

The Promoter SNPs Were Associated with Both the Contents of Poly-Unsaturated Fatty Acids (PUFAs) and the Expressions of PUFA-Related Genes in Common Carp

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling and Quantifying PUFA Content
2.2. Amplifying and Sequencing the Promoters of Common Carp fads2a, fads2b, elovl5a, and elovl5b
2.3. Genotyping and Estimating the Genetic Diversities
2.4. Association between the pSNPs and the PUFA Contents
2.5. Joint Effect of the pSNPs and cSNPs of elovl5b and fads2b on the PUFA Contents
2.6. Correlation between the Expression Level of Each Gene and the PUFA Content
3. Results
3.1. The Genetic Diversities of the Promoters of fads2a, fads2b, elovl5a, and elovl5b
3.2. Associations between the pSNPs with Common Carp PUFA Contents
3.3. Joint Effect of pSNPs and cSNPs from fads2b and elovl5b on the PUFA Contents
3.4. The Correlation between the PUFA Contents and the Expression Levels of Four Genes
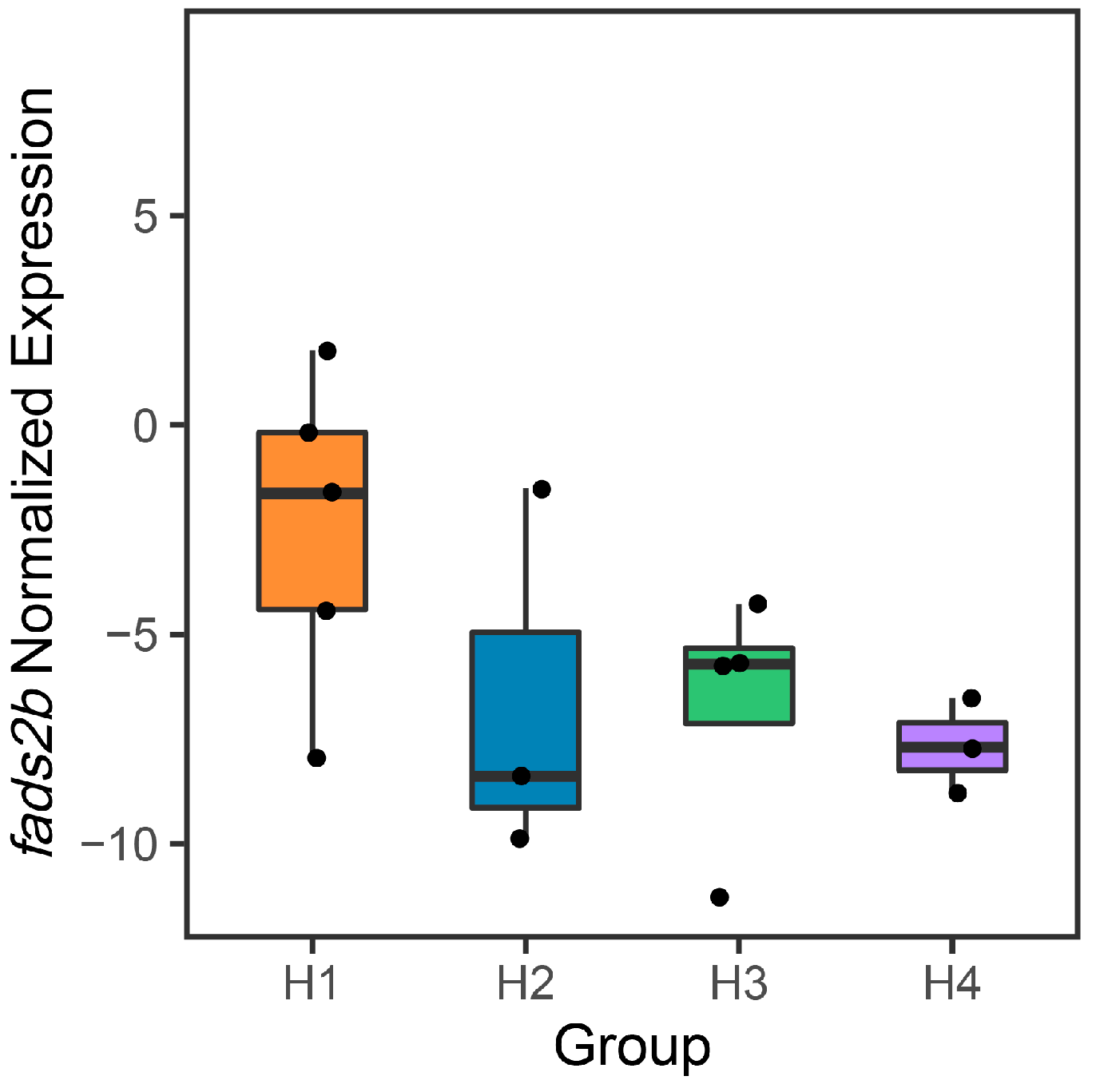
3.5. The Association of pSNPs on the fads2b Expression Levels
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hoppenbrouwers, T.; Hogervorst, J.H.C.; Garssen, J.; Wichers, H.J.; Willemsen, L.E.M. Long chain polyunsaturated fatty acids (LCPUFAs) in the prevention of food allergy. Front. Immunol. 2019, 10, 1118. [Google Scholar] [CrossRef]
- Soo, H.J.; Sam, K.K.; Chong, J.; Lau, N.S.; Ting, S.Y.; Kuah, M.K.; Kwang, S.Y.; Ranjani, M.; Shu-Chien, A.C. Functional characterisation of fatty acyl desaturase, Fads2, and elongase, Elovl5, in the Boddart’s goggle-eyed goby Boleophthalmus boddarti (Gobiidae) suggests an incapacity for long-chain polyunsaturated fatty acid biosynthesis. J. Fish Biol. 2020, 97, 83–99. [Google Scholar] [CrossRef] [PubMed]
- Xie, D.; Chen, C.; Dong, Y.; You, C.; Wang, S.; Monroig, O.; Tocher, D.R.; Li, Y. Regulation of long-chain polyunsaturated fatty acid biosynthesis in teleost fish. Prog. Lipid Res. 2021, 82, 101095. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Bunn, S.E.; Brett, M.T.; Kainz, M.J. Polyunsaturated fatty acids in stream food webs-high dissimilarity among producers and consumers. Freshw. Biol. 2017, 62, 1325–1334. [Google Scholar] [CrossRef] [Green Version]
