
Unveiling Subtle Geographical Clines: Phenotypic Effects and Dynamics of Circadian Clock Gene Polymorphisms

Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Fly Maintenance
2.2. Identification of Clinal SNPs
2.3. Functional Analysis of Clinal SNPs
2.3.1. Experimental Populations
2.3.2. Locomotor Activity Analysis
2.3.3. Diapause Response
2.3.4. Chill Coma Recovery
3. Results
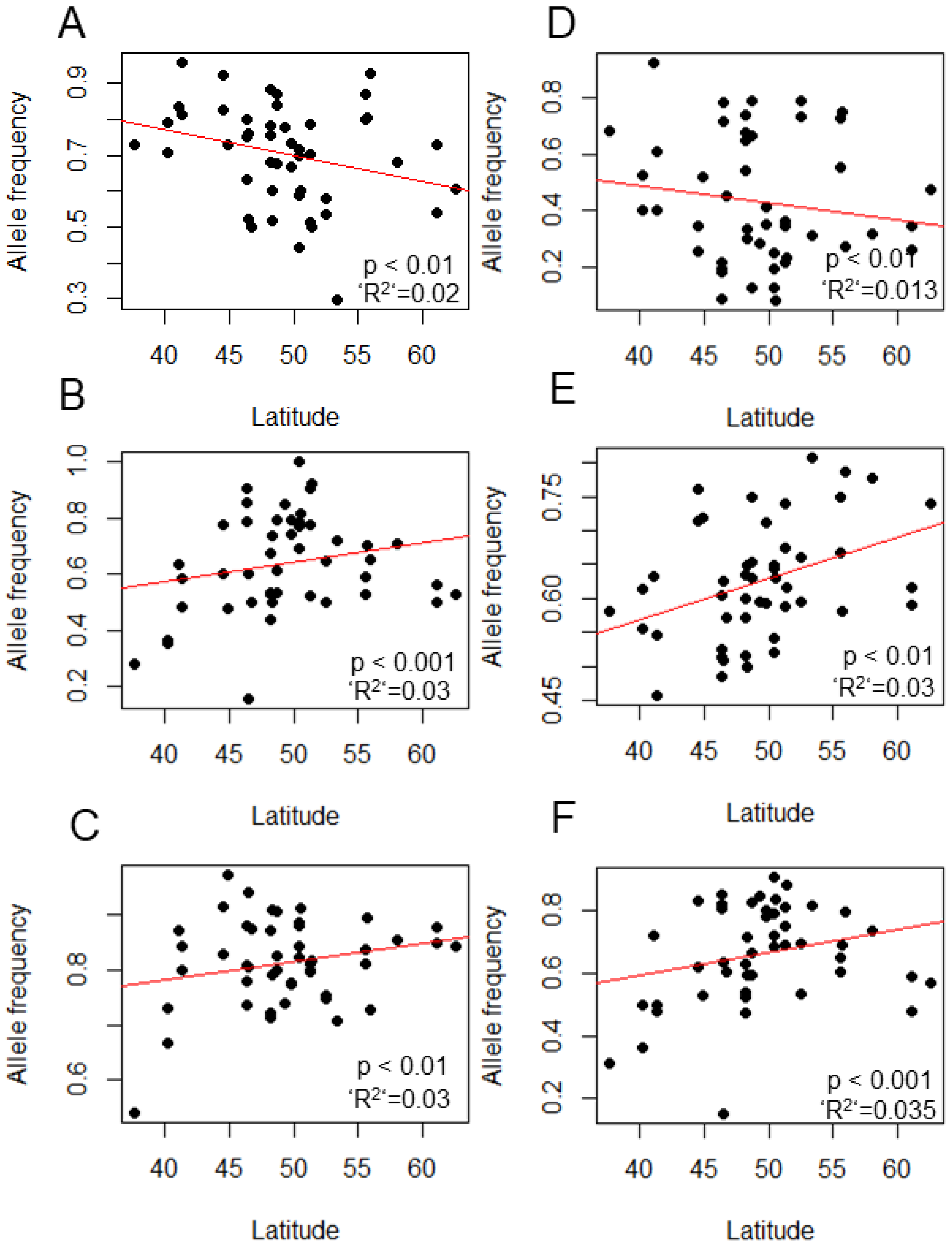
3.1. Identifying Clinal SNPs
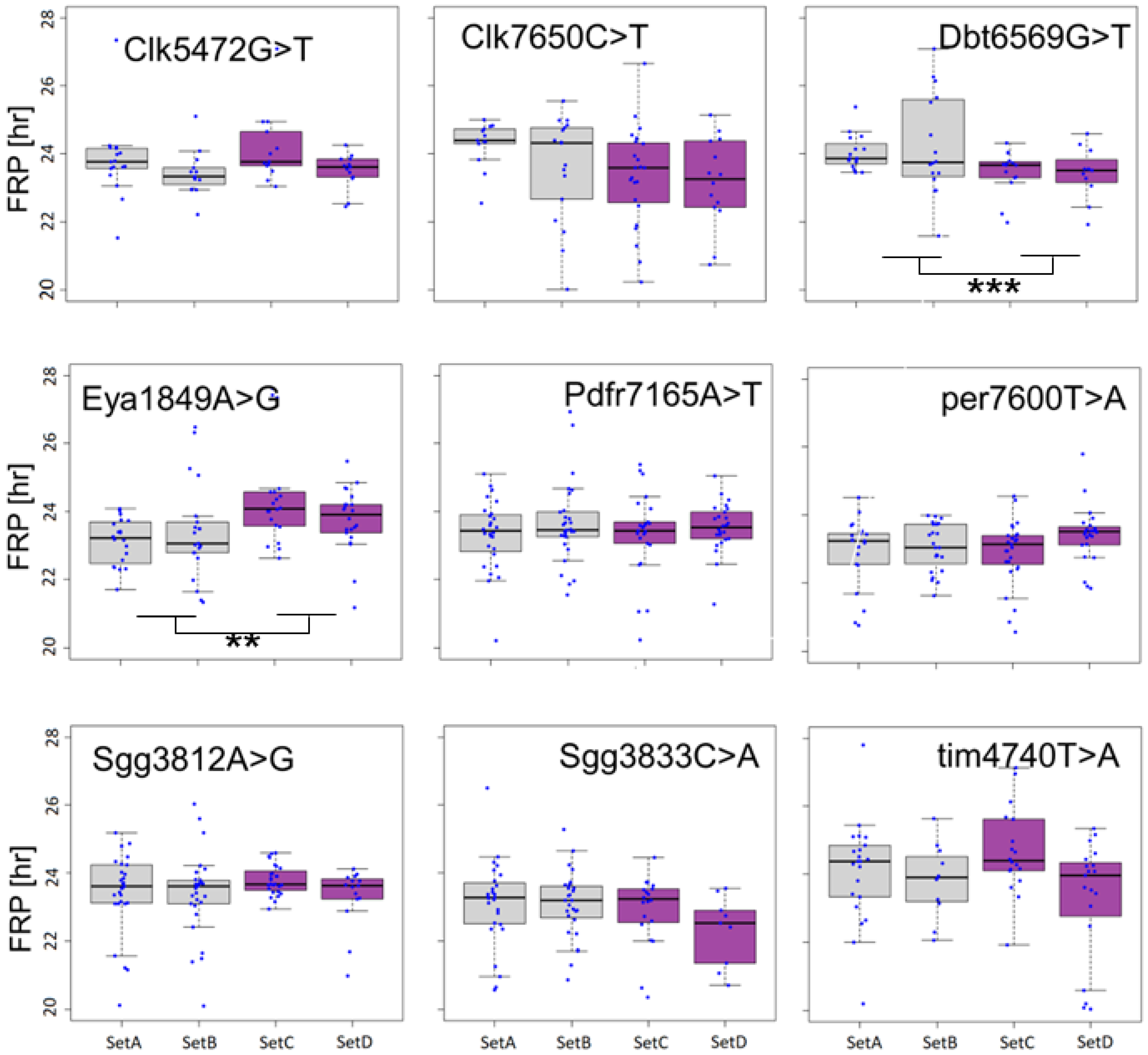
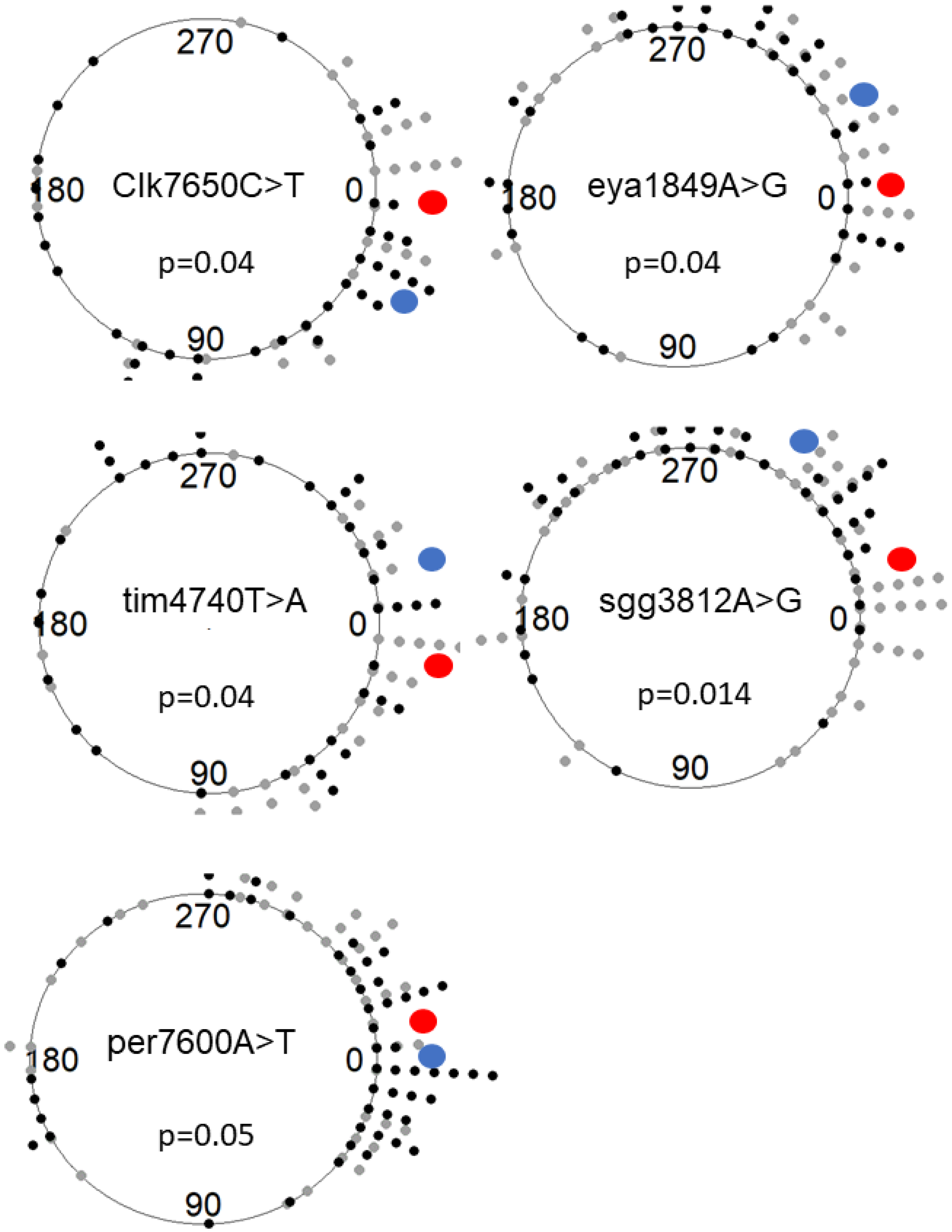
3.2. Association between SNPs and Circadian Phenotypes
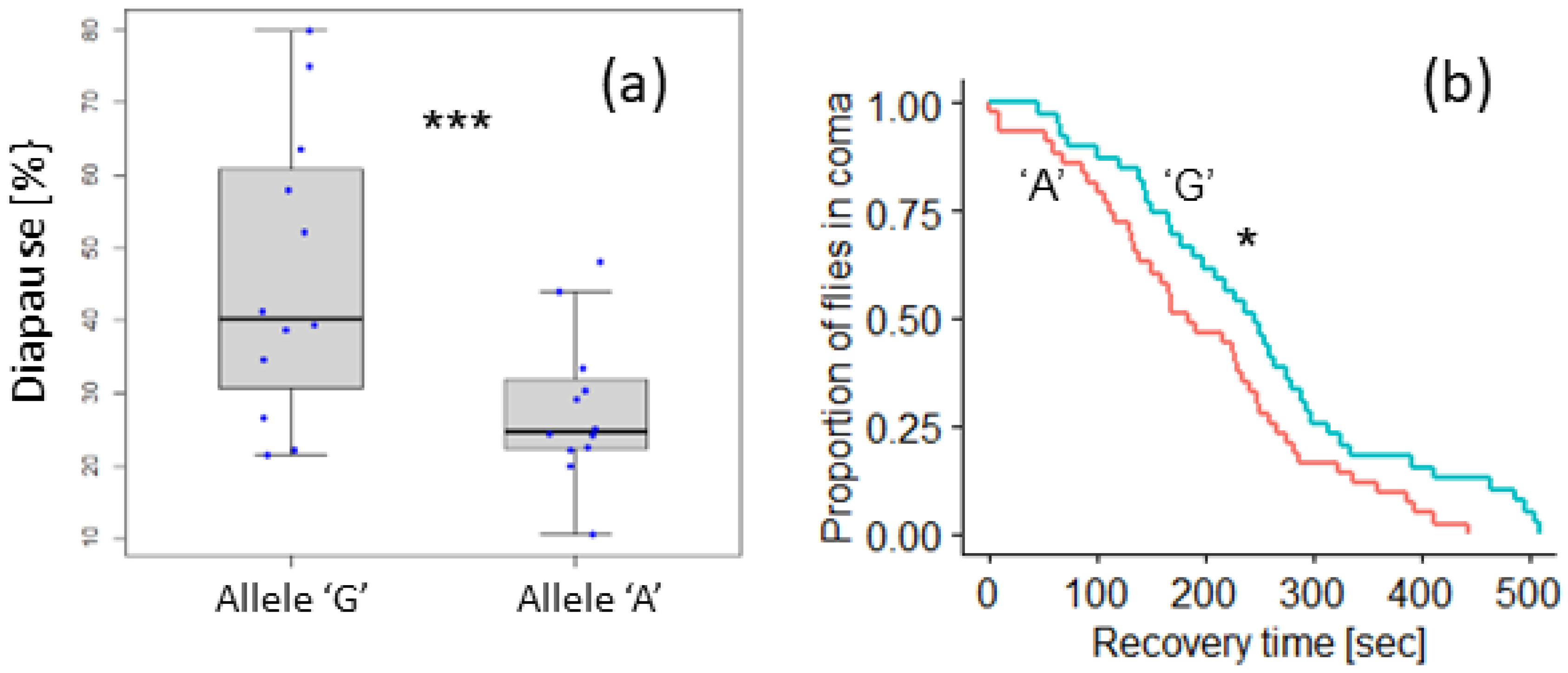
3.3. Polymorphism in Eya Is Associated with Seasonal Phenotypes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rosbash, M. Circadian Rhythms and the Transcriptional Feedback Loop (Nobel Lecture). Angew. Chem. Int. Ed. 2021, 60, 8650–8666. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, J.S. Transcriptional Architecture of the Mammalian Circadian Clock. Nat. Rev. Genet. 2016, 18, 164–179. [Google Scholar] [CrossRef] [PubMed]
- Fetzer, I.; Johst, K.; Schawea, R.; Banitz, T.; Harms, H.; Chatzinotas, A. The Extent of Functional Redundancy Changes as Species’ Roles Shift in Different Environments. Proc. Natl. Acad. Sci. USA 2015, 112, 14888–14893. [Google Scholar] [CrossRef] [PubMed]
- Cyran, S.A.; Buchsbaum, A.M.; Reddy, K.L.; Lin, M.C.; Glossop, N.R.J.; Hardin, P.E.; Young, M.W.; Storti, R.V.; Blau, J. Vrille, Pdp1, and dClock Form a Second Feedback Loop in the Drosophila Circadian Clock. Cell 2003, 112, 329–341. [Google Scholar] [CrossRef] [PubMed]
