholar] [CrossRef]
- Dahl, R.B.; Sigsgaard, E.E.; Mwangi, G.; Thomsen, P.F.; Jørgensen, R.D.; de Oliveira Torquato, F.; Olsen, L.; Møller, P.R. The Sandy Zebra Shark: A New Color Morph of the Zebra Shark Stegostoma tigrinum, with a Redescription of the Species and a Revision of Its Nomenclature. Copeia 2019, 107, 524–541. [Google Scholar] [CrossRef]
- Ahnelt, H.; Sauberer, M.; Ramler, D.; Koch, L.; Pogoreutz, C. Negative allometric growth during ontogeny in the large pelagic filter-feeding basking shark. Zoomorphology 2020, 139, 71–83. [Google Scholar] [CrossRef]
- Carbonara, P.; Cannas, R.; Donnaloia, M.; Melis, R.; Porcu, C.; Spedicato, M.T.; Zupa, W.; Follesa, M.C. On the presence of Dipturus nidarosiensis (Storm, 1881) in the Central Mediterranean area. PeerJ 2019, 7, e7009. [Google Scholar] [CrossRef]
- Carbonara, P.; Bellodi, A.; Zupa, W.; Donnaloia, M.; Gaudio, P.; Neglia, C.; Follesa, M.C. Morphological Traits and Capture Depth of the Norwegian Skate (Dipturus nidarosiensis (Storm, 1881)) from Two Mediterranean Populations. J. Mar. Sci. Eng. 2021, 9, 1462. [Google Scholar] [CrossRef]
- Rodríguez Mendoza, R.P. Population Structure of the Bluemouth, Helicolenus dactylopterus (Teleostei: Sebastidae), in the Northeast Atlantic and Mediterranean Using Geometric Morphometric Techniques. Ph.D. Thesis, University of Vigo, Vigo, Spain, 2013. [Google Scholar]
- Lowry, D.; Motta, P.J.; Hueter, R.E. The ontogeny of feeding behavior and cranial morphology in the leopard shark Triakis semifasciata (Girard 1854): A longitudinal perspective. J. Exp. Mar. Biol. Ecol. 2007, 341, 153–167. [Google Scholar] [CrossRef]
- Reiss, K.L.; Bonnan, M.F. Ontogenetic Scaling of Caudal Fin Shape in Squalus acanthias (Chondrichthyes, Elasmobranchii): A Geometric Morphometric Analysis with Implications for Caudal Fin Functional Morphology. Anat. Rec. 2010, 293, 1184–1191. [Google Scholar] [CrossRef] [PubMed]
- Karachle, P.K.; Stergiou, K.I. Mouth allometry and feeding habits of some Mediterranean fishes. Acta Ichthyol. Piscat. 2011, 41, 265–275. [Google Scholar] [CrossRef]
- Roos, G.; Van Wassenbergh, S.; Aerts, P.; Herrel, A.; Adriaens, D. Effects of snout dimensions on the hydrodynamics of suction feeding in juvenile and adult seahorses. J. Theor. Biol. 2011, 269, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Lingham-Soliar, T. Caudal fin allometry in the white shark Carcharodon carcharias: Implications for locomotory performance and ecology. Naturwissenschaften 2005, 92, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Higham, T.E.; Seamone, S.G.; Arnold, A.; Toews, D.; Janmohamed, Z.; Smith, S.J.; Rogers, S.M. The ontogenetic scaling of form and function in the spotted ratfish, Hydrolagus colliei (Chondrichthyes: Chimaeriformes): Fins, muscles, and locomotion. J. Morphol. 2018, 279, 1408–1418. [Google Scholar] [CrossRef]
- Regan, M.C. Tails of the Sea: Morphological Scaling and Ontogeny of Shark Caudal Fins. Master’s Thesis, Wake Forest University, Winston-Salem, NC, USA, 2019. [Google Scholar]
- Filiz, H.; Taşkavak, E. Sexual dimorphism in the head, mouth, and body morphology of the smallspotted catshark, Scyliorhinus canicula (Linnaeus, 1758) (Chondrichthyes: Scyliorhinidae) from Turkey. Acta Adriat. 2006, 47, 37–47. [Google Scholar]