- Badoni, P.; Nazir, I.; Aier, M.; Maity, P.B.; Samanta, S.; Das, A. Significant Role of Fish Nutrients with Special Emphasis to Essential Fatty Acid in Human Nutrition. Int. J. Curr. Microbiol. Appl. Sci. 2021, 10, 2034–2046. [Google Scholar]
- Montero, D.; Grasso, V.; Izquierdo, M.S.; Ganga, R.; Real, F.; Tort, L.; Caballero, M.J.; Acosta, F. Total substitution of fish oil by vegetable oils in gilthead sea bream (Sparus aurata) diets: Effects on hepatic Mx expression and some immune parameters. Fish Shellfish Immunol. 2008, 24, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Nasopoulou, C.; Zabetakis, I. Benefits of fish oil replacement by plant originated oils in compounded fish feeds. A review. LWT-Food Sci. Technol. 2012, 47, 217–224. [Google Scholar] [CrossRef]
- Monroig, O.; Tocher, D.R.; Castro, L.F.C. Chapter 3—Polyunsaturated fatty acid biosynthesis and metabolism in fish. In Polyunsaturated Fatty Acid Metabolism; Burdge, G.C., Ed.; Academic Press and AOCS Press: Cambridge, MA, USA, 2018; pp. 31–60. [Google Scholar]
- Tocher, D.R. Fatty acid requirements in ontogeny of marine and freshwater fish. Aquac. Res. 2010, 41, 717–732. [Google Scholar] [CrossRef]
- Castro, L.F.C.; Tocher, D.R.; Monroig, O. Long-chain polyunsaturated fatty acid biosynthesis in chordates: Insights into the evolution of Fads and Elovl gene repertoire. Prog. Lipid Res. 2016, 62, 25–40. [Google Scholar] [CrossRef]
- Monroig, O.; Tocher, D.R.; Navarro, J.C. Biosynthesis of polyunsaturated fatty acids in marine invertebrates: Recent advances in molecular mechanisms. Mar. Drugs 2013, 11, 3998–4018. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, M.T.; Nara, T.Y. Structure, function, and dietary regulation of delta6, delta5, and delta9 desaturases. Annu. Rev. Nutr. 2004, 24, 345–376. [Google Scholar] [CrossRef] [PubMed]
- Tinti, E.; Geay, F.; Lopes Rodrigues, M.; Kestemont, P.; Perpète, E.A.; Michaux, C. Molecular cloning and 3D model of a fatty-acid elongase in a carnivorous freshwater teleost, the European perch (Perca fluviatilis). 3 Biotech 2019, 9, 242. [Google Scholar] [CrossRef] [PubMed]
