

Harnessing Diesel-Degrading Potential of an Antarctic Microalga from Greenwich Island and Its Physiological Adaptation

 , ,
, ,  ,
, 
 ,
,  , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Microalgal Culture
2.2. Growth Characterisation
2.3. Polyphasic Strain Identification and Characterisation of Surface Properties
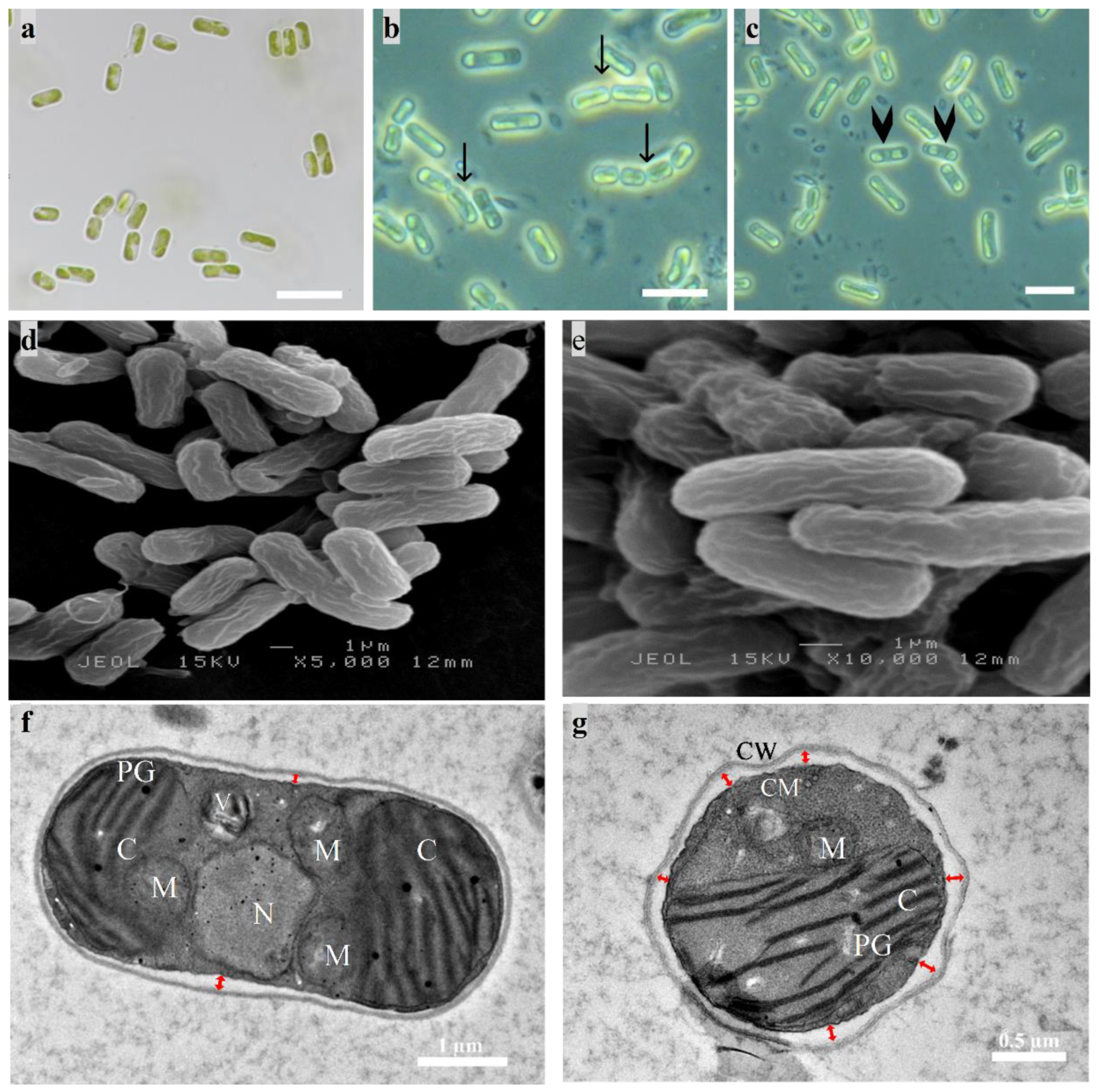
2.3.1. Morphological Identification
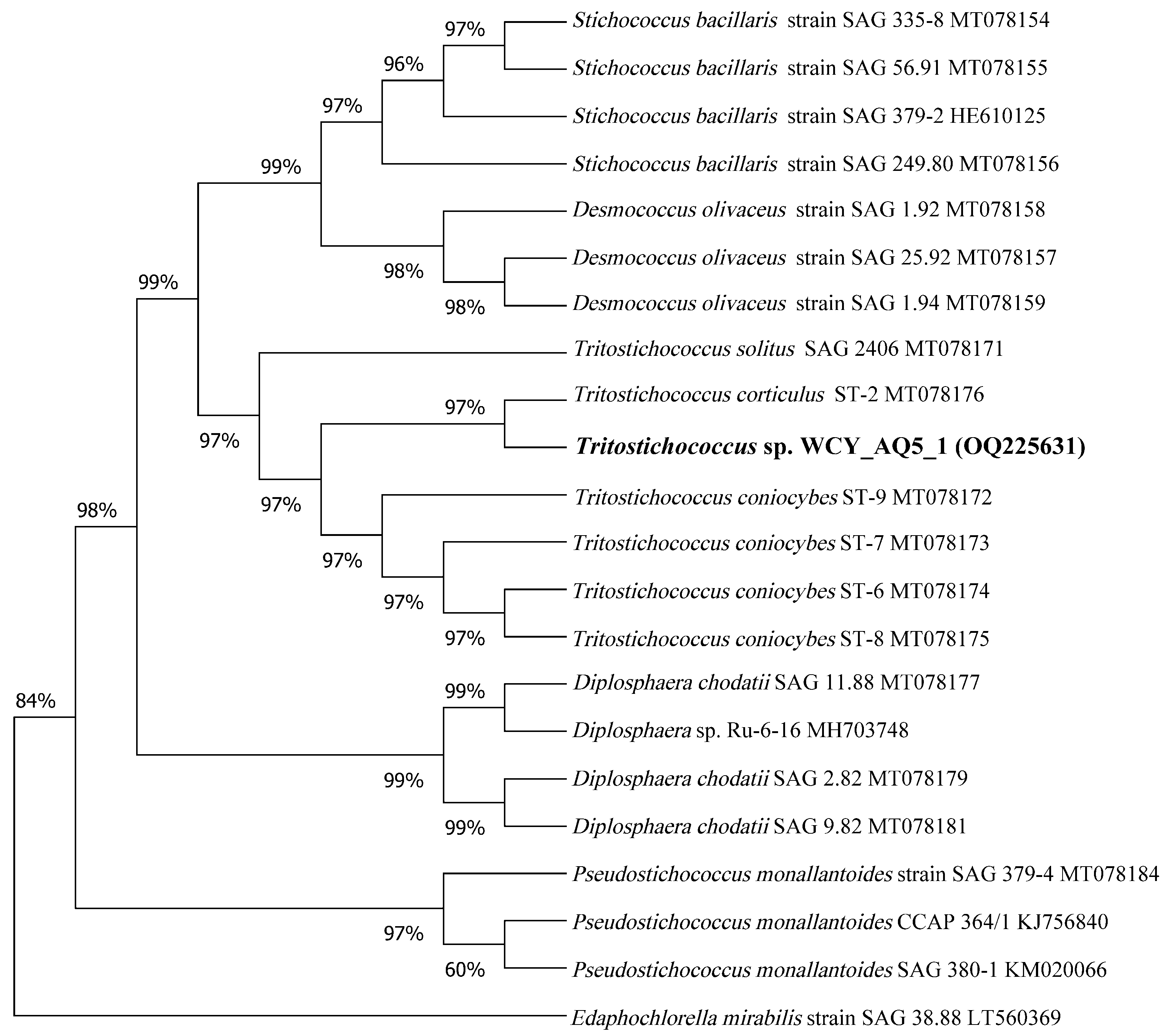
2.3.2. Molecular Identification Using a Dual Barcode Approach
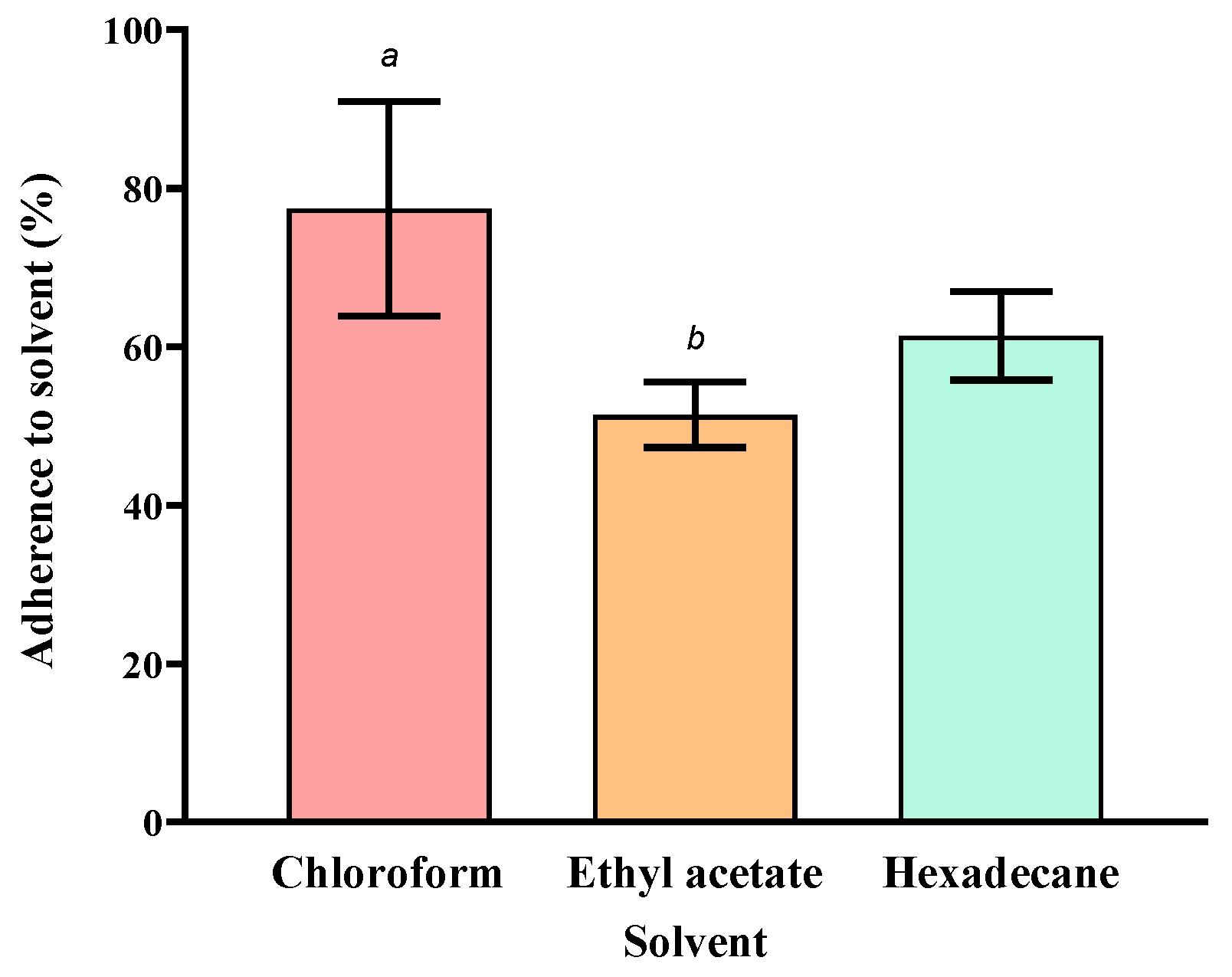
2.3.3. Characterisation of Cell Surface Properties
2.4. Phytoremediation of Diesel Hydrocarbons
2.5. Quantitative Assessment of Microalgal Biodegradation Efficiency
2.5.1. Gravimetric Analysis
2.5.2. Chromatographic Analysis
2.6. Statistical Analyses
3. Results
3.1. Polyphasic Identification
3.2. Surface Property Characterisation
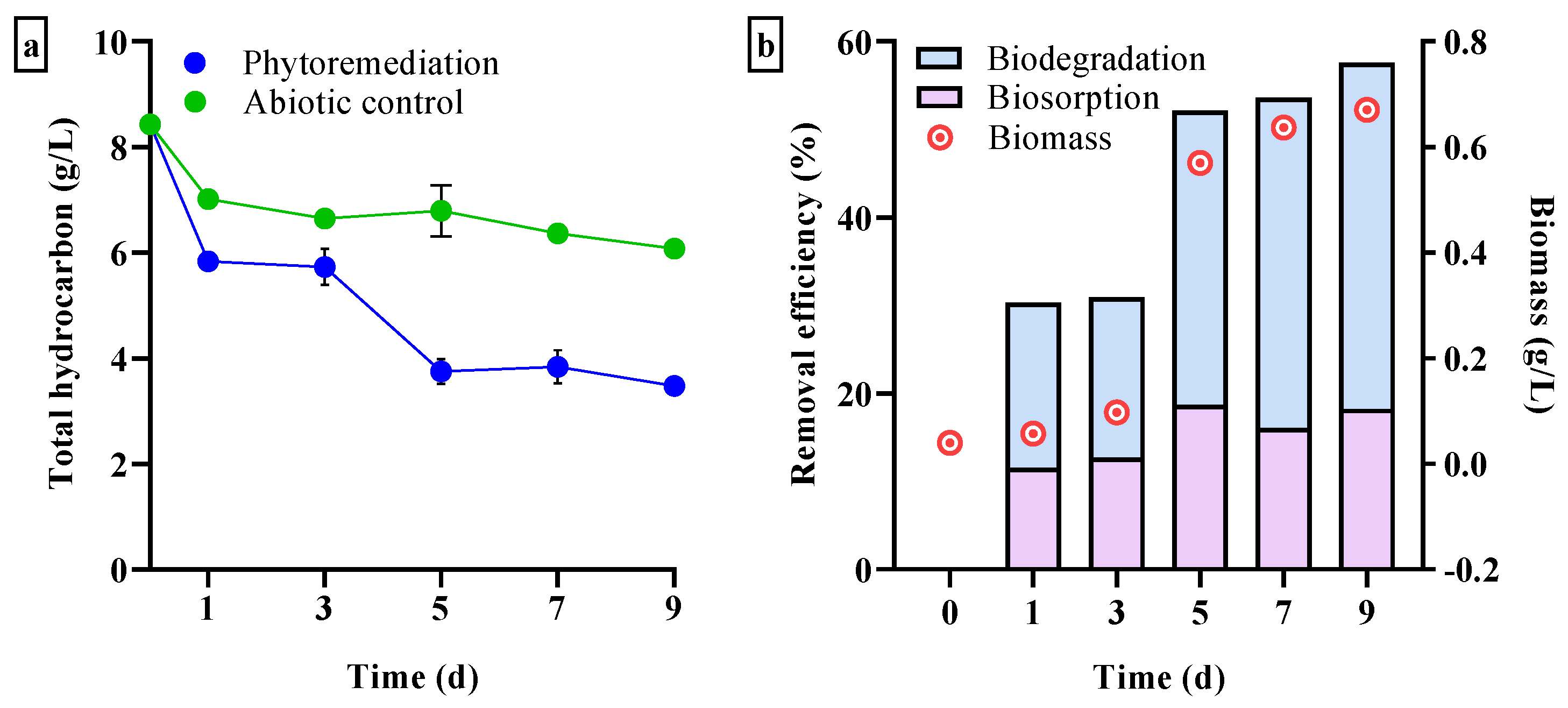
3.3. Efficiency of Phytoremediation of Diesel
3.3.1. Total Hydrocarbon Loss
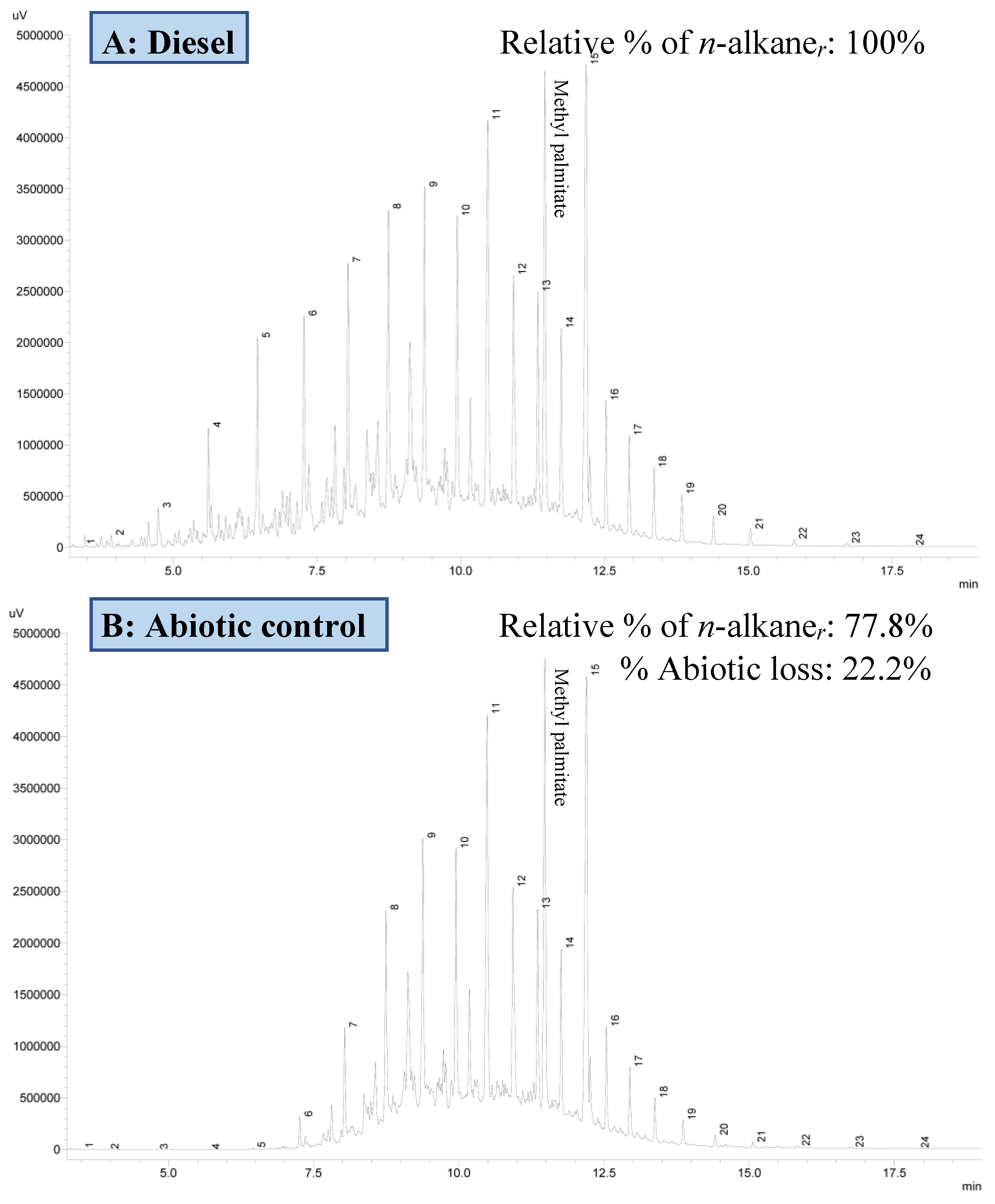
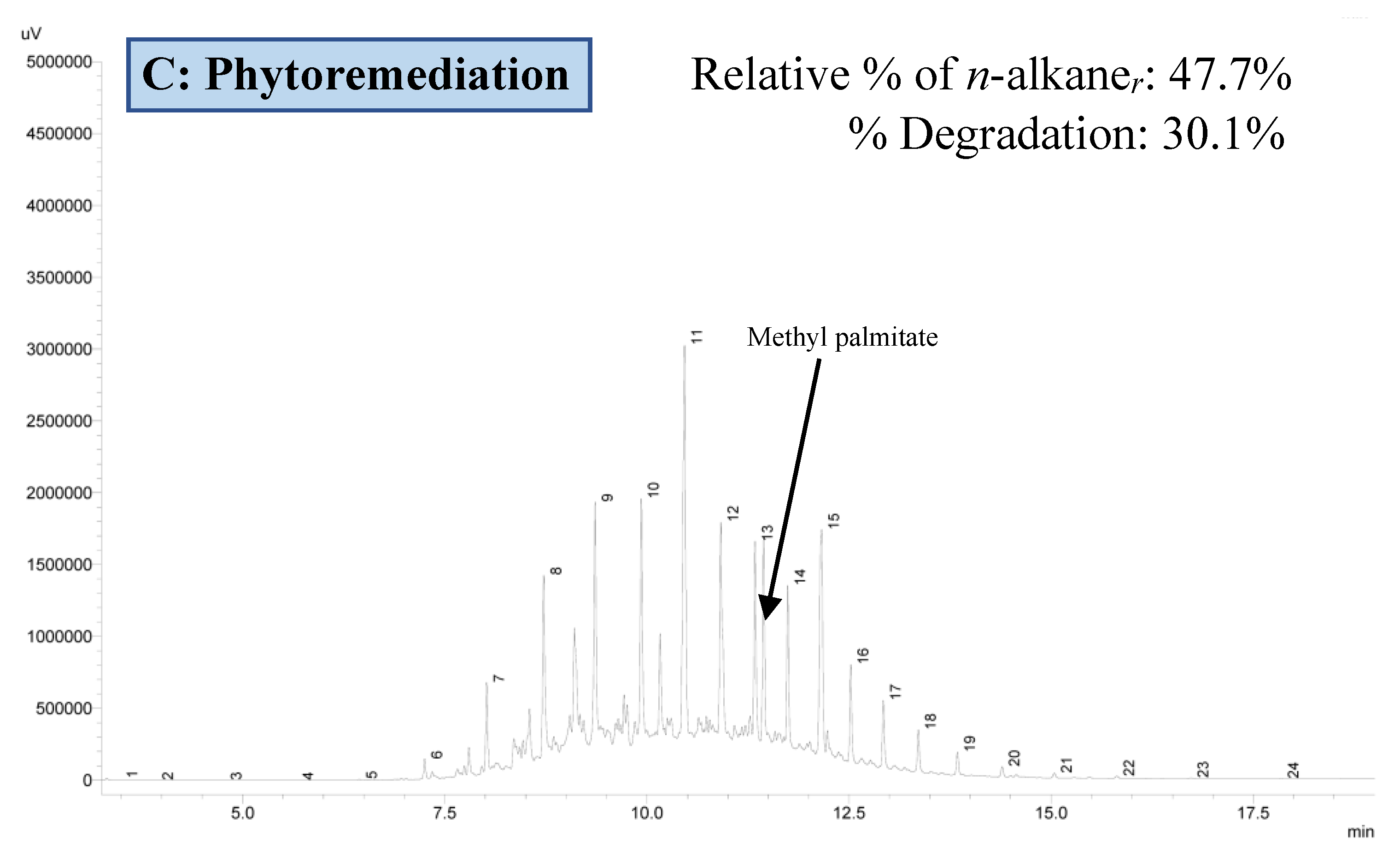
3.3.2. Residual Hydrocarbon Profile
3.4. Physiological Effects on Microalgal Cells Exposed to Diesel
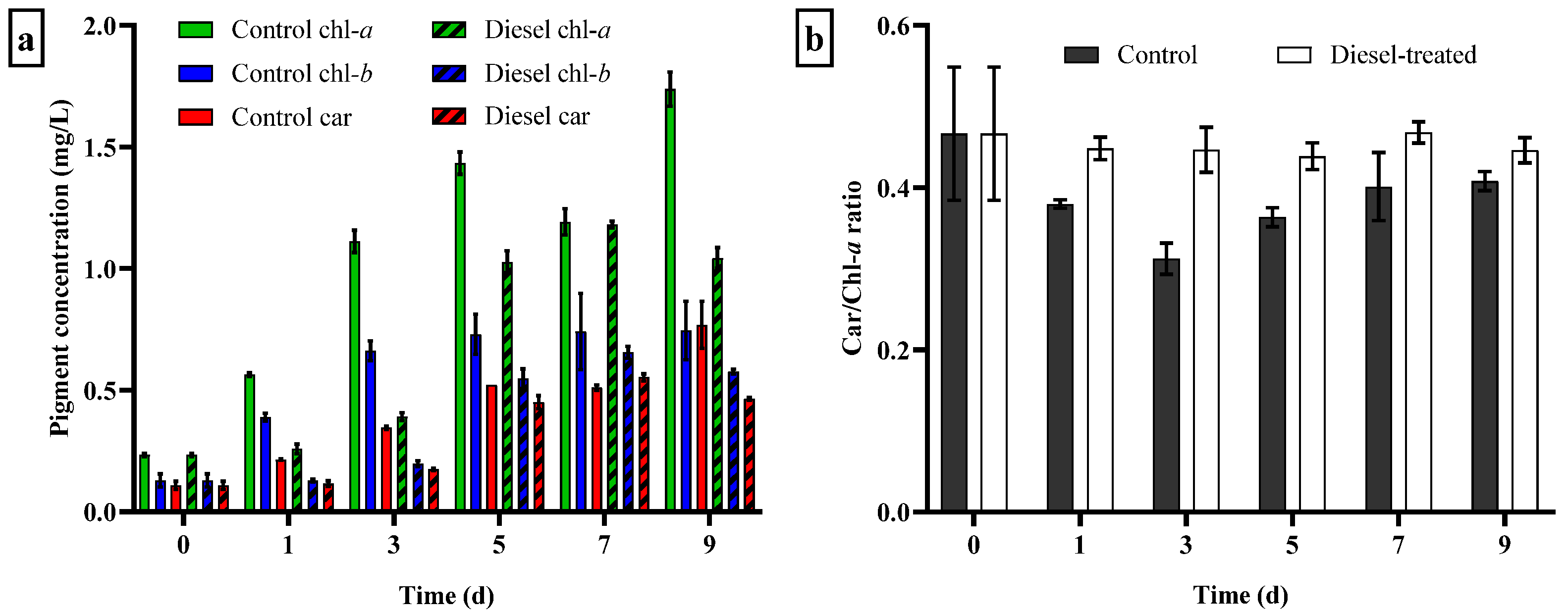
3.4.1. Growth Response and Changes in Photosynthetic Pigment Content
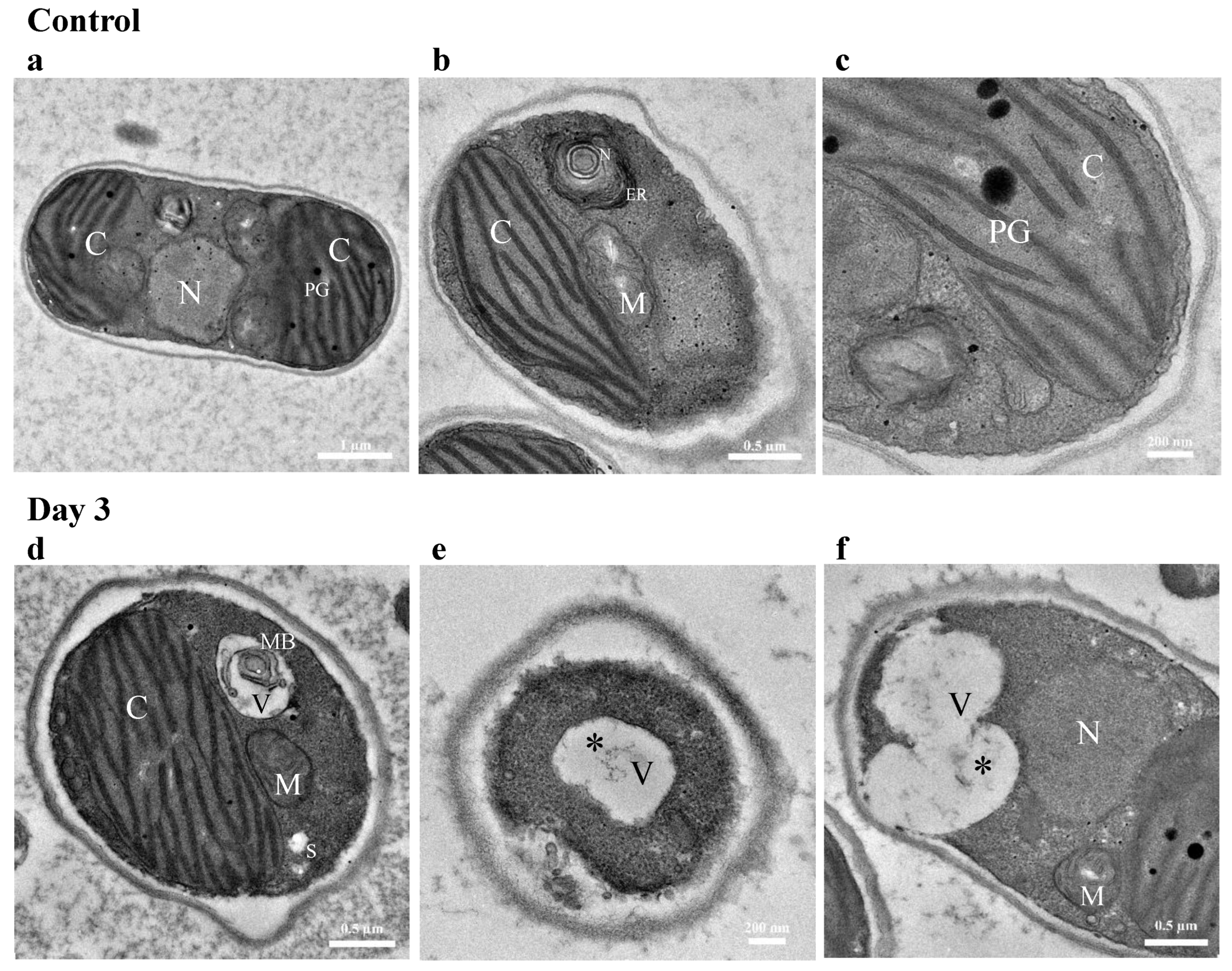
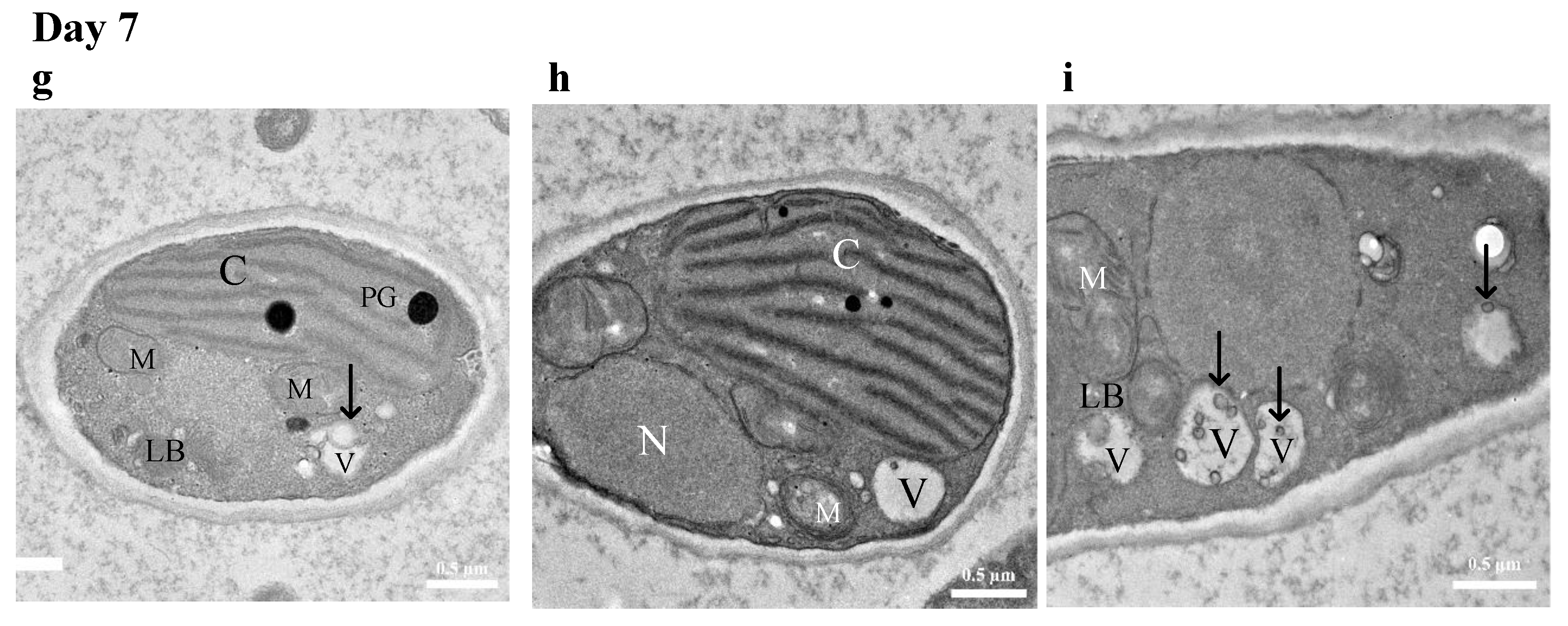
3.4.2. TEM Imaging of Cellular Response to Diesel Exposure
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McMinn, A.; Pankowskii, A.; Ashworth, C.; Bhagooli, R.; Ralph, P.; Ryan, K. In situ net primary productivity and photosynthesis of Antarctic sea ice algal, phytoplankton and benthic algal communities. Mar. Biol. 2010, 157, 1345–1356. [Google Scholar] [CrossRef]
- Van Leeuwe, M.A.; Tedesco, L.; Arrigo, K.R.; Assmy, P.; Campbell, K.; Meiners, K.M.; Rintala, J.M.; Selz, V.; Thomas, D.N.; Stefels, J. Microalgal community structure and primary production in Arctic and Antarctic sea ice: A synthesis. Elementa 2018, 6. [Google Scholar] [CrossRef]
- Laraib, N.; Hussain, A.; Javid, A.; Hafeez-ur-Rehman, M.; Bukhari, S.M.; Rashid, M.; Ali, W. Recent Advancements in Microalgal-Induced Remediation of Wastewaters; INC: New York, NY, USA, 2020. [Google Scholar] [CrossRef]