- Aka Erdogan, Z.; Torcu Koç, H.; Tûrker Çakir, D.; Nerlovic, V.; Dulcic, J. Sexual dimorphism in the small-spotted catshark, Scyliorhinus canicula (L.1758), from the edremit bay (Turkey). Acta Adriat. 2004, 47, 37–47. [Google Scholar]
- Marongiu, M.F.; Porcu, C.; Bellodi, A.; Cannas, R.; Cau, A.; Cuccu, D.; Mulas, A.; Follesa, M.C. Temporal dynamics of demersal chondrichthyan species in the central western Mediterranean Sea: The case study in Sardinia Island. Fish. Res. 2017, 193, 81–94. [Google Scholar] [CrossRef]
- Follesa, M.C.; Marongiu, M.F.; Zupa, W.; Bellodi, A.; Cau, A.; Cannas, R.; Colloca, F.; Djurovic, M.; Isajlovic, I.; Jadaud, A.; et al. Spatial variability of Chondrichthyes in the northern Mediterranean. Sci. Mar. 2019, 83, 81–100. [Google Scholar] [CrossRef]
- Porcu, C.; Marongiu, M.F.; Olita, A.; Bellodi, A.; Cannas, R.; Carbonara, P.; Cau, A.; Mulas, A.; Pesci, P.; Follesa, M.C. The demersal bathyal fish assemblage of the Central-Western Mediterranean: Depth distribution, sexual maturation and reproduction. Deep-Sea Res. Part I Oceanogr. Res. 2020, 166, 103394. [Google Scholar] [CrossRef]
- Serena, F.; Abella, A.J.; Bargnesi, F.; Barone, M.; Colloca, F.; Ferretti, F.; Fiorentino, F.; Jenrette, J.; Moro, S. Species diversity, taxonomy and distribution of Chondrichthyes in the Mediterranean and Black Sea. Eur. Zool. J. 2020, 87, 497–536. [Google Scholar] [CrossRef]
- Mulas, A.; Bellodi, A.; Carbonara, P.; Cau, A.; Marongiu, M.F.; Pesci, P.; Porcu, C.; Follesa, M.C. Bio-Ecological Features Update on Eleven Rare Cartilaginous Fish in the Central-Western Mediterranean Sea as a Contribution for Their Conservation. Life 2021, 11, 871. [Google Scholar] [CrossRef]
- Spedicato, M.T.; Massutí, E.; Mérigot, B.; Tserpes, G.; Jadaud, A.; Relini, G. The MEDITS trawl survey specifications in an ecosystem approach to fishery management. Sci. Mar. 2019, 83, 9–20. [Google Scholar] [CrossRef]
- Follesa, M.C.; Carbonara, P. Atlas of the Maturity Stages of Mediterranean Fishery Resources; Studies and Reviews n. 99; Food and Agriculture Organisation of United Nations: Rome, Italy, 2019; 268p. [Google Scholar]
- Follesa, M.C.; Agus, B.; Bellodi, A.; Cannas, R.; Capezzuto, F.; Casciaro, L.; Cau, A.; Cuccu, D.; Donnaloia, M.; Fernandez-Arcaya, U.; et al. The MEDITS maturity scales as a useful tool for investigating the reproductive traits of key species in the Mediterranean Sea. Sci. Mar. 2019, 83, 235–256. [Google Scholar] [CrossRef]
- Compagno, L.J.V. Sharks of the World. An Annotated and Illustrated Catalogue of Shark Species Known to Date. Volume 2. Bullhead, Mackerel and Carpet Sharks (Heterodontiformes, Lamniformes and Orectolobiformes); Report No. 1; Food and Agriculture Organisation of United Nations: Rome, Italy, 2001; Volume 2, 269p. [Google Scholar]
- Last, P.R.; White, W.T.; Pogonoski, J.J.; Gledhill, D.C.; Yearsley, G.K.; Ward, B. Application of a rapid taxonomic approach to genus Squalus. In Description of New Dogfishes of the Genus Squalus (Squaloidea: Squalidae); Last, P.R., White, W.T., Pogonoski, J.J., Eds.; CSIRO Marine and Atmospheric Research: Tasmania, Australia, 2007; pp. 1–10. [Google Scholar]
- Bellodi, A.; Porcu, C.; Cau, A.; Marongiu, M.F.; Melis, R.; Mulas, A.; Pesci, P.; Follesa, M.C.; Cannas, R. Investigation on the genus Squalus in the Sardinian waters (Central-Western Mediterranean) with implications on its management. Medit. Mar. Sci. 2018, 19, 256–272. [Google Scholar] [CrossRef]