- Wright, S.; Dobzhansky, T.; Lewontin, R.C.; Moore, J.A.; Provine, W.B.; Wallace, B. Dobzhansky’s Genetics of Natural Populations, I-XLIII. Evolution 1982, 36, 1102. [Google Scholar] [CrossRef]
- Adrion, J.R.; Hahn, M.W.; Cooper, B.S. Revisiting Classic Clines in Drosophila melanogaster in the Age of Genomics. Trends Genet. 2015, 31, 434–444. [Google Scholar] [CrossRef]
- Hut, R.A.; Paolucci, S.; Dor, R.; Kyriacou, C.P.; Daan, S. Latitudinal Clines: An Evolutionary View on Biological Rhythms. Proc. R. Soc. B Biol. Sci. 2013, 280, 20130433. [Google Scholar] [CrossRef]
- Costa, R.; Peixoto, A.A.; Barbujani, G.; Kyriacou, C.P. A Latitudinal Cline in a Drosophila Clock Gene. Proc. R. Soc. B Biol. Sci. 1992, 250, 43–49. [Google Scholar] [CrossRef]
- Tauber, E.; Zordan, M.; Sandrelli, F.; Pegoraro, M.; Osterwalder, N.; Breda, C.; Daga, A.; Selmin, A.; Monger, K.; Benna, C.; et al. Natural Selection Favors a Newly Derived timeless Allele in Drosophila melanogaster. Science 2007, 316, 1895–1898. [Google Scholar] [CrossRef]
- Deppisch, P.; Prutscher, J.M.; Pegoraro, M.; Tauber, E.; Wegener, C.; Helfrich-Förster, C. Adaptation of Drosophila melanogaster to Long Photoperiods of High-Latitude Summers Is Facilitated by the ls-Timeless Allele. J. Biol. Rhythm. 2022, 37, 185–201. [Google Scholar] [CrossRef]
- Lamaze, A.; Chen, C.; Leleux, S.; Xu, M.; George, R.; Stanewsky, R. A Natural timeless Polymorphism Allowing Circadian Clock Synchronization in “White Nights”. Nat. Commun. 2022, 13, 1724. [Google Scholar] [CrossRef] [PubMed]