- Lattka, E.; Eggers, S.; Moeller, G.; Heim, K.; Weber, M.; Mehta, D.; Prokisch, H.; Illig, T.; Adamski, J. A common FADS2 promoter polymorphism increases promoter activity and facilitates binding of transcription factor ELK1. J. Lipid Res. 2010, 51, 182–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto, H.; Nogi, T.; Tabuchi, I.; Oyama, K.; Mannen, H.; Sasazaki, S. The SNPs in the promoter regions of the bovine FADS2 and FABP4 genes are associated with beef quality traits. Livest. Sci. 2014, 163, 34–40. [Google Scholar] [CrossRef]
- Pegolo, S.; Cecchinato, A.; Casellas, J.; Conte, G.; Mele, M.; Schiavon, S.; Bittante, G. Genetic and environmental relationships of detailed milk fatty acids profile determined by gas chromatography in Brown Swiss cows. J. Dairy Sci. 2016, 99, 1315–1330. [Google Scholar] [CrossRef] [Green Version]
- Proskura, W.S.; Liput, M.; Zaborski, D.; Sobek, Z.; Yu, Y.H.; Cheng, Y.H.; Dybus, A. The effect of polymorphism in the FADS2 gene on the fatty acid composition of bovine milk. Arch. Anim. Breed. 2019, 62, 547–555. [Google Scholar] [CrossRef] [Green Version]
- Gol, S.; Pena, R.N.; Rothschild, M.F.; Tor, M.; Estany, J. A polymorphism in the fatty acid desaturase-2 gene is associated with the arachidonic acid metabolism in pigs. Sci. Rep. 2018, 8, 14336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turkmen, S.; Perera, E.; Zamorano, M.J.; Simo-Mirabet, P.; Xu, H.L.; Perez-Sanchez, J.; Izquierdo, M. Effects of dietary lipid composition and fatty acid desaturase 2 expression in broodstock Gilthead sea bream on lipid metabolism-related genes and methylation of the fads2 gene promoter in Their offspring. Int. J. Mol. Sci. 2019, 20, 6250. [Google Scholar] [CrossRef] [Green Version]
- Peng, X.; Jiongtang, L.; Yan, L.; Runzi, C.; Jintu, W.; Jian, W.; Yan, Z.; Zixia, Z.; Xiaowen, S. Genomic insight into the common carp (Cyprinus carpio) genome by sequencing analysis of BAC-end sequences. BMC Genom. 2011, 12, 188. [Google Scholar]
- Grela, E.R.; Pisarski, R.K.; Kowalczuk-Vasilev, E.; Rudnicka, A. Content of nutrients and minerals, and fatty acid profile in some fish flesh depending on fishing period. Zywn.-Nauk Technol. Jakosc 2010, 17, 63–72. [Google Scholar] [CrossRef]