- Bellodi, A.; Benvenuto, A.; Melis, R.; Mulas, A.; Barone, M.; Barría, C.; Cariani, A.; Carugati, L.; Chatzispyrou, A.; Desrochers, M.; et al. Call me by my name: Unravelling the taxonomy of the gulper shark genus Centrophorus in the Mediterranean Sea through an integrated taxonomic approach. Zool. J. Linn. Soc. 2022, 195, 815–840. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N. PRIMER V7: User Manual/Tutorial; PRIMER-E: Plymouth, MA, USA, 2015. [Google Scholar]
- Rohlf, F.J. The Tps series of software. Hystrix 2005, 26, 9–12. [Google Scholar]
- Klingenberg, C.P. MorphoJ: An integrated software package for geometric morphometrics. Mol. Ecol. Resour. 2011, 11, 353–357. [Google Scholar] [CrossRef]
- Nordell, S.E. Observations of the mating behavior and dentition of the round stingray, Urolophus halleri. Environ. Biol. Fish. 1994, 39, 219–229. [Google Scholar] [CrossRef]
- Conrath, C.L. Reproductive biology. In Management Techniques for Elasmobranch Fisheries; Musick, J.A., Bonfil, R., Eds.; Food and Agriculture Organisation of United Nations: Rome, Italy, 2005; pp. 103–126. [Google Scholar]
- Porcu, C.; Marongiu, M.F.; Follesa, M.C.; Bellodi, A.; Mulas, A.; Pesci, P.; Cau, A. Reproductive aspects of the velvet belly Etmopterus spinax (Chondrichthyes: Etmopteridae), from the central western Mediterranean Sea. Notes on gametogenesis and oviducal gland microstructure. Medit. Mar. Sci. 2014, 15, 313–326. [Google Scholar] [CrossRef]
- Marongiu, M.F.; Porcu, C.; Pascale, N.; Bellodi, A.; Cau, A.; Mulas, A.; Pesci, P.; Porceddu, R.; Follesa, M.C. A Taxonomic Survey of Female Oviducal Glands in Chondrichthyes: A Comparative Overview of Microanatomy in the Two Reproductive Modes. Animals 2021, 11, 2653. [Google Scholar] [CrossRef] [PubMed]
- Higham, T.E. The integration of locomotion and prey capture in vertebrates: Morphology, behaviour, and performance. Integr. Comp. Biol. 2007, 47, 82–95. [Google Scholar] [CrossRef] [PubMed]
- Webb, P.W. Body Form, Locomotion and Foraging in Aquatic Vertebrates. Am. Zool. 1984, 24, 107–120. [Google Scholar] [CrossRef]
- Scacco, U.; La Mesa, G.; Vacchi, M. Body morphometrics, swimming diversity and niche in demersal sharks: A comparative case study from the Mediterranean Sea. Sci. Mar. 2010, 74, 37–53. [Google Scholar] [CrossRef]
- Odum, W.E.; Heald, E.J. The detritus-based food web of an estuarine mangrove community. In Estuarine Research; Cronin, L.E., Ed.; Academic Press: New York, NY, USA, 1975; Volume 1, pp. 265–286. [Google Scholar]
- Bottaro, M.; Sinopoli, M.; Bertocci, I.; Follesa, M.C.; Cau, A.; Consalvo, I.; Scarcelli, F.; Sperone, E.; Vacchi, M.; Marsili, L.; et al. Jaws from the deep: Biological and ecological insights on the kitefin shark Dalatias licha from the Mediterranean Sea. Front. Mar. Sci. 2023, 10, 1155731. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Acronym | Description | Acronym | Description |
|---|---|---|---|
| TL | Total length | MOW | Mouth width |
| FL | Fork length | D1A | First dorsal fin anterior margin |
| PD1 | Pre-first dorsal fin length | D1B | First dorsal fin base length |
| PD2 | Pre-second dorsal fin length | D1BI | First dorsal fin spine to inner margin |
