In Vitro Antiprotozoal Activity of Hibiscus sabdariffa Extract against a Ciliate Causing High Mortalities in Turbot Aquaculture

 ,
,  , and
, and
Abstract
Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Isolation and Culture of Parasites
2.2. Preparation of H. sabdariffa Extract
2.3. Analysis of H. sabdariffa Extract Bioactivity
2.3.1. Effect on Parasite Population Growth
2.3.2. Oxidative Stress Biomarkers
2.3.3. Proteolytic Activity
2.3.4. Expression Analysis of Protease-Encoding Genes by qRT-PCR
2.4. Chemical Analysis of the Crude Extract
2.4.1. Ultra-High-Performance Liquid Chromatography (UHPLC) Analysis
2.4.2. Electrospray Ionization Mass Spectrometric Detection (ESI–MSn) Analysis
2.5. Statistical Analysis
3. Results
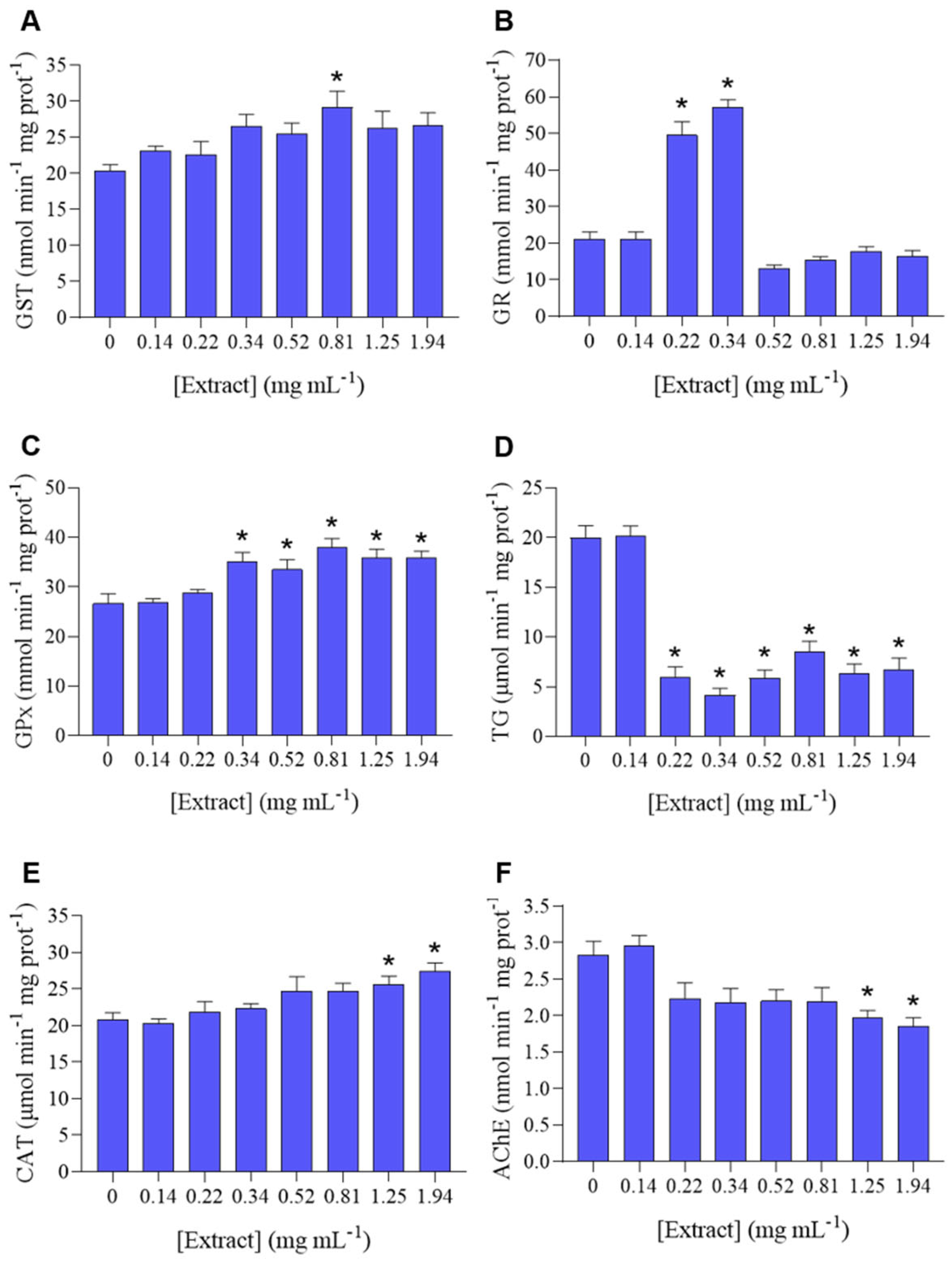
3.1. Effect of Hibiscus Extract on P. dicentrarchi Biological Responses
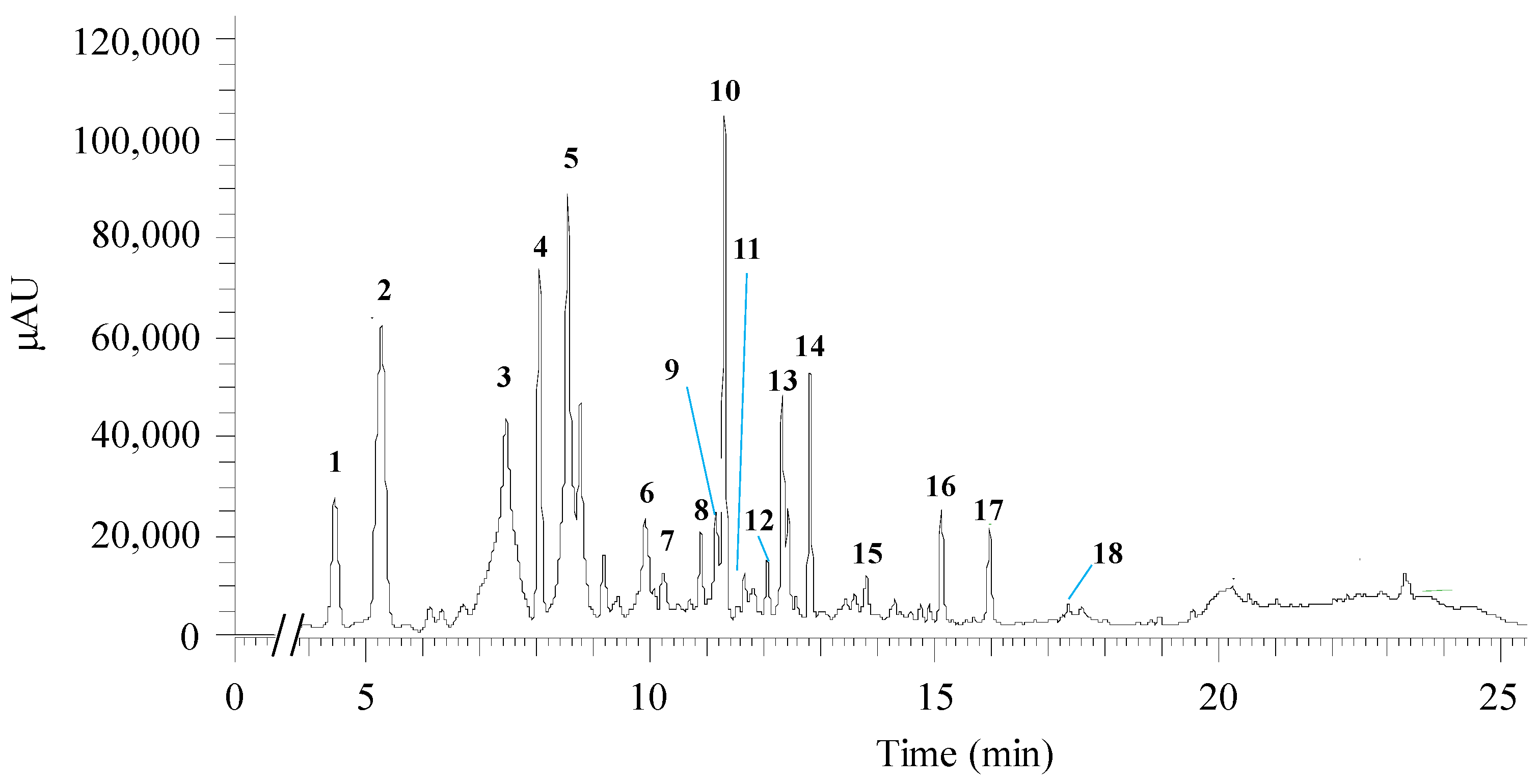
3.2. Qualitative Chemical Analysis of Hibiscus Extract
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kim, S.M.; Cho, J.B.; Kim, S.K.; Nam, Y.K.; Kim, K.H. Occurrence of scuticociliatosis in olive flounder Paralichthys olivaceus by Phiasterides dicentrarchi (Ciliophora: Scuticociliatida). Dis. Aquat. Org. 2004, 62, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Budiño, B.; Leiro, J.; Cabaleiro, S.; Lamas, J. Characterization of Philasterides dicentrarchi isolates that are pathogenic to turbot: Serology and cross-protective immunity. Aquaculture 2012, 364–365, 130–136. [Google Scholar] [CrossRef]
- Paramá, A.; Iglesias, R.; Álvarez, M.F.; Leiro, J.; Ubeira, F.M.; Sanmartín, M.L. Cysteine proteinase activities in the fish pathogen Philasterides dicentrarchi (Ciliophora: Scuticociliatida). Parasitology 2004, 128, 541–548. [Google Scholar] [CrossRef]
- Shin, S.P.; Han, S.Y.; Han, J.E.; Jun, J.W.; Kim, J.H.; Park, S.C. Expression and characterization of cathepsin L-like cysteine protease from Philasterides dicentrarchi. Parasitol. Int. 2014, 63, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Paramá, A.; Castro, R.; Lamas, J.; Sanmartín, M.L.; Santamarina, M.T.; Leiro, J. Scuticociliate proteinases may modulate turbot immune response by inducing apoptosis in pronephric leucocytes. Int. J. Parasitol. 2007, 37, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Dzik, J.M. Molecules released by helminth parasites involved in host colonization. Acta Biochim. Pol. 2006, 53, 33–64. [Google Scholar] [CrossRef]
- McKerrow, J.H.; Caffrey, C.; Kelly, B.; Loke, P.; Sajid, M. Proteases in parasitic diseases. Annu. Rev. Pathol. Mech. Dis. 2006, 1, 497–536. [Google Scholar] [CrossRef]
- Valle, A.; Leiro, J.M.; Pereiro, P.; Figueras, A.; Novoa, B.; Dirks, R.P.H.; Lamas, J. Interactions between the parasite Philasterides dicentrarchi and the immune system of the turbot Scophthalmus maximus. A transcriptomic analysis. Biology 2020, 9, 337. [Google Scholar] [CrossRef]
- Folgueira, I.; Lamas, J.; de Felipe, A.P.; Sueiro, R.A.; Leiro, J.M. Identification and Molecular Characterization of Superoxide Dismutases Isolated From A Scuticociliate Parasite: Physiological Role in Oxidative Stress. Sci. Rep. 2019, 9, 13329:1–13329:14. [Google Scholar] [CrossRef]
- Dai, J.R.; Wang, W.; Liang, Y.S.; Li, H.J.; Guan, X.H.; Zhu, Y.C. A novel molluscicidal formulation of niclosamide. Parasitol. Res. 2008, 103, 405–412. [Google Scholar] [CrossRef]
- Cogliano, V.J.; Grosse, Y.; Baan, R.A.; Straif, K.; Secretan, M.B.; Ghissassi, F.E. Meeting Report: Summary of IARC Monographs on Formaldehyde, 2-Butoxyethanol, and 1-tert-Butoxy-2-Propanol. Environ. Health Perspect. 2005, 113, 1205–1208. [Google Scholar] [CrossRef] [PubMed]
- Fullerton, M.; Khatiwada, J.; Johnson, J.U.; Davis, S.; Williams, L.L. Determination of antimicrobial activity of sorrel (Hibiscus sabdariffa) on Esherichia coli O157:H7 isolated from food, veterinary, and clinical samples. J. Med. Food 2011, 14, 950–956. [Google Scholar] [CrossRef]