- Sobczak, M.; Panicz, R.; Eljasik, P.; Sadowski, J.; Tórz, A.; Żochowska-Kujawska, J.; Barbosa, V.; Domingues, V.; Marques, A.; Dias, J. Quality improvement of common carp (Cyprinus carpio L.) meat fortified with n-3 PUFA. Food Chem. Toxicol. 2020, 139, 111261. [Google Scholar] [CrossRef]
- Li, J.T.; Wang, Q.; Yang, M.D.; Li, Q.S.; Cui, M.S.; Dong, Z.J.; Wang, H.W.; Yu, J.H.; Zhao, Y.J.; Yang, C.R.; et al. Parallel subgenome structure and divergent expression evolution of allo-tetraploid common carp and goldfish. Nat. Genet. 2021, 53, 1493–1503. [Google Scholar] [CrossRef]
- Zhang, Y.; Sun, X.; Ye, Y.; Wang, Q.; Li, Q.; Zhao, R.; Wang, H.; JiongTang, L. Association between the polymorphisms of fads2a and fads2b and poly-unsaturated fatty acids in Common carp (Cyprinus carpio). Animals 2021, 11, 1780. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, Q.S.; Ye, Y.Q.; Wang, Q.; Sun, X.Q.; Zhao, R.; Li, J.T. Association analysis between genetic variants of elovl5a and elovl5b and poly-unsaturated fatty acids in Common carp (Cyprinus carpio). Biology 2022, 11, 466. [Google Scholar] [CrossRef]
- Akhtar, J.; Kreim, N.; Marini, F.; Mohana, G.; Brüne, D.; Binder, H.; Roignant, J.Y. Promoter-proximal pausing mediated by the exon junction complex regulates splicing. Nat. Commun. 2019, 10, 521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.S.; Lee, C.; Wu, C.C.; Shiue, Y.L. Primer design using genetic algorithm. Bioinformatics 2004, 20, 1710–1717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weckx, S.; Del-Favero, J.; Rademakers, R.; Claes, L.; Cruts, M.; De Jonghe, P.; Van Broeckhoven, C.; De Rijk, P. novoSNP, a novel computational tool for sequence variation discovery. Genome Res. 2005, 15, 436–442. [Google Scholar] [CrossRef] [Green Version]
- Rousset, F. genepop’007: A complete re-implementation of the genepop software for Windows and Linux. Mol. Ecol. Resour. 2008, 8, 103–106. [Google Scholar] [CrossRef]
- Nagy, S.; Poczai, P.; Cernák, I.; Gorji, A.M.; Hegedűs, G.; Taller, J. PICcalc: An online program to calculate polymorphic information content for molecular genetic studies. Biochem. Genet. 2012, 50, 670–672. [Google Scholar] [CrossRef] [Green Version]