- Pegoraro, M.; Sayegh Rezek, E.; Fishman, B.; Tauber, E. Nucleotide Variation in Drosophila cryptochrome Is Linked to Circadian Clock Function: An Association Analysis. Front. Physiol. 2022, 13, 96. [Google Scholar] [CrossRef] [PubMed]
- Schlötterer, C.; Tobler, R.; Kofler, R.; Nolte, V. Sequencing Pools of Individuals-Mining Genome-Wide Polymorphism Data without Big Funding. Nat. Rev. Genet. 2014, 15, 749–763. [Google Scholar] [CrossRef]
- Kapun, M.; Barrón, M.G.; Staubach, F.; Obbard, D.J.; Wiberg, R.A.W.; Vieira, J.; Goubert, C.; Rota-Stabelli, O.; Kankare, M.; Bogaerts-Márquez, M.; et al. Genomic Analysis of European Drosophila melanogaster Populations Reveals Longitudinal Structure, Continent-Wide Selection, and Previously Unknown DNA Viruses. Mol. Biol. Evol. 2020, 37, 2661–2678. [Google Scholar] [CrossRef]
- Abrieux, A.; Xue, Y.; Cai, Y.; Lewald, K.M.; Nguyen, H.N.; Zhang, Y.; Chiu, J.C. EYES ABSENT and TIMELESS Integrate Photoperiodic and Temperature Cues to Regulate Seasonal Physiology in Drosophila. Proc. Natl. Acad. Sci. USA 2020, 117, 15293–15304. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Lafuente, E.; Duneau, D.; Beldade, P. Genetic Basis of Thermal Plasticity Variation in Drosophila melanogaster Body Size. PLoS Genet. 2018, 14, e1007686. [Google Scholar] [CrossRef]
- Mackay, T.F.C.; Richards, S.; Stone, E.A.; Barbadilla, A.; Ayroles, J.F.; Zhu, D.; Casillas, S.; Han, Y.; Magwire, M.M.; Cridland, J.M.; et al. The Drosophila melanogaster Genetic Reference Panel. Nature 2012, 482, 173–178. [Google Scholar] [CrossRef]