- Xu, A.; Shang-Guan, J.; Li, Z.; Gao, Z.; Huang, Y.; Chen, Q. Effects of garlic powder on feeding attraction activity, growth and digestive enzyme activities of Japanese seabass, Lateolabrax japonicus. Aquac. Nutr. 2020, 26, 390–399. [Google Scholar] [CrossRef]
- Da-Costa-Rocha, L.; Bonnlaender, B.; Sievers, H.; Pischel, I.; Heinrich, M. Hibiscus sabdariffa L.—A phytochemical and pharmacological review. Food Chem. 2014, 165, 424–443. [Google Scholar] [CrossRef]
- Borrás-Linares, I.; Fernández-Arroyo, S.; Arráez-Roman, D.; Palmeros-Suárez, P.A.; Del Val-Díaz, R.; Andrade-Gonzáles, I.; Fernández-Gutiérrez, A.; Gómez-Leyva, J.F.; Segura-Carretero, A. Characterization of phenolic compounds, anthocyanidin, antioxidant and antimicrobial activity of 25 varieties of Mexican Roselle (Hibiscus sabdariffa). Ind. Crop. Prod. 2015, 69, 385–394. [Google Scholar] [CrossRef]
- Tsai, P.J.; McIntosh, J.; Pearce, P.; Camden, B.; Jordan, B.R. Anthocyanin and antioxidant capacity in Roselle (Hibiscus sabdariffa L.) extract. Food Res. Int. 2002, 35, 351–356. [Google Scholar] [CrossRef]
- Mak, Y.W.; Chuah, L.O.; Ahmad, R.; Bhat, R. Antioxidant and antibacterial activities of hibiscus (Hibiscus rosa-sinensis L.) and Cassia (Senna bicapsularis L.) flower extracts. J. King Saud Univ. Sci. 2013, 25, 275–282. [Google Scholar] [CrossRef]
- Higginbotham, K.L.; Burris, K.P.; Zivanovic, S.; Davidson, P.M.; Stewart, C.N. Antimicrobial activity of Hibiscus sabdariffa aqueous extracts against Escherichia coli O157:H7 and Staphylococcus aureus in a microbiological medium and milk of various fat concentrations. J. Food Prot. 2014, 77, 262–268. [Google Scholar] [CrossRef]
- Mensah, J.K.; Golomeke, D. Antioxidant and antimicrobial activities of the extracts of the calyx of Hibiscus sabdariffa Linn. Curr. Sci. Perspect. 2015, 1, 69–76. [Google Scholar]
- Youns, M.A.D.; Hashim, A.I.; Suliman, I.S.; Alla, A.B.A. In vitro evaluation of the antimicrobial and antioxidant activity of Hibiscus sabdariffa leaves. J. Dent. Med. Sci. 2018, 17, 69–74. [Google Scholar]
- Iglesias, R.; Paramá, A.; Álvarez, M.F.; Leiro, J.; Aja, C.; Sanmartín, M.L. In vitro growth requirements for the fish pathogen Philasterides dicentrarchi (Ciliophora, Scuticociliatida). Vet. Parasitol. 2003, 111, 19–30. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [CrossRef] [PubMed]
- Frasco, M.F.; Guilhermino, L. Effects of dimethoate and beta-naphthoflavone on selected biomarkers of Poecilia reticulata. Fish Physiol. Biochem. 2002, 26, 149–156. [Google Scholar] [CrossRef]
- Schaedle, M.; Bassham, J.A. Chloroplast Glutathione Reductase. Plant Physiol. 1977, 59, 1011–1012. [Google Scholar] [CrossRef] [PubMed]
- Paglia, D.E.; Valentine, W.N. Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J. Lab. Clin. Med. 1967, 70, 158–169. [Google Scholar] [PubMed]
- Cichoski, A.J.; Rotta, R.B.; Scheuermann, G.; Cunha Junior, A.; Smanioto Barin, J. Investigation of glutathione peroxidase activity in chicken meat under different experimental conditions. Food Sci. Technol. 2012, 32, 661–667. [Google Scholar] [CrossRef]
- Tietze, F. Enzymic method for quantitative determination of nanogram amounts of total and oxidized glutathione: Applications to mammalian blood and other tissues. Anal. Biochem. 1969, 27, 502–522. [Google Scholar] [CrossRef]
- Claiborne, A. Handbook of Methods for Oxygen Radical Research, 1st ed.; Greenwald, R.A., Ed.; CRC Press: Boca Raton, FL, USA, 1985; Catalase Activity; pp. 283–284. ISBN 9781351072922. [Google Scholar]
- Ellman, G.L.; Courtney, K.D.; Andres, V.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Cupp-Enyard, C.; Aldrich, S. Use of the Protease Fluorescent Detection Kit to Determine Protease Activity. J. Vis. Exp. 2009, 30, 1514:1–1514:3. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Rodríguez-Medina, I.C.; Beltrán-Debón, R.; Molina, V.M.; Alonso-Villaverde, C.; Joven, J.; Menéndez, J.A.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Direct characterization of aqueous extract of Hibiscus sabdariffa using HPLC with diode array detection coupled to ESI and ion trap MS. J. Sep. Sci. 2009, 32, 3441–3448. [Google Scholar] [CrossRef]