- Tsunoda, T.; Takagi, T. Estimating transcription factor bindability on DNA. Bioinformatics 1999, 15, 622–630. [Google Scholar] [CrossRef] [Green Version]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.; Daly, M.J.; et al. PLINK: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2011; ISBN 3-900051-07-0. [Google Scholar]
- Bombail, V.; Taylor, K.; Gibson, G.G.; Plant, N. Role of Sp1, C/EBP alpha, HNF3, and PXR in the basal- and xenobiotic-mediated regulation of the CYP3A4 gene. Drug Metab. Dispos. 2004, 32, 525–535. [Google Scholar] [CrossRef]
- Xie, D.Z.; Fu, Z.X.; Wang, S.Q.; You, C.H.; Monroig, O.; Tocher, D.R.; Li, Y.Y. Characteristics of the fads2 gene promoter in marine teleost Epinephelus coioides and role of Sp1-binding site in determining promoter activity. Sci. Rep. 2018, 8, 5305. [Google Scholar] [CrossRef] [Green Version]
- Schübel, R.; Jaudszus, A.; Krüger, R.; Roth, A.; Klempt, M.; Barth, S.W. Dietary essential α-linolenic acid and linoleic acid differentially modulate TNFα-induced NFκB activity in FADS2-deficient HEK-293 cells. Int. J. Food Sci. Nutr. 2017, 68, 553–559. [Google Scholar] [CrossRef]
- Ren, H.-T.; Huang, Y.; Tang, Y.-K.; Yu, J.-H.; Xu, P. Two Elovl5-like elongase genes in Cyprinus carpio var. Jian: Gene characterization, mRNA expression, and nutritional regulation. Mol. Biol. 2015, 49, 592–600. [Google Scholar] [CrossRef]
- Zhao, R.; Wang, Y.X.; Yang, C.R.; Li, S.Q.; Li, J.C.; Sun, X.Q.; Wang, H.W.; Wang, Q.; Zhang, Y.; Li, J.T. Dominant Elongase Activity of Elovl5a but Higher Expression of Elovl5b in Common Carp (Cyprinus carpio). Int. J. Mol. Sci. 2022, 23, 14666. [Google Scholar] [CrossRef]
- Li, Y.Y.; Zhao, J.H.; Dong, Y.W.; Yin, Z.Y.; Li, Y.; Liu, Y.; You, C.H.; Monroig, O.; Tocher, D.R.; Wang, S.Q. Sp1 is involved in vertebrate LC-PUFA biosynthesis by upregulating the expression of liver desaturase and elongase genes. Int. J. Mol. Sci. 2019, 20, 5066. [Google Scholar] [CrossRef] [Green Version]