- Huang, W.; Massouras, A.; Inoue, Y.; Peiffer, J.; Ràmia, M.; Tarone, A.M.; Turlapati, L.; Zichner, T.; Zhu, D.; Lyman, R.F.; et al. Natural Variation in Genome Architecture among 205 Drosophila melanogaster Genetic Reference Panel Lines. Genome Res. 2014, 24, 1193–1208. [Google Scholar] [CrossRef]
- Rosato, E.; Kyriacou, C.P. Analysis of Locomotor Activity Rhythms in Drosophila. Nat. Protoc. 2006, 1, 559–568. [Google Scholar] [CrossRef]
- Chiu, J.C.; Low, K.H.; Pike, D.H.; Yildirim, E.; Edery, I. Assaying Locomotor Activity to Study Circadian Rhythms and Sleep Parameters in Drosophila. J. Vis. Exp. 2010, 43, e2157. [Google Scholar] [CrossRef]
- Zielinski, T.; Moore, A.M.; Troup, E.; Halliday, K.J.; Millar, A.J. Strengths and Limitations of Period Estimation Methods for Circadian Data. PLoS ONE 2014, 9, e96462. [Google Scholar] [CrossRef] [PubMed]
- Pegoraro, M.; Zonato, V.; Tyler, E.R.; Fedele, G.; Kyriacou, C.P.; Tauber, E. Geographical Analysis of Diapause Inducibility in European Drosophila melanogaster Populations. J. Insect Physiol. 2017, 98, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Pegoraro, M.; Gesto, J.S.; Kyriacou, C.P.; Tauber, E. Role for Circadian Clock Genes in Seasonal Timing: Testing the Bünning Hypothesis. PLoS Genet. 2014, 10, e1004603. [Google Scholar] [CrossRef]
- Dardente, H.; Wyse, C.A.; Birnie, M.J.; Dupré, S.M.; Loudon, A.S.I.; Lincoln, G.A.; Hazlerigg, D.G. A Molecular Switch for Photoperiod Responsiveness in Mammals. Curr. Biol. 2010, 20, 2193–2198. [Google Scholar] [CrossRef]