- Ramirez-Rodrigues, M.M.; Plaza, M.; Azeredo, A.; Balaban, M.O.; Marshall, M.R. Physicochemical and phytochemical properties of cold and hot water extraction from Hibiscus sabdariffa. J. Food Sci. 2011, 76, C428–C435. [Google Scholar] [CrossRef]
- Jabeur, I.; Pereira, E.; Barros, L.; Calhelha, R.C.; Soković, M.; Oliveira, M.B.P.P.; Ferreira, I.C.F.R. Hibiscus sabdariffa L. as a source of nutrients, bioactive compounds and colouring agents. Food Res. Int. 2017, 100, 717–723. [Google Scholar] [CrossRef]
- Jabeur, I.; Pereira, E.; Caleja, C.; Calhelha, R.C.; Soković, M.; Catarino, L.; Barros, L.; Ferreira, I.C.F.R. Exploring the chemical and bioactive properties of: Hibiscus sabdariffa L. calyces from Guinea-Bissau (West Africa). Food Funct. 2019, 10, 2234–2243. [Google Scholar] [CrossRef]
- Miyamae, Y.; Kurisu, M.; Han, J.; Isoda, H.; Shigemori, H. Structure-activity relationship of caffeoylquinic acids on the accelerating activity on ATP production. Chem. Pharm. Bull. 2011, 59, 502–507. [Google Scholar] [CrossRef]
- Bajko, E.; Kalinowska, M.; Borowski, P.; Siergiejczyk, L.; Lewandowski, W. 5-O-Caffeoylquinic acid: A spectroscopic study and biological screening for antimicrobial activity. LWT Food Sci. Technol. 2006, 65, 471–479. [Google Scholar] [CrossRef]
- Herranz-López, M.; Fernández-Arroyo, S.; Pérez-Sanchez, A.; Barrajón-Catalán, E.; Beltrán-Debón, R.; Menéndez, J.A.; Alonso-Villaverde, C.; Segura-Carretero, A.; Joven, J.; Micol, V. Synergism of plant-derived polyphenols in adipogenesis: Perspectives and implications. Phytomedicine 2012, 19, 253–261. [Google Scholar] [CrossRef]
- Long, Q.; Chen, H.; Yang, W.; Zhang, L. Delphinidin-3-sambubioside from Hibiscus sabdariffa. L attenuates hyperlipidemia in high fat diet-induced obese rats and oleic acid-induced steatosis in HepG2 cells. Bioengineered 2021, 12, 3837–3849. [Google Scholar] [CrossRef]
- Lazalde-Cruz, R.; Miranda-Romero, L.A.; Tirado-González, D.N.; Carrillo-Díaz, M.I.; Medina-Cuéllar, S.E.; Mendoza-Martínez, G.D.; Lara-Bueno, A.; Tirado-Estrada, G.; Salem, A.Z.M. Potential Effects of Delphinidin-3-O-sambubioside and Cyanidin-3-O-sambubioside of Hibiscus sabdariffa L. in Ruminant Meat and Milk Production and Quality. Animals 2021, 11, 2827. [Google Scholar] [CrossRef]
- Segura-Carretero, A.; Puertas-Mejía, M.A.; Cortacero-Ramírez, S.; Beltrán, R.; Alonso-Vilaverde, C.; Joven, J.; Dinelli, G.; Fernández-Gutiérrez, A. Selective extraction, separation, and identification of anthocyanins from Hibiscus sabdariffa L. using solid phase extraction-capillary electrophoresis-mass spectrometry (time-of-flight/ion trap). Electrophoresis 2008, 29, 2852–2861. [Google Scholar] [CrossRef]
- Beltrán-Debón, R.; Alonso-Villaverde, C.; Aragonès, G.; Rodríguez-Medina, I.; Rull, A.; Micol, V.; Segura-Carretero, A.; Fernández-Gutiérrez, A.; Camps, J.; Joven, J. The aqueous extract of Hibiscus sabdariffa calices modulates the production of monocyte chemoattractant protein-1 in humans. Phytomedicine 2010, 17, 186–191. [Google Scholar] [CrossRef]
- Chakraborty, M.; Mitra, A. The antioxidant and antimicrobial activity properties of the methanolic extract from Cocos nucifera mesocarp. Food Chem. 2008, 107, 994–999. [Google Scholar] [CrossRef]
- García-Marino, M.; Hernández-Hierro, J.M.; Rivas-Gonzalo, J.C.; Escribano-Bailón, M.T. Colour and pigment composition of red wines obtained from co-maceration of Tempranillo and Graciano varieties. Anal. Chim. Acta 2010, 660, 134–142. [Google Scholar] [CrossRef]
- Ramirez-Lopez, L.M.; DeWitt, C.A.M. Analysis of phenolic compounds in commercial dried grape pomace by high-performance liquid chromatography electrospray ionization mass spectrometry’. Food Sci. Nutr. 2014, 2, 470–477. [Google Scholar] [CrossRef] [PubMed]
- Kähkönen, M.P.; Heinonen, M. Antioxidant activity of anthocyanins and their aglycons. J. Agric. Food Chem. 2003, 51, 628–633. [Google Scholar] [CrossRef]