| PG1 | Prebranchial length | DIH | First dorsal fin height |
| IDS | Interdorsal space | D1I | First dorsal fin inner margin |
| DCS | Dorsal caudal fin space | D2A | Second dorsal fin anterior margin |
| CDM | Dorsal caudal fin margin | D2B | Second dorsal fin base length |
| PCA | Pelvic fin caudal fin space | D2BI | Second dorsal fin spine to inner margin |
| CPV | Preventral caudal fin margin | D2H | Second dorsal fin height |
| HDL | Head length | D2I | Second dorsal fin inner margin |
| PP2 | Prepelvic fin length | P1A | Pectoral fin anterior margin |
| SVL | Snout-vent length | P1B | Pectoral fin base |
| POB | Preorbital length | P1H | Pectoral fin height |
| INO | Interorbital length | P1I | Pectoral fin inner margin |
| PSP | Prespiracular length | P2A | Pelvic fin anterior margin |
| PN | Prenostril length | P2B | Pelvic fin base |
| POR | Preoral length | P2H | Pelvic fin height |
| INW | Internostril space |
| Sex | Size Group | Total | ||||
|---|---|---|---|---|---|---|
| Species | M | F | S | L | ||
| D. licha | Number | 46 | 57 | 81 | 22 | 103 |
| Size range | 30.64–87.39 | 30.27–109.44 | 30.27–58.39 | 68.8–109.44 | 30.27–109.44 | |
| Mean TL | 45.72 | 49.45 | 36.43 | 87.27 | 47.29 | |
| Standard deviation | 18.77 | 24.64 | 4.24 | 11.48 | 21.90 | |
| C. uyato | Number | 55 | 40 | 48 | 47 | 95 |
| Size range | 37.95–89.84 | 40.42–104.00 | 37.95–53.97 | 55.90–104.00 | 37.95–104 | |
| Mean TL | 62.93 | 65.41 | 46.45 | 81.87 | 63.97 | |
| Standard deviation | 17.08 | 22.76 | 4.01 | 10.99 | 19.60 | |
| S. blainville | Number | 51 | 29 | 56 | 24 | 80 |
| Size range | 28.7–59.50 | 22.00–79.20 | 22.00–39.90 | 40.50–79.20 | 22.00–79.20 | |
| Mean TL | 39.39 | 41.69 | 34.91 | 52.62 | 40.22 | |
| Standard deviation | 7.88 | 13.48 | 3.33 | 10.23 | 10.24 | |
| E. spinax | Number | 50 | 74 | 60 | 64 | 124 |
| Size range | 11.99–31.62 | 10.61–42.30 | 10.61–24.83 | 25.12–42.30 | 10.61–42.3 | |
| Mean TL | 22.39 | 25.39 | 17.55 | 30.40 | 24.18 | |
| Standard deviation | 6.13 | 8.57 | 4.02 | 4.72 | 7.79 | |
| O. centrina | Number | 22 | 25 | 28 | 19 | 47 |
| Size range | 22.97–63.15 | 24.57–74.61 | 22.97–54.48 | 55.02–74.61 | 22.97–74.61 | |
| Mean TL | 48.61 | 57.01 | 44.82 | 65.25 | 53.08 | |
| Standard deviation | 11.80 | 13.92 | 10.56 | 6.07 | 13.51 | |
| Sex as Factor | ||||||||
|---|---|---|---|---|---|---|---|---|
| Computed Group | ||||||||
| Species | Correlation | Corr. Sq. | Misclassification Error (%): | Orig. Group | F | M | Total | %Correct |
| Centrophorus uyato | 0.5744 | 0.3299 | 36.842 | F | 27 | 13 | 40 | 67.500 |
| M | 22 | 33 | 55 | 60.000 | ||||
| Dalatias licha | 0.6361 | 0.4047 | 39.806 | F | 35 | 22 | 57 | 61.404 |
| M | 19 | 27 | 46 | 58.696 | ||||
| Etmopterus spinax | 0.4129 | 0.1705 | 33.065 | F | 51 | 23 | 74 | 68.919 |
| M | 18 | 32 | 50 | 64.000 | ||||
| Oxynotus centrina | 0.4531 | 0.2053 | 34.043 | F | 18 | 7 | 25 | 72.000 |
| M | 9 | 13 | 22 | 59.091 | ||||
| Squalus blainville | 0.685 | 0.4692 | 25.000 | M | 40 | 11 | 51 | 78.431 |
| F | 9 | 20 | 29 | 68.966 | ||||
| Size as Factor | ||||||||
| Computed Group | ||||||||
| Correlation | Corr. Sq. | Misclassification Error (%): | Orig. Group | Large | Small | Total | %Correct | |
| Centrophorus uyato | 0.8459 | 0.7155 | 6.316 | Large | 42 | 5 | 47 | 89.362 |
| Small | 1 | 47 | 48 | 97.917 | ||||