- Masumoto, K.H.; Ukai-Tadenuma, M.; Kasukawa, T.; Nagano, M.; Uno, K.D.; Tsujino, K.; Horikawa, K.; Shigeyoshi, Y.; Ueda, H.R. Acute Induction of Eya3 by Late-Night Light Stimulation Triggers TSHβ Expression in Photoperiodism. Curr. Biol. 2010, 20, 2199–2206. [Google Scholar] [CrossRef]
- Pittendrigh, C.S.; Takamura, T. Latitudinal Clines in the Properties of a Circadian Pacemaker. J. Biol. Rhythm. 1989, 4, 105–123. [Google Scholar] [CrossRef]
- Michael, T.P.; Salomé, P.A.; Yu, H.J.; Spencer, T.R.; Sharp, E.L.; McPeek, M.A.; Alonso, J.M.; Ecker, J.R.; McClung, C.R. Enhanced Fitness Conferred by Naturally Occurring Variation in the Circadian Clock. Science 2003, 302, 1049–1053. [Google Scholar] [CrossRef]
- Vesala, L.; Hoikkala, A. Effects of Photoperiodically Induced Reproductive Diapause and Cold Hardening on the Cold Tolerance of Drosophila montana. J. Insect Physiol. 2011, 57, 46–51. [Google Scholar] [CrossRef]
- Rand, D.M.; Weinreich, D.M.; Lerman, D.; Folk, D.; Gilchrist, G.W. Three Selections Are Better than One: Clinal Variation of Thermal QTL from Independent Selection Experiments in Drosophila. Evolution 2010, 64, 2921. [Google Scholar] [CrossRef]
- Kiat, Y.; Davaasuren, B.; Erdenechimeg, T.; Troupin, D.; Sapir, N. Large-Scale Longitudinal Climate Gradient across the Palearctic Region Affects Passerine Feather Moult Extent. Ecography 2021, 44, 124–133. [Google Scholar] [CrossRef]
- Sandrelli, F.; Tauber, E.; Pegoraro, M.; Mazzotta, G.; Cisotto, P.; Landskron, J.; Stanewsky, R.; Piccin, A.; Rosato, E.; Zordan, M.; et al. A Molecular Basis for Natural Selection at the timeless Locus in Drosophila melanogaster. Science 2007, 316, 1898–1900. [Google Scholar] [CrossRef] [PubMed]