- Lim, Z.S.; Wong, R.R.; Wong, C.-Y.; Zulkharnain, A.; Shaharuddin, N.A.; Ahmad, S.A. Bibliometric analysis of research on diesel pollution in Antarctica and a review on remediation techniques. Appl. Sci. 2021, 11, 1123. [Google Scholar] [CrossRef]
- Sweet, S.T.; Kennicutt, M.C.; Klein, A.G. The grounding of the Bahía Paraíso, Arthur Harbor, Antarctica. In Handbook of Oil Spill Science and Technology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015; pp. 547–556. [Google Scholar] [CrossRef]
- McWatters, R.S.; Rowe, R.K.; Wilkins, D.; Spedding, T.; Hince, G.; Richardson, J.; Snape, I. Modelling of vapour intrusion into a building impacted by a fuel spill in Antarctica. J. Environ. Manag. 2019, 231, 467–482. [Google Scholar] [CrossRef] [PubMed]
- Brown, K.E. Impacts of Diesel and Residual Fuel Oil on Antarctic Marine Invertebrates; Southern Cross University: Lismore, Australia, 2017. [Google Scholar]
- Ruoppolo, V.; Woehler, E.J.; Morgan, K.; Clumpner, C.J. Wildlife and oil in the Antarctic: A recipe for cold disaster. Polar Rec. 2013, 49, 97–109. [Google Scholar] [CrossRef]
- Roslee, A.F.A.; Zakaria, N.N.; Convey, P.; Zulkharnain, A.; Lee, G.L.Y.; Gomez-Fuentes, C.; Ahmad, S.A. Statistical Optimisation of Growth Conditions and Diesel Degradation by the Antarctic bacterium, Rhodococcus Sp. Strain AQ5–07. Extremophiles 2020, 24, 277–291. [Google Scholar] [CrossRef]
- van Dorst, J.; Wilkins, D.; Crane, S.; Montgomery, K.; Zhang, E.; Spedding, T.; Hince, G.; Ferrari, B. Microbial Community analysis of biopiles in Antarctica provides evidence of successful hydrocarbon biodegradation and initial soil ecosystem recovery. Environ. Pollut. 2021, 290, 117977. [Google Scholar] [CrossRef]
- Maßmig, M.; Lüdke, J.; Krahmann, G.; Engel, A. Bacterial degradation activity in the eastern tropical South Pacific oxygen minimum zone. Biogeosciences 2020, 17, 215–230. [Google Scholar] [CrossRef]
- Baghour, M. Algal degradation of organic pollutants. In Handbook of Ecomaterials; Springer International Publishing: Cham, Switzerland, 2019; Volume 1, pp. 565–586. [Google Scholar] [CrossRef]
- Das, B.; Deka, S. A cost-effective and environmentally sustainable process for phycoremediation of oil field formation water for its safe disposal and reuse. Sci. Rep. 2019, 9, 15232. [Google Scholar] [CrossRef]