| Dalatias licha | 0.894 | 0.7993 | 0.971 | large | 22 | 0 | 22 | 100.000 |
| small | 1 | 80 | 81 | 98.765 | ||||
| Etmopterus spinax | 0.7817 | 0.611 | 16.129 | Large | 55 | 9 | 64 | 85.938 |
| Small | 11 | 49 | 60 | 81.667 | ||||
| Oxynotus centrina | 0.8034 | 0.6454 | 12.766 | Large | 16 | 3 | 19 | 84.211 |
| Small | 3 | 25 | 28 | 89.286 | ||||
| Squalus blainville | 0.756 | 0.5716 | 12.500 | Small | 49 | 7 | 56 | 87.500 |
| Large | 3 | 21 | 24 | 87.500 | ||||
| Sex | Size Group | Total | ||||
|---|---|---|---|---|---|---|
| Species | M | F | S | L | ||
| D. licha | Number | 24 | 42 | 53 | 13 | 66 |
| Size range | 33.90–89.00 | 31.50–104.30 | 31.39–58.39 | 68.80–109.44 | 31.50–109.30 | |
| Mean TL | 42.61 | 51.46 | 36.56 | 88.71 | 47.51 | |
| Standard deviation | 16.55 | 25.17 | 4.13 | 13.97 | 22.06 | |
| C. uyato | Number | 45 | 25 | 37 | 33 | 70 |
| Size range | 41.58–89.84 | 38.70–104.00 | 38.70–54.30 | 55.90–104.00 | 38.70–104.00 | |
| Mean TL | 63.96 | 60.14 | 46.68 | 80.44 | 62.59 | |
| Standard deviation | 17.00 | 21.77 | 3.80 | 11.05 | 18.77 | |
| S. blainville | Number | 45 | 25 | 49 | 21 | 70 |
| Size range | 28.70–59.50 | 22.00–79.20 | 22.00–39.90 | 40.50–79.20 | 22.00–79.20 | |
| Mean TL | 39.80 | 41.66 | 34.92 | 53.40 | 40.47 | |
| Standard deviation | 8.22 | 13.99 | 3.47 | 10.32 | 10.59 | |
| E. spinax | Number | 27 | 43 | 23 | 47 | 70 |
| Size range | 13.20–39.00 | 11.60–43.5 | 11.60–24.60 | 25.5–43.5 | 11.60–43.50 | |
| Mean TL | 24.89 | 28.85 | 18.42 | 31.68 | 27.32 | |
| Standard deviation | 6.52 | 8.27 | 3.72 | 5.15 | 7.84 | |
| O. centrina | Number | 22 | 25 | 28 | 19 | 47 |
| Size range | 22.97–63.15 | 24.57–74.61 | 22.97–54.48 | 55.02–74.61 | 22.97–74.61 | |
| Mean TL | 48.61 | 57.01 | 44.82 | 65.25 | 53.08 | |
| Standard deviation | 11.80 | 13.92 | 10.56 | 6.07 | 13.51 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bellodi, A.; Mulas, A.; Daniel, L.; Cau, A.; Porcu, C.; Carbonara, P.; Follesa, M.C. Ontogenetic Shifts in Body Morphology of Demersal Sharks’ Species (Order: Squaliformes) Inhabiting the Western-Central Mediterranean Sea, with Implications for Their Bio-Ecological Role. Biology 2023, 12, 1150. https://doi.org/10.3390/biology12081150
Bellodi A, Mulas A, Daniel L, Cau A, Porcu C, Carbonara P, Follesa MC. Ontogenetic Shifts in Body Morphology of Demersal Sharks’ Species (Order: Squaliformes) Inhabiting the Western-Central Mediterranean Sea, with Implications for Their Bio-Ecological Role. Biology. 2023; 12(8):1150. https://doi.org/10.3390/biology12081150
Chicago/Turabian StyleBellodi, Andrea, Antonello Mulas, Louise Daniel, Alessandro Cau, Cristina Porcu, Pierluigi Carbonara, and Maria Cristina Follesa. 2023. "Ontogenetic Shifts in Body Morphology of Demersal Sharks’ Species (Order: Squaliformes) Inhabiting the Western-Central Mediterranean Sea, with Implications for Their Bio-Ecological Role" Biology 12, no. 8: 1150. https://doi.org/10.3390/biology12081150
APA StyleBellodi, A., Mulas, A., Daniel, L., Cau, A., Porcu, C., Carbonara, P., & Follesa, M. C. (2023). Ontogenetic Shifts in Body Morphology of Demersal Sharks’ Species (Order: Squaliformes) Inhabiting the Western-Central Mediterranean Sea, with Implications for Their Bio-Ecological Role. Biology, 12(8), 1150. https://doi.org/10.3390/biology12081150