- Machado, H.E.; Bergland, A.O.; O’Brien, K.R.; Behrman, E.L.; Schmidt, P.S.; Petrov, D.A. Comparative Population Genomics of Latitudinal Variation in Drosophila simulans and Drosophila melanogaster. Mol. Ecol. 2016, 25, 723–740. [Google Scholar] [CrossRef] [PubMed]
- Fabian, D.K.; Kapun, M.; Nolte, V.; Kofler, R.; Schmidt, P.S.; Schlötterer, C.; Flatt, T. Genome-Wide Patterns of Latitudinal Differentiation among Populations of Drosophila melanogaster from North America. Mol. Ecol. 2012, 21, 4748–4769. [Google Scholar] [CrossRef] [PubMed]
- Houle, D.; Govindaraju, D.R.; Omholt, S. Phenomics: The next Challenge. Nat. Rev. Genet. 2010, 11, 855–866. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.M.; Marques, J.G.; Sugino, K.; Wei, D.; Miyares, R.L.; Lee, T. CAMIO: A Transgenic CRISPR Pipeline to Create Diverse Targeted Genome Deletions in Drosophila. Nucleic Acids Res. 2020, 48, 4344–4356. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Genomic Region | Total SNPs | Clinal SNP |
|---|---|---|---|
| Clk | 3L: 7,763,233–7,775,603 | 1172 | 18 |
| cry | 3R: 19,212,154–19,215,442 | 488 | 37 |
| cwo | 3R: 10,388,258–10,400,752 | 751 | 25 |
| cyc | 3L: 19,813,770–19,815,938 | 302 | 18 |
| dbt | 3R: 31,054,085–31,061,210 | 727 | 49 |
| 3R: 26,455,462–26,456,141 | 311 | 18 | |
| Pdfr | X: 2,552,206–2,578,640 | 685 | 18 |
| per | X: 2,685,580–2,692,780 | 283 | 7 |
| sgg | X: 2,633,952–2,679,553 | 1110 | 5 |
| tim | 2L: 3,493,986–3,508,119 | 1864 | 36 |
| vri | 2L: 5,288,944–5,311,223 | 2083 | 45 |
| Gene | SNP Position | Alleles | Annotation | p Value |