- Lei, A.-P.; Wong, Y.-S.; Tam, N.F.-Y. Pyrene-induced changes of glutathione-s-transferase activities in different microalgal species. Chemosphere 2003, 50, 293–301. [Google Scholar] [CrossRef]
- Ke, L.; Luo, L.; Wang, P.; Luan, T.; Tam, N.F.Y. Effects of metals on biosorption and biodegradation of mixed polycyclic aromatic hydrocarbons by a freshwater green alga Selenastrum capricornutum. Bioresour. Technol. 2010, 101, 6950–6961. [Google Scholar] [CrossRef]
- Hong, Y.W.; Yuan, D.X.; Lin, Q.M.; Yang, T.L. Accumulation and biodegradation of phenanthrene and fluoranthene by the algae enriched from a mangrove aquatic ecosystem. Mar. Pollut. Bull. 2008, 56, 1400–1405. [Google Scholar] [CrossRef]
- Lei, A.-P.; Hu, Z.-L.; Wong, Y.-S.; Tam, N.F.-Y. Removal of fluoranthene and pyrene by different microalgal species. Bioresour. Technol. 2007, 98, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Ghasemi, Y.; Rasoul-Amini, S.; Fotooh-Abadi, E. The biotransformation, biodegradation, and bioremediation of organic compounds by microalgae. J. Phycol. 2011, 47, 969–980. [Google Scholar] [CrossRef]
- Nydahl, A.C.; King, C.K.; Wasley, J.; Jolley, D.F.; Robinson, S.A. Toxicity of fuel-contaminated soil to antarctic moss and terrestrial algae. Environ. Toxicol. Chem. 2015, 34, 2004–2012. [Google Scholar] [CrossRef] [PubMed]
- Young, J.N.; Schmidt, K. It’s what’s inside that matters: Physiological adaptations of high-latitude marine microalgae to environmental change. New Phytol. 2020, 227, 1307–1318. [Google Scholar] [CrossRef] [PubMed]
- Nichols, H.W. Growth media-freshwater. In Handbook of Phycological Methods: Culture Methods and Growth Measurements; Stein, J., Ed.; Cambridge University Press: Cambridge, UK, 1973; pp. 7–24. [Google Scholar]
- Lichtenthaler, H.K.; Buschmann, C. Chlorophylls and carotenoids: Measurement and characterization by UV-VIS spectroscopy. Curr. Protoc. Food Anal. Chem. 2001, 1, F4.3.1–F4.3.8. [Google Scholar] [CrossRef]
- Krzemińska, I.; Pawlik-Skowrońska, B.; Trzcińska, M.; Tys, J. Influence of photoperiods on the growth rate and biomass productivity of green microalgae. Bioprocess Biosyst. Eng. 2014, 37, 735–741. [Google Scholar] [CrossRef] [PubMed]
- John, D.; Brooks, A.J.; Whitton, B. The Freshwater Algal Flora of the British Isles: An Identification Guide to Freshwater and Terrestrial Algae; Cambridge University Press: Cambridge, UK, 2011. [Google Scholar]