- David, A.V.A.; Arulmoli, R.; Parasuraman, S. Overviews of biological importance of quercetin: A bioactive flavonoid. Pharmacogn. Rev. 2016, 10, 84–89. [Google Scholar] [CrossRef]
- Patel, K.; Patel, D.K. The beneficial role of rutin, a naturally occurring flavonoid in health promotion and disease prevention: A systematic review and update. In Bioactive Food as Dietary Interventions for Arthritis and Related Inflammatory Diseases, 2nd ed.; Academic Press: London, UK; Elsevier: London, UK, 2019; pp. 457–479. ISBN 978-0-12-813820-5. [Google Scholar]
- Taheri, Y.; Suleria, H.A.R.; Martins, N.; Sytar, O.; Beyatli, A.; Yeskaliyeva, B.; Seitimova, G.; Salehi, B.; Semwal, P.; Painuli, S.; et al. Myricetin bioactive effects: Moving from preclinical evidence to potential clinical applications. BMC Complement. Med. Ther. 2020, 20, 241. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Yu, L.; Wang, M.H. N-trans-feruloyltyramine inhibits LPS-induced NO and PGE2 production in RAW 264.7 macrophages: Involvement of AP-1 and MAP kinase signalling pathways. Chem. Biol. Interact. 2015, 235, 56–62. [Google Scholar] [CrossRef]
- Ezzat, S.M.; Salama, M.M.; el-Din, S.H.; Saleh, S.; El-Lakkany, N.M.; Hammam, O.A.; Salem, M.B.; Botros, S.S. Metabolic profile and hepatoprotective activity of the anthocyanin-rich extract of Hibiscus sabdariffa calyces. Pharm. Biol. 2016, 54, 3172–3181. [Google Scholar] [CrossRef]
- Osman, A.G.; Chittiboyina, A.G.; Khan, I.A. Cytoprotective role of dietary phytochemicals against cancer development via induction of phase II and antioxidant enzymes. In Advances in Molecular Toxicology, 1st ed.; Academic Press: Cambridge, MA, USA; Elsevier: Cambridge, MA, USA, 2016; Volume 10, pp. 99–137. ISBN 978-0-12-804700-2. [Google Scholar]
- Reverter, M.; Bontemps, N.; Lecchini, D.; Banaigs, B.; Sasal, P. Use of plant extracts in fish aquaculture as an alternative to chemotherapy: Current status and future perspectives. Aquaculture 2015, 433, 50–61. [Google Scholar] [CrossRef]
- Jack, J.D.; Gilbert, J.J. Effects of Metazoan Predators on Ciliates in Freshwater Plankton Communities. J. Eukaryot. Microbiol. 1997, 44, 194–199. [Google Scholar] [CrossRef]
- Plaskova, A.; Mlcek, J. New insights of the application of water or ethanol-water plant extract rich in active compounds in food. Front. Nutr. 2023, 10, 1118761. [Google Scholar] [CrossRef]
- Borges, A.; José, H.; Homem, V.; Simões, M. Comparison of techniques and solvents on the antimicrobial and antioxidant potential of extracts from Acacia dealbata and Olea europaea. Antibiotics 2020, 9, 48. [Google Scholar] [CrossRef]
- Costantini, D. Oxidative Stress and Hormesis in Evolutionary Ecology and Physiology: A Marriage between Mechanistic and Evolutionary Approaches, 1st ed.; Springer: Berlin/Heidelberg, Germany, 2014; ISBN 978-3-642-54662-4. [Google Scholar]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine, 5th ed.; Oxford University Press: Oxford, UK, 2015; ISBN 978–0–19–871747–8. [Google Scholar]
- Sheehan, D.; Meade, G.; Foley, V.M.; Dowd, C.A. Structure, function and evolution of glutathione transferases: Implications for classification of non-mammalian members of an ancient enzyme superfamily. Biochem. J. 2001, 360 Pt 1, 1–16. [Google Scholar] [CrossRef]
- Margis, R.; Dunand, C.; Teixeira, F.K.; Margis-Pinheiro, M. Glutathione peroxidase family—An evolutionary overview. FEBS J. 2008, 275, 3959–3970. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.S.; Chan, H.W.; Yu, L.C. Glutathione peroxidase and glutathione reductase activities are partially responsible for determining the susceptibility of cells to oxidative stress. Toxicology 2006, 226, 126–130. [Google Scholar] [CrossRef]
- Bachhawat, A.K.; Yadav, S. The glutathione cycle: Glutathione metabolism beyond the γ-glutamyl cycle. IUBMB Life 2018, 70, 585–592. [Google Scholar] [CrossRef]