|---|---|---|---|---|
| Clk | 3L: 7,765,472 | G/T | missense_variant | 0.02 |
| Clk | 3L: 7,757,650 | C/T | Non-synonymous (missense) | a |
| dbt | 3R: 31,056,569 | G/T | missense_variant | 0.018 |
| Eya | 2L: 6,511,849 | A/G | upstream_gene_variant | a |
| Pdfr | X: 2,577,165 | A/T | intron_variant | 0.04 |
| per | X: 2,687,600 | T/A | intron_variant | 0.051 |
| sgg | X: 2,643,833 | C/A | intron_variant | 0.03 |
| sgg | X: 2,643,812 | A/G | upstream_gene_variant | 0.03 |
| tim | 2L: 3,504,740 | T/A | intron_variant | 0.014 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khatib, L.; Subasi, B.S.; Fishman, B.; Kapun, M.; Tauber, E. Unveiling Subtle Geographical Clines: Phenotypic Effects and Dynamics of Circadian Clock Gene Polymorphisms. Biology 2023, 12, 858. https://doi.org/10.3390/biology12060858
Khatib L, Subasi BS, Fishman B, Kapun M, Tauber E. Unveiling Subtle Geographical Clines: Phenotypic Effects and Dynamics of Circadian Clock Gene Polymorphisms. Biology. 2023; 12(6):858. https://doi.org/10.3390/biology12060858
Chicago/Turabian StyleKhatib, Loren, Bengisu Sezen Subasi, Bettina Fishman, Martin Kapun, and Eran Tauber. 2023. "Unveiling Subtle Geographical Clines: Phenotypic Effects and Dynamics of Circadian Clock Gene Polymorphisms" Biology 12, no. 6: 858. https://doi.org/10.3390/biology12060858
APA StyleKhatib, L., Subasi, B. S., Fishman, B., Kapun, M., & Tauber, E. (2023). Unveiling Subtle Geographical Clines: Phenotypic Effects and Dynamics of Circadian Clock Gene Polymorphisms. Biology, 12(6), 858. https://doi.org/10.3390/biology12060858