- Pröschold, T.; Darienko, T. The green puzzle Stichococcus (Trebouxiophyceae, Chlorophyta): New generic and species concept among this widely distributed genus. Phytotaxa 2020, 441, 113–142. [Google Scholar] [CrossRef]
- Hodač, L.; Hallmann, C.; Spitzer, K.; Elster, J.; Faßhauer, F.; Brinkmann, N.; Lepka, D.; Diwan, V.; Friedl, T. Widespread green algae Chlorella and Stichococcus exhibit polar-temperate and tropical-temperate biogeography. FEMS Microbiol. Ecol. 2016, 92, fiw122. [Google Scholar] [CrossRef]
- Rosenberg, M.; Gutnick, D.; Rosenberg, E. Adherence of bacteria to hydrocarbons: A simple method for measuring cell-surface hydrophobicity. FEMS Microbiol. Lett. 1980, 9, 29–33. [Google Scholar] [CrossRef]
- Grace, C.E.E.; Lakshmi, P.K.; Meenakshi, S.; Vaidyanathan, S.; Srisudha, S.; Mary, M.B. Biomolecular transitions and lipid accumulation in green microalgae monitored by FTIR and Raman analysis. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2020, 224, 117382. [Google Scholar] [CrossRef] [PubMed]
- Murdock, J.N.; Wetzel, D.L. FT-IR Microspectroscopy enhances biological and ecological analysis of algae. Appl. Spectrosc. Rev. 2009, 44, 335–361. [Google Scholar] [CrossRef]
- Department of Standards Malaysia. MS123-3:2016 Malaysian Standard: Diesel Fuel—Specification—Part 3: Euro 5; Department of Standards Malaysia: Kuala Lumpur, Malaysia, 2016. [Google Scholar]
- Nägeli, C. Gattungen Einzelliger Algen Physiologisch und Systematisch Bearbeitet; Friedrich Schulthess: Zürich, Switzerland, 1849. [Google Scholar]
- Syuhada, N.H.; Merican, F.; Zaki, S.; Broady, P.A.; Convey, P.; Muangmai, N. Strong and widespread cycloheximide resistance in Stichococcus-like eukaryotic algal taxa. Sci. Rep. 2022, 12, 1080. [Google Scholar] [CrossRef] [PubMed]
- Van, A.T.; Glaser, K. Pseudostichococcus stands out from its siblings due to high salinity and desiccation tolerance. Phycology 2022, 2, 108–119. [Google Scholar] [CrossRef]
- Almomani, F.; Bohsale, R.R. Bio-sorption of toxic metals from industrial wastewater by algae strains Spirulina platensis and Chlorella vulgaris: Application of isotherm, kinetic models and process optimization. Sci. Total Environ. 2021, 755, 142654. [Google Scholar] [CrossRef]