- Banyal, H.S.; Sharma, S.K. Glutathione metabolism in parasitic protozoa. J. Parasit. Dis. 2007, 31, 92–102. [Google Scholar]
- Müller, S. Role and regulation of glutathione metabolism in Plasmodium falciparum. Molecules 2015, 20, 10511–10534. [Google Scholar] [CrossRef]
- Kim, S.H.; Kim, S.J.; Lee, J.S.; Lee, Y.M. Acute effects of heavy metals on the expression of glutathione-related antioxidant genes in the marine ciliate Euplotes crassus. Mar. Pollut. Bull. 2014, 85, 455–462. [Google Scholar] [CrossRef]
- Zou, S.; Zhang, Q.; Gong, J. Comparative transcriptomics reveals distinct gene expressions of a model ciliated protozoan feeding on bacteria-free medium, digestible, and digestion-resistant bacteria. Microorganisms 2020, 8, 559. [Google Scholar] [CrossRef] [PubMed]
- Livingstone, D.R. Contaminant-stimulated reactive oxygen species production and oxidative damage in aquatic organisms. Mar. Poll. Bull. 2001, 42, 656–666. [Google Scholar] [CrossRef] [PubMed]
- Lamas, J.; Morais, P.; Arranz, J.A.; Sanmartín, M.L.; Orallo, F.; Leiro, J. Resveratrol promotes an inhibitory effect on the turbot scuticociliate parasite Philasterides dicentrarchi by mechanisms related to cellular detoxification. Vet. Parasitol. 2009, 161, 307–315. [Google Scholar] [CrossRef]
- Amaroli, A. Neurotransmitters or Biomediators? The Cholinergic System in Protozoa. EC Microbiol. 2017, 7, 40–41. [Google Scholar]
- Bullock, T.H.; Nachmansohn, D. Choline esterase in primitive nervous systems. J. Cell. Comp. Physiol. 1942, 20, 239–242. [Google Scholar] [CrossRef]
- Trielli, F.; Amaroli, A.; Sifredi, F.; Marchi, B.; Falugi, C.; Corrado, M.U.D. Effects of xenobiotic compounds on the cell activities of Euplotes crassus, a single-cell eukaryotic test organism for the study of the pollution of marine sediments. Aquat. Toxicol. 2007, 83, 272–283. [Google Scholar] [CrossRef]
- Corrado, M.U.D.; Politi, H.; Trielli, F.; Angelini, C.; Falugi, C. Evidence for the presence of a mammalian-like cholinesterase in Paramecium primaurelia (Protista, ciliophora) developmental cycle. J. Exp. Zool. 1999, 283, 102–105. [Google Scholar] [CrossRef]
- Wolkmer, P.; da Silva, C.B.; Paim, F.C.; Da Silva, A.S.; Tavares, K.C.S.; Lazzarotto, C.R.; Palma, H.E.; Thomé, G.R.; Miletti, L.C.; Schetinger, M.R.C.; et al. Biochemistry detection of acetylcholinesterase activity in Trypanosoma evansi and possible functional correlations. Exp. Parasitol. 2012, 132, 546–549. [Google Scholar] [CrossRef]
- Acorn, A.R.; Fraser Clark, K.; Jones, S.; Després, B.M.; Munro, S.; Cawthorn, R.J.; Greenwood, S.J. Analysis of expressed sequence tags (ESTs) and gene expression changes under different growth conditions for the ciliate Anophryoides haemophila, the causative agent of bumper car disease in the American lobster (Homarus americanus). J. Inverteb. Pathol. 2011, 107, 146–154. [Google Scholar] [CrossRef]
- Murase, L.S.; Souza, J.V.P.; Neto, Q.A.L.; Mello, T.F.P.M.; Cardoso, B.M.; Lera-Nonose, D.S.S.L.; Teixeira, J.J.V.T.; Lonardoni, M.V.C.; Demarchi, I.G. The role of metalloproteases in Leishmania species infection in the New World: A systematic review. Parasitology 2018, 145, 1499–1509. [Google Scholar] [CrossRef] [PubMed]
- Thiakaki, M.; Kolli, B.; Chang, K.P.; Soteriadou, K. Down-regulation of gp63 level in Leishmania amazonensis promastigotes reduces their infectivity in BALB/c mice. Microbes Infect. 2006, 8, 1455–1463. [Google Scholar] [CrossRef] [PubMed]
- Srikanth, S.; Chen, Z. Plant protease inhibitors in therapeutics-focus on cancer therapy. Front. Pharmacol. 2016, 7, 470. [Google Scholar] [CrossRef] [PubMed]
- Abu-Tarboush, H.M.; Ahmed, S.A.B. Studies on karkade (Hibiscus sabdariffa): Protease inhibitors, phytate, in vitro protein digestibility and gossypol content. Food Chem. 1996, 56, 15–19. [Google Scholar] [CrossRef]
- Loo, S.; Kam, A.; Xiao, T.; Nguyen, G.K.T.; Liu, F.; Tam, J.P. Identification and characterization of roseltide, a knottin-type neutrophil elastase inhibitor derived from Hibiscus sabdariffa. Sci. Rep. 2016, 6, 39401. [Google Scholar] [CrossRef]