- Nwankwo, J.O.; Spector, A.A.; Domann, F.E. A nucleotide insertion in the transcriptional regulatory region of FADS2 gives rise to human fatty acid delta-6-desaturase deficiency. J. Lipid Res. 2003, 44, 2311–2319. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Li, J.; Li, Y.; Du, J.; Zhao, N.; Mai, K.; Ai, Q. Regulation of Δ6Fads2 gene involved in LC-PUFA biosynthesis subjected to fatty acid in large yellow croaker (Larimichthys crocea) and rainbow trout (Oncorhynchus mykiss). Biomolecules 2022, 12, 659. [Google Scholar] [CrossRef]
- Ching, T.; Huang, S.; Garmire, L.X. Power analysis and sample size estimation for RNA-Seq differential expression. RNA 2014, 20, 1684–1696. [Google Scholar] [CrossRef] [Green Version]
- Bottomly, D.; Walter, N.A.; Hunter, J.E.; Darakjian, P.; Kawane, S.; Buck, K.J.; Searles, R.P.; Mooney, M.; McWeeney, S.K.; Hitzemann, R. Evaluating gene expression in C57BL/6J and DBA/2J mouse striatum using RNA-Seq and microarrays. PLoS ONE 2011, 6, e17820. [Google Scholar] [CrossRef] [Green Version]
- Tian, H.M.; Yu, H.T.; Lin, Y.Q.; Li, Y.T.; Xu, W.H.; Chen, Y.R.; Liu, G.L.; Xie, L. Association between FADS gene expression and polyunsaturated fatty acids in breast milk. Nutrients 2022, 14, 457. [Google Scholar] [CrossRef]
- Xu, H.L.; Ferosekhan, S.; Turkmen, S.; Afonso, J.M.; Zamorano, M.J.; Izquierdo, M. Influence of parental fatty acid desaturase 2 (fads2) expression and diet on Gilthead seabream (Sparus aurata) offspring fads2 expression during ontogenesis. Animals 2020, 10, 2191. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| SNP | Ref/Alt | Ho | He | PIC | MAF | Genotype | Fre. Geno./No. |
|---|---|---|---|---|---|---|---|
| E5a.-122 | A/G | 0.115 | 0.216 | 0.193 | 0.123 | AA | 0.0661/15 |
| AG | 0.1145/26 | ||||||
| GG | 0.8194/186 | ||||||
| E5a.-257 | C/T | 0.211 | 0.229 | 0.203 | 0.132 | CC | 0.7621/173 |
| CT | 0.2115/48 | ||||||
| TT | 0.0264/6 | ||||||
| E5a.-264 | A/T | 0.458 | 0.436 | 0.341 | 0.322 | AA | 0.4493/102 |
| AT | 0.4581/104 | ||||||
| TT | 0.0925/21 | ||||||
| E5a.-351 | C/T | 0.401 | 0.5 | 0.375 | 0.487 | CC | 0.3128/71 |
| CT | 0.4009/91 | ||||||
| TT | 0.2863/65 | ||||||
| E5a.-430 | G/T | 0.075 | 0.088 | 0.084 | 0.046 | GG | 0.0088/2 |
| GT | 0.0749/17 | ||||||