- Gupta, V.K.; Shrivastava, A.K.; Jain, N. Biosorption of chromium(vi) from aqueous solutions by green algae Spirogyra species. Water Res. 2001, 35, 4079–4085. [Google Scholar] [CrossRef]
- Sultana, N.; Hossain, S.M.Z.; Mohammed, M.E.; Irfan, M.F.; Haq, B.; Faruque, M.O.; Razzak, S.A.; Hossain, M.M. Experimental study and parameters optimization of microalgae based heavy metals removal process using a hybrid response surface methodology-crow search algorithm. Sci. Rep. 2020, 10, 15068. [Google Scholar] [CrossRef]
- Hadjoudja, S.; Deluchat, V.; Baudu, M. Cell surface characterisation of Microcystis aeruginosa and Chlorella vulgaris. J. Colloid Interface Sci. 2010, 342, 293–299. [Google Scholar] [CrossRef]
- Xia, L.; Li, H.; Song, S. Cell surface characterization of some oleaginous green algae. J. Appl. Phycol. 2016, 28, 2323–2332. [Google Scholar] [CrossRef]
- Obuekwe, C.O.; Al-Jadi, Z.K.; Al-Saleh, E.S. Hydrocarbon degradation in relation to cell-surface hydrophobicity among bacterial hydrocarbon degraders from petroleum-contaminated Kuwait desert environment. Int. Biodeterior. Biodegrad. 2009, 63, 273–279. [Google Scholar] [CrossRef]
- Zhao, Q.; Zhang, J.-Y.; Chen, L.-Z.; Zheng, J.-X.; Zhao, L.; Yin, H.-M. Cell-surface hydrophobicity and degradation characteristics of hydrophobic hydrocarbon degrading bacteria. Huan Jing Ke Xue Huanjing Kexue 2005, 26, 132–136. [Google Scholar] [PubMed]
- Qasim, M.; Ansari, T.M.; Hussain, M. Combustion, performance, and emission evaluation of a diesel engine with biodiesel like fuel blends derived from a mixture of Pakistani waste canola and waste transformer oils. Energies 2017, 10, 1023. [Google Scholar] [CrossRef]
- de Llasera, M.P.G.; de Olmos-Espejel, J.J.; Díaz-Flores, G.; Montaño-Montiel, A. Biodegradation of benzo(a)pyrene by two freshwater microalgae Selenastrum capricornutum and Scenedesmus acutus: A comparative study useful for bioremediation. Environ. Sci. Pollut. Res. 2016, 23, 3365–3375. [Google Scholar] [CrossRef]
- Souza, L.S.; Simioni, C.; Bouzon, Z.L.; Schneider, R.C.S.; Gressler, P.; Miotto, M.C.; Rossi, M.J.; Rörig, L.R. Morphological and ultrastructural characterization of the acidophilic and lipid-producer strain Chlamydomonas acidophila LAFIC-004 (Chlorophyta) under different culture conditions. Protoplasma 2017, 254, 1385–1398. [Google Scholar] [CrossRef]