- Kam, A.; Loo, S.; Fan, J.S.; Sze, S.K.; Yang, D.; Tam, J.P. Roseltide rT7 is a disulfide-rich, anionic, and cell-penetrating peptide that inhibits proteasomal degradation. J. Biol. Chem. 2019, 294, 19604–19615. [Google Scholar] [CrossRef]
- Ali, B.H.; Al Wabel, N.; Blunden, G. Phytochemical, pharmacological and toxicological aspects of Hibiscus sabdariffa L.: A review. Phytother. Res. 2005, 19, 369–375. [Google Scholar] [CrossRef]
- McKerrow, J.H.; Rosenthal, P.J.; Swenerton, R.; Doyle, P. Development of protease inhibitors for protozoan infections. Curr. Opin. Infect. Dis. 2008, 21, 668–672. [Google Scholar] [CrossRef]
- Zheoat, A.M.; Gray, A.I.; Igoli, J.O.; Ferro, V.A.; Drummond, R.M. Hibiscus acid from Hibiscus sabdariffa (Malvaceae) has a vasorelaxant effect on the rat aorta. Fitoterapia 2019, 134, 5–13. [Google Scholar] [CrossRef]
- Alonso, J.; Zamilpa, A.; Aguilar, F.A.; Herrera-Ruiz, M.; Tortoriello, J.; Jimenez-Ferrer, E. Pharmacological characterization of the diuretic effect of Hibiscus sabdariffa Linn (Malvaceae) extract. J. Ethnopharmacol. 2012, 139, 751–756. [Google Scholar] [CrossRef]
- Ganzon, J.G.; Chen, L.G.; Wang, C.C. 4-O-Caffeoylquinic acid as an antioxidant marker for mulberry leaves rich in phenolic compounds. J. Food Drug Anal. 2018, 26, 985–993. [Google Scholar] [CrossRef]
- Li, Y.; Yao, J.; Han, C.; Yang, J.; Chaudhry, M.T.; Wang, S.; Liu, H.; Yin, Y. Quercetin, inflammation and immunity. Nutrients 2016, 8, 167. [Google Scholar] [CrossRef] [PubMed]
- Hou, D.X.; Tong, X.; Terahara, N.; Luo, D.; Fujii, M. Delphinidin 3-sambubioside, a Hibiscus anthocyanin, induces apoptosis in human leukemia cells through reactive oxygen species-mediated mitochondrial pathway. Arch. Biochem. Biophys. 2005, 440, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Abeda, H.Z.; Kouassi, M.K.; Yapo, K.D.; Koffi, E.; Sie, R.S.; Kone, M.; Kouakou, H.T. Production and enhancement of anthocyanin in callus line of roselle (Hibiscus sabdariffa L.). Int. J. Recent Biotechnol. 2014, 2, 45–56. [Google Scholar]
- Opletal, L.; Chocholousova-Havlikova, L.; Siatka, T.; Cahliková, L.; Locarek, M.; Ali, B.H.; Manoj, P.; Ramkumar, A.; Al Suleimani, Y.M.; Al Za’abi, M.; et al. Preparation and validated analysis of anthocyanin concentrate from the calyces of Hibiscus sabdariffa. Nat. Prod. Commun. 2017, 12, 43–45. [Google Scholar] [CrossRef]
- Gandhi, S.P.; Lokhande, K.B.; Swamy, V.K.; Nanda, R.K.; Chitlange, S.S. Computational data of phytoconstituents from Hibiscus rosa-sinensis on various anti-obesity targets. Data Brief 2019, 24, 103994. [Google Scholar] [CrossRef]
- Olivas-Aguirre, F.J.; Rodrigo-García, J.; Martínez-Ruiz, N.D.R.; Cárdenas-Robles, A.I.; Mendoza-Díaz, S.O.; Álvarez-Parrilla, E.; González-Aguilar, G.A.; De La Rosa, L.A.; Ramos-Jiménez, A.; Wall-Medrano, A. Cyanidin-3-O-glucoside: Physical-chemistry, foodomics and health effects. Molecules 2016, 21, 1264. [Google Scholar] [CrossRef]
- Noda, Y.; Kaneyuki, T.; Mori, A.; Packer, L. Antioxidant activities of pomegranate fruit extract and its anthocyanidins: Delphinidin, cyanidin, and pelargonidin. J. Agric. Food Chem. 2002, 50, 166–171. [Google Scholar] [CrossRef] [PubMed]
- Beninger, C.W.; Hosfield, G.L. Antioxidant activity of extracts, condensed tannin fractions, and pure flavonoids from Phaseolus vulgaris L. seed coat color genotypes. J. Agric. Food Chem. 2003, 51, 7879–7883. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Proteases | Origin, Accession Number | Primer Sets (5′ 3′) | Amplicon Size (bp) | Efficiency of Primers (%) |
|---|---|---|---|---|---|
| PD1 | Cathepsin L-like cysteine protease | Philasterides dicentrarchi, AFF59216.1 | F: TCTTGAGAGCTTCTGCTGCCAC | 271 | 105.1 |
| R: TCTTGGATGTTTAATTCGGTGCTGT | |||||
| PD2 | Catepsin 90 protease | Philasterides dicentrarchi, QBH22552.1 | F: TAGCTTCAATTGCTTCTGGTAGTCTTG | 227 | 105.3 |
| R: ATCCATGTTTATTCCACATAGTCCATTAC | |||||