- Young, E.B.; Reed, L.; Berges, J.A. Growth parameters and responses of green algae across a gradient of phototrophic, mixotrophic and heterotrophic conditions. PeerJ 2022, 10, e13776. [Google Scholar] [CrossRef]
- Walker, J.D.; Colwell, R.R.; Vaituzis, Z.; Meyer, S.A. Petroleum-degrading achlorophyllous alga Prototheca zopfii. Nature 1975, 254, 423–424. [Google Scholar] [CrossRef]
- Kalhor, A.X.; Movafeghi, A.; Mohammadi-Nassab, A.D.; Abedi, E.; Bahrami, A. Potential of the green alga Chlorella vulgaris for biodegradation of crude oil hydrocarbons. Mar. Pollut. Bull. 2017, 123, 286–290. [Google Scholar] [CrossRef]
- El-Sheekh, M.M.; Hamouda, R.A.; Nizam, A.A. Biodegradation of crude oil by Scenedesmus obliquus and Chlorella vulgaris growing under heterotrophic conditions. Int. Biodeterior. Biodegrad. 2013, 82, 67–72. [Google Scholar] [CrossRef]
- Okoh, A. Biodegradation alternative in the cleanup of petroleum. Biotechnol. Mol. Biol. 2006, 1, 38–50. [Google Scholar]
- Subashchandrabose, S.R.; Logeshwaran, P.; Venkateswarlu, K.; Naidu, R.; Megharaj, M. Pyrene degradation by Chlorella sp. MM3 in liquid medium and soil slurry: Possible role of dihydrolipoamide acetyltransferase in pyrene biodegradation. Algal Res. 2017, 23, 223–232. [Google Scholar] [CrossRef]
- Tossavainen, M.; Ilyass, U.; Ollilainen, V.; Valkonen, K.; Ojala, A.; Romantschuk, M. Influence of long term nitrogen limitation on lipid, protein and pigment production of Euglena gracilis in photoheterotrophic cultures. PeerJ 2019, 7, e6624. [Google Scholar] [CrossRef] [PubMed]
- Cheirsilp, B.; Torpee, S. Enhanced growth and lipid production of microalgae under mixotrophic culture condition: Effect of light intensity, glucose concentration and fed-batch cultivation. Bioresour. Technol. 2012, 110, 510–516. [Google Scholar] [CrossRef] [PubMed]
- Shi, T.Q.; Wang, L.R.; Zhang, Z.X.; Sun, X.M.; Huang, H. Stresses as first-line tools for enhancing lipid and carotenoid production in microalgae. Front. Bioeng. Biotechnol. 2020, 8, 610. [Google Scholar] [CrossRef]
- Solovchenko, A.E.; Khozin-Goldberg, I.; Didi-Cohen, S.; Cohen, Z.; Merzlyak, M.N. Effects of light and nitrogen starvation on the content and composition of carotenoids of the green microalga Parietochloris incisa. Russ. J. Plant Physiol. 2008, 55, 455–462. [Google Scholar] [CrossRef]
- Kokabi, K.; Gorelova, O.; Zorin, B.; Didi-Cohen, S.; Itkin, M.; Malitsky, S.; Solovchenko, A.; Boussiba, S.; Khozin-Goldberg, I. Lipidome remodeling and autophagic respose in the arachidonic-acid-rich microalga Lobosphaera incisa under nitrogen and phosphorous deprivation. Front. Plant Sci. 2020, 11, 614846. [Google Scholar] [CrossRef]
- Roncaglia, B.; Papini, A.; Chini Zittelli, G.; Rodolfi, L.; Tredici, M.R. Cell wall and organelle modifications during nitrogen starvation in Nannochloropsis oceanica F&M-M24. J. Appl. Phycol. 2021, 33, 2069–2080. [Google Scholar] [CrossRef]
- Shebanova, A.; Ismagulova, T.; Solovchenko, A.; Baulina, O.; Lobakova, E.; Ivanova, A.; Moiseenko, A.; Shaitan, K.; Polshakov, V.; Nedbal, L.; et al. Versatility of the green microalga cell vacuole function as revealed by analytical transmission electron microscopy. Protoplasma 2017, 254, 1323–1340. [Google Scholar] [CrossRef]
- Sanz-Luque, E.; Bhaya, D.; Grossman, A.R. Polyphosphate: A multifunctional metabolite in cyanobacteria and algae. Front. Plant Sci. 2020, 11, 938. [Google Scholar] [CrossRef]
- Nguyen, H.C.; Su, C.H.; Yu, Y.K.; Huong, D.T.M. Sugarcane bagasse as a novel carbon source for heterotrophic cultivation of oleaginous microalga Schizochytrium sp. Ind. Crops Prod. 2018, 121, 99–105. [Google Scholar] [CrossRef]
- Sajadian, S.; Morowvat, M.; Ghasemi, Y. Investigation of autotrophic, heterotrophic, and mixotrophic modes of cultivation on lipid and biomass production in Chlorella vulgaris. Natl. J. Physiol. Pharm. Pharmacol. 2018, 8, 594–599. [Google Scholar] [CrossRef]
- Zhang, T.Y.; Wu, Y.H.; Wang, J.H.; Wang, X.X.; Deantes-Espinosa, V.M.; Dao, G.H.; Tong, X.; Hu, H.Y. Heterotrophic cultivation of microalgae in straw lignocellulose hydrolysate for production of high-value biomass rich in polyunsaturated fatty acids (PUFA). Chem. Eng. J. 2019, 367, 37–44. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Specifications | |
|---|---|
| Oven program | Initial: 40 °C Ramp: 20 °C/min until 200 °C then 30 °C/min until 300 °C (10 min hold time) |
| Injection mode | Split (10:1 ratio) |
| Carrier gas | Helium |