| PD3 | Leishmanolysin family protein | Philasterides dicentrarchi, QBH22559.1 | F: TGTTTTAGAAGATTCTGGATTTTATG | 331 | 105.4 |
| R: TATGTCAATTATATTACTGTAGAAGC | |||||
| ß-tubulin | - | Philasterides dicentrarchi, MH444695.1 | F: GTATGATCATTGATAACGAAGCCCTCTACG | 323 | 104.3 |
| Parameter | One-Way ANOVA Outcome | Toxicity Point Estimates | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| DF | SS | MS | F | p | NOEC | LOEC | IC50 | CL | R2 | |
| PGR | 7 | 11,841.83 | 1691.69 | 30.413 | <0.001 | 0.15 | 0.29 | 1.57 | 1.09–2.31 | 0.67 |
| GST | 7 | 444.035 | 63.434 | 2.429 | 0.029 | 0.52 | 0.81 | nd | nd | nd |
| GR | 7 | 15,743.5 | 2249.071 | 67.499 | <0.001 | 0.14 | 0.22 | nd | nd | nd |
| GPx | 7 | 1184.466 | 169.209 | 7.289 | <0.001 | 0.22 | 0.34 | nd | nd | nd |
| TG | 7 | 1920.084 | 274.298 | 29.331 | <0.001 | 0.14 | 0.22 | nd | nd | nd |
| CAT | 7 | 368.225 | 52.604 | 4.403 | <0.001 | 0.81 | 1.25 | nd | nd | nd |
| AChE | 7 | 8.675 | 1.239 | 5.07 | <0.001 | 0.81 | 1.25 | nd | nd | nd |
| PA | 6 | 0.0548 | 0.00913 | 37.99 | <0.001 | 0.22 | 0.34 | 0.76 | 0.45–1.45 | 0.76 |
| Peak # | RT (min) | λmax | [M-H]− (m/z) | MS2 Fragments (m/z) | Proposed Compound | Chemical Class | References | Bioactivity | References |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 4.44 | 277 | 189 | [189] 127 | hibiscus acid | Organic acids | [33,34] | Anti-diabetic | [14] |
| 2 | 5.27 | 324 | 353 | [353] 191; 179 | 3-O-caffeoylquinic acid | Phenolic acid (polyphenol) | [35,36] | Antioxidant, antibacterial, anticancer, antihistamine | [37,38] |
| 3 | 7.46 | 281; 523 | 595 | [595] 355; 300 | delphinidin-3-sambubioside | Anthocyanin (polyphenol) | [39] | Antioxidant, antibacterial, antiviral, anthelmintic | [14,40,41] |
| 4 | 8.04 | 325 | 353 | [353] 191; 173 | 4-O-caffeoylquinic acid | Phenolic acid (polyphenol) | [35] | Antioxidant, antibacterial, anticancer, antihistamine | [37,38] |
| 5 | 8.54 | 293; 325 | 353; 398.7 | nd | Unknown | nd | nd | nd | nd |
| 6 | 9.91 | 276; 528 | 263 | [263] 217 | hibiscus acid hydroxyethyldimethylesther | Organic acids | [15] | nd | nd |
| 7 | 10.22 | 317 | 611 | [611] | cyanidin 3,5-O-diglucoside | Anthocyanin (polyphenol) | [42] | Antioxidant, antibacterial, antiviral, anthelmintic | [40,41] |
| 353 | [353] 191 | 5-O-caffeoylquinic acid (5-CQA) | Phenolic acid (polyphenol) | [33,35,43] | Antioxidant, antibacterial, anticancer, antihistamine | [37,38] | |||
| 8 | 10.89 | 328 | 335 | [335] 161; 135 | 5-O-caffeoylshikimic acid | Phenolic acid (polyphenol) | [15,33,43] | Potentially antioxidant and antimicrobial | [44] |
| 479 | [479] 317 | petunidin-3-O-glucoside | Anthocyanin (polyphenol) | [45,46] | Antioxidant | [47] | |||
| 9 | 11.16 | 354 | 595 | [595] 299.9 | quercetin-O-sambubioside | Flavonoid (polyphenol) | [33] | Antibacterial, antiviral, anti-inflammatory | [48] |
| 10 | 11.31 | 240; 325 | 381 | [381] 161 | 5-CQA:formic acid (FA) complex | Phenolic acid (polyphenol) + organic acid | Based in 5-CQA m/z [33] | nd | nd |
| (335 (5-CQA) + 46 (FA) = 381) | |||||||||
| 11 | 11.66 | 256; 352 | 609 | [609] 301 | quercetin-3-O-rutinoside | Flavonoid (polyphenol) | [33] | Antibacterial, antitumoral, anti-inflammatory, anti-allergenic, antiviral, vasoactive, antiprotozoal | [49] |
| 12 | 12.06 | 256; 353 | 463 | [463] 301 | quercetin-3-O-diglucoside | Flavonoid (polyphenol) | [33] | Antibacterial, antiviral, anti-inflammatory | [48] |
| 13 | 12.33 | 239; 326 | 381 | [381] 161; 135 | Unknown | nd | nd | nd | nd |
| 14 | 12.81 | 242; 327 | 381 | [381] 179; 191; 135 | Unkown | ||||
| 15 | 13.81 | 252; 368 | 317 | [317] 165 | myricetin | Flavonoid (polyphenol) | [39] | Antioxidant, antimicrobial | [50] |
| 16 | 15.13 | 240; 291; 317 | 312 | [312] 297; 178; 135 | N-feruloyltyramide | Flavonoid (polyphenol) | [33,43] | Antioxidant, antimicrobial, anticancer | [51] |
| 17 | 15.97 | 255; 368 | 301 | [301] 179; 151 | quercetin or delphinidin | Flavonoid or Anthocyanin (polyphenol) | [33,43,46,52] | Antioxidant, anti-inflammatory, antibacterial, antiviral | [48,53] |