| Gas flow rate | Helium: 30 mL/min Hydrogen: 40 mL/min Compressed air: 400 mL/min |
| Column flow rate | 0.9 mL/min |
| Pressure | 73.9 kPa |
| Back detector (FID) | 310 °C |
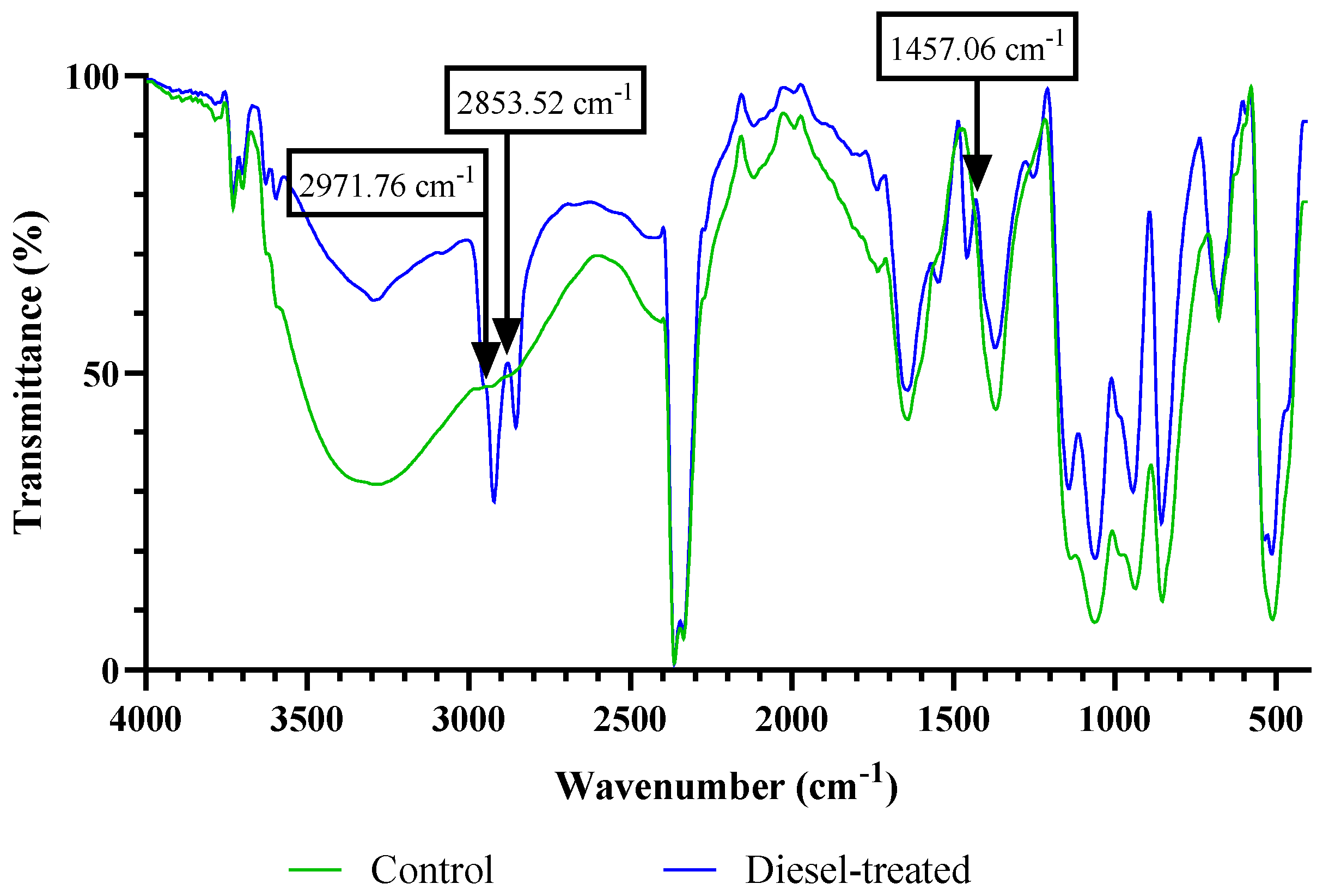
| Main Peaks (cm−1) | Tentative Band Assignment | Biomolecule | Wavenumber Range (cm−1) | |
|---|---|---|---|---|
| Control | Diesel-Treated | |||
| 3276.99 | 3294.07 | ν O-H/ν N-H | Water, protein | 3639–3029 |
| - | 2971.76, 2853.52 | νas CH2, ν CH2, ν CH3 | Lipid, proteins | 2970–2850 |
| 1640.04 | 1639.95 | ν C=O of amide I | Proteins | 1655–1638 |
| - | 1457.06 | δasCH3, δasCH2 | Lipid, proteins | 1456–1450 |
| 1368.67 | 1370.58 | Δ CH3 | Lipid, proteins | 1398–1370 |
| 1061.31 | 1141.70, 1060.49 | P=O | Phospholipids, nucleic acids | 1200–900 |
| 936.75, 853.09 | 943.12, 854.69 | ν (C-O-C) of polysaccharides | Nucleotides, carbohydrates | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, Z.S.; Wong, C.-Y.; Ahmad, S.A.; Puasa, N.A.; Phang, L.Y.; Shaharuddin, N.A.; Merican, F.; Convey, P.; Zulkharnain, A.; Shaari, H.; et al. Harnessing Diesel-Degrading Potential of an Antarctic Microalga from Greenwich Island and Its Physiological Adaptation. Biology 2023, 12, 1142. https://doi.org/10.3390/biology12081142
Lim ZS, Wong C-Y, Ahmad SA, Puasa NA, Phang LY, Shaharuddin NA, Merican F, Convey P, Zulkharnain A, Shaari H, et al. Harnessing Diesel-Degrading Potential of an Antarctic Microalga from Greenwich Island and Its Physiological Adaptation. Biology. 2023; 12(8):1142. https://doi.org/10.3390/biology12081142
Chicago/Turabian StyleLim, Zheng Syuen, Chiew-Yen Wong, Siti Aqlima Ahmad, Nurul Aini Puasa, Lai Yee Phang, Noor Azmi Shaharuddin, Faradina Merican, Peter Convey, Azham Zulkharnain, Hasrizal Shaari, and et al. 2023. "Harnessing Diesel-Degrading Potential of an Antarctic Microalga from Greenwich Island and Its Physiological Adaptation" Biology 12, no. 8: 1142. https://doi.org/10.3390/biology12081142
APA StyleLim, Z. S., Wong, C.-Y., Ahmad, S. A., Puasa, N. A., Phang, L. Y., Shaharuddin, N. A., Merican, F., Convey, P., Zulkharnain, A., Shaari, H., Azmi, A. A., Kok, Y.-Y., & Gomez-Fuentes, C. (2023). Harnessing Diesel-Degrading Potential of an Antarctic Microalga from Greenwich Island and Its Physiological Adaptation. Biology, 12(8), 1142. https://doi.org/10.3390/biology12081142