| 18 | 17.36 | 287 | 597 | [597] 579 | delphinidin-3-O-sambubioside | Anthocyanin (polyphenol) | [34,36,42] | Antioxidant, antibacterial, antiviral, anthelmintic | [14,40,41] |
| Extract | Group of Organisms | Pathogen Species | Parameter | Exposure Time (h) | LOEC or MIC (mg mL−1) | Reference |
|---|---|---|---|---|---|---|
| (In Vitro Assays) | ||||||
| 5-O-Caffeoylquinic acid | Bacteria | Staphylococcus aureus | Growth inihibition | 48 | 10 * | [44] |
| Enterococcus faecium | 10 * | |||||
| Escherichia coli | 10 * | |||||
| Proteus vulgaris | 10 * | |||||
| Pseudomonas aeruginosa | 10 * | |||||
| Klebsiella pneumoniae | 5 * | |||||
| Fungi | Candida albicans | 10 * | ||||
| H. sabdariffa hydroethanolic extract | Bacteria | Staphylococcus aureus | Growth inhibition | 24 | 0.15–0.45 | [35,36] |
| Bacillus cereus | 0.15 | |||||
| Micrococcus flavus | 0.20–0.45 | |||||
| Listeria monocytogenes | 0.15–0.45 | |||||
| Escherichia coli | 0.20 | |||||
| Enterobacter cloacae | 0.15–0.3 | |||||
| Salmonella typhimirium | 0.15–0.45 | |||||
| Pseudomonas aeruginosa | 0.15–0.45 | |||||
| Fungi | Aspergillus fumigatus | Growth inhibition | 72 | 0.30–0.45 | ||
| Aspergillus versicolor | 0.10 | |||||
| Aspergillus ochraceus | 0.15 | |||||
| Aspergillus niger | 0.30–0.60 | |||||
| Penicillium ochrochloron | 0.20–0.30 | |||||
| Penicillium verrucosum var. cyclopium | 0.20–0.30 | |||||
| Trichoderma viride | 0.075 | |||||
| H. sabdariffa infusion | Bacteria | Staphylococcus aureus | Growth inhibition | 24 | 0.20–0.30 | |
| Bacillus cereus | 0.10 | |||||
| Micrococcus flavus | 0.30 | |||||
| Listeria monocytogenes | 0.20–0.30 | |||||
| Escherichia coli | 0.20 | |||||
| Enterobacter cloacae | 0.30–0.15 | |||||
| Salmonella typhimirium | 0.20–0.45 | |||||
| Pseudomonas aeruginosa | 0.20–0.30 | |||||
| Fungi | Aspergillus fumigatus | Growth inhibition | 72 | 0.30–0.60 | ||
| Aspergillus versicolor | 0.15–0.30 | |||||
| Aspergillus ochraceus | 0.04 | |||||
| Aspergillus niger | 0.30–0.60 | |||||
| Penicillium ochrochloron | 0.20–0.30 | |||||
| Penicillium verrucosum var. cyclopium | 0.30–0.60 | |||||
| Trichoderma viride | 0.075–0.30 | |||||
| H. sabdariffa methanolic extract | Bacteria | Escherichia coli | Growth inhibition | 24 | 12.5 | [20] |
| Pseudomonas aeruginosa | 12.5 | |||||
| Klebsiella pneumoniae | 12.5 | |||||
| Salmonella typhi | 12.5 | |||||
| Bacillus subtilis | 6.25 | |||||
| Staphylococcus aureus | 6.25 | |||||
| Protozoa (ciliate) | Giardia lamblia | Mortality | 24, 48, 72, 96 | 0.125 | ||
| H. sabdariffa ethanolic extract | Protozoa (ciliate) | Philasterides dicentrarchi | PGR | 96 | 0.29 | This study |
| Proteases activity | 4 | 0.34 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carvalho, A.; Domingues, I.; Carvalho, C.; Silva, A.M.S.; Soares, A.M.V.M.; Marques, C.R. In Vitro Antiprotozoal Activity of Hibiscus sabdariffa Extract against a Ciliate Causing High Mortalities in Turbot Aquaculture. Biology 2023, 12, 912. https://doi.org/10.3390/biology12070912
Carvalho A, Domingues I, Carvalho C, Silva AMS, Soares AMVM, Marques CR. In Vitro Antiprotozoal Activity of Hibiscus sabdariffa Extract against a Ciliate Causing High Mortalities in Turbot Aquaculture. Biology. 2023; 12(7):912. https://doi.org/10.3390/biology12070912
Chicago/Turabian StyleCarvalho, Ana, Inês Domingues, Carla Carvalho, Artur M. S. Silva, Amadeu M. V. M. Soares, and Catarina R. Marques. 2023. "In Vitro Antiprotozoal Activity of Hibiscus sabdariffa Extract against a Ciliate Causing High Mortalities in Turbot Aquaculture" Biology 12, no. 7: 912. https://doi.org/10.3390/biology12070912
APA StyleCarvalho, A., Domingues, I., Carvalho, C., Silva, A. M. S., Soares, A. M. V. M., & Marques, C. R. (2023). In Vitro Antiprotozoal Activity of Hibiscus sabdariffa Extract against a Ciliate Causing High Mortalities in Turbot Aquaculture. Biology, 12(7), 912. https://doi.org/10.3390/biology12070912