| TT | 0.9163/208 | ||||||
| E5b.-330 | A/T | 0.208 | 0.186 | 0.169 | 0.104 | AA | 0.7921/160 |
| AT | 0.2079/42 | ||||||
| E5b.-521 | G/T | 0.075 | 0.072 | 0.069 | 0.037 | GG | 0.9254/186 |
| GT | 0.0746/15 | ||||||
| F2a.-199 | C/G | 0.406 | 0.365 | 0.299 | 0.241 | CC | 0.0377/8 |
| CG | 0.4057/86 | ||||||
| GG | 0.5566/118 | ||||||
| F2b.-99 | G/T | 0.066 | 0.119 | 0.112 | 0.064 | GG | 0.9035/206 |
| GT | 0.0658/15 | ||||||
| TT | 0.0307/7 | ||||||
| F2b.-104 | A/T | 0.339 | 0.492 | 0.371 | 0.438 | AA | 0.2687/61 |
| AT | 0.3392/77 | ||||||
| TT | 0.3921/89 | ||||||
| F2b.-304 | C/T | 0.246 | 0.306 | 0.259 | 0.189 | CC | 0.6886/157 |
| CT | 0.2456/56 | ||||||
| TT | 0.0658/15 | ||||||
| F2b.-323 | A/C | 0.145 | 0.149 | 0.138 | 0.081 | AA | 0.0088/2 |
| AC | 0.1447/33 | ||||||
| CC | 0.8465/193 | ||||||
| F2b.-609 | A/G | 0.085 | 0.174 | 0.159 | 0.096 | AA | 0.0538/12 |
| AG | 0.0852/19 | ||||||
| GG | 0.861/192 | ||||||
| F2b.-650 | A/C | 0.049 | 0.114 | 0.107 | 0.061 | AA | 0.9148/204 |
| AC | 0.0493/11 | ||||||
| CC | 0.0359/8 | ||||||
| F2b.-700 | G/T | 0.126 | 0.178 | 0.162 | 0.099 | GG | 0.8386/187 |
| GT | 0.1256/28 | ||||||
| TT | 0.0359/8 | ||||||
| F2b.-721 | G/T | 0.143 | 0.171 | 0.156 | 0.094 | GG | 0.8341/186 |
| GT | 0.1435/32 | ||||||
| TT | 0.0224/5 | ||||||
| F2b.-744 | G/T | 0.139 | 0.174 | 0.159 | 0.096 | GG | 0.8341/186 |
| GT | 0.139/31 | ||||||
| TT | 0.0269/6 | ||||||
| F2b.-785 | A/T | 0.363 | 0.496 | 0.373 | 0.455 | AA | 0.3632/81 |
| AT | 0.3632/81 | ||||||
| TT | 0.2735/61 | ||||||
| F2b.-880 | C/T | 0.137 | 0.41 | 0.326 | 0.288 | CC | 0.2192/48 |
| CT | 0.137/30 | ||||||
| TT | 0.6438/141 |
| Trait | SNP | GLM (Permutation) | ANOVA | ||||
|---|---|---|---|---|---|---|---|
| PUFA | ID | p Value | Marker PVE (%) | Genotype | p Value (FDR) | ||
| MM | Mm | mm | |||||
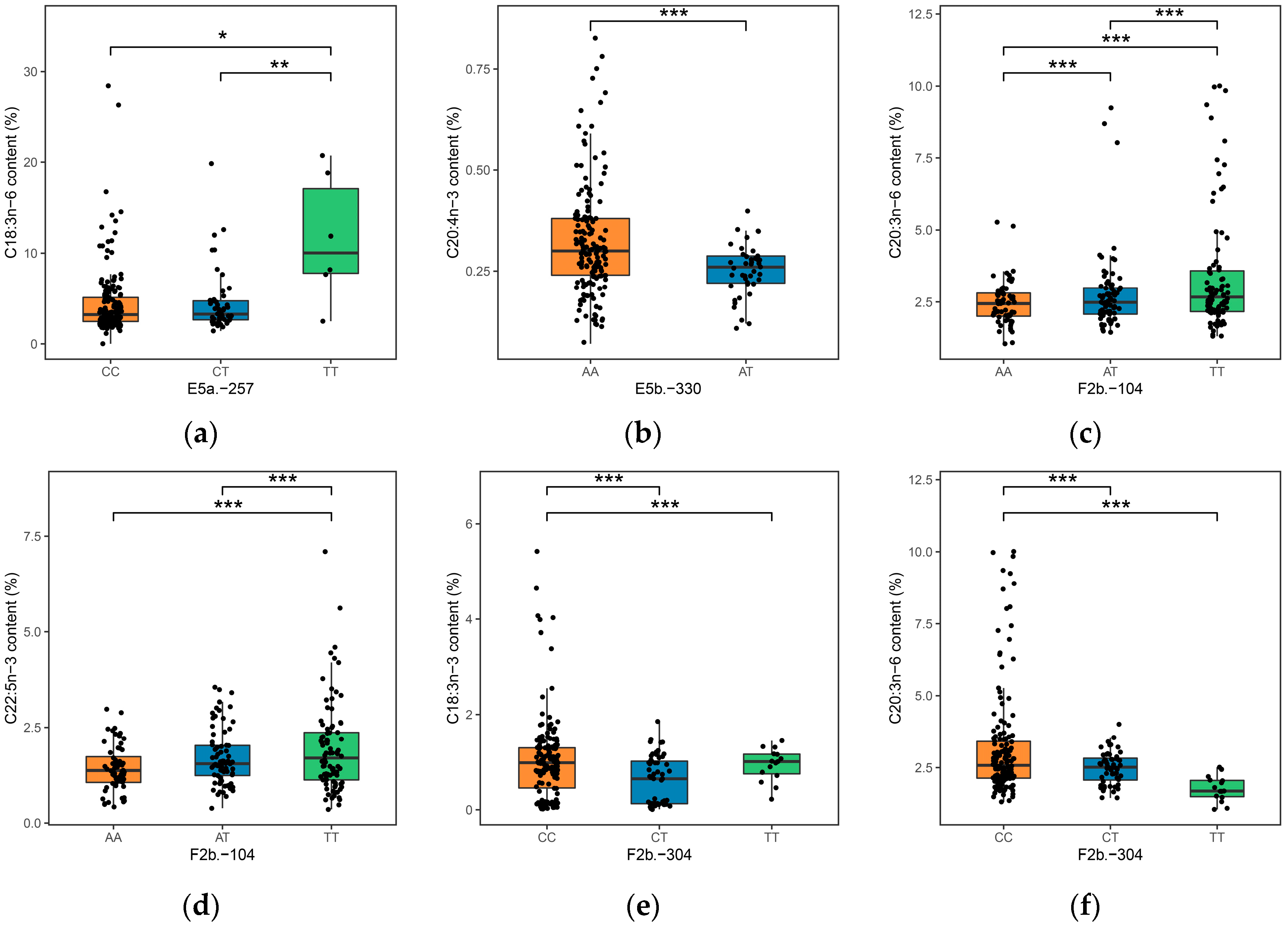
| C18:3n-6 | E5a.-257 | 6.12 × 10−5 | 8.55% | 4.51 (CC) | 4.53 (CT) | 11.63 (TT) | 1.16 × 10−3 |
| C20:4n-3 | E5b.-330 | 2.48 × 10−3 | 4.53% | 0.32 (AA) | 0.25 (AT) | -- | 4.7 × 10−2 |
| C20:3n-6 | F2b.-104 | 1.75 × 10−3 | 5.66% | 3.4 (TT) | 2.79 (AT) | 2.46 (AA) | 3.14 × 10−2 |
| C22:5n-3 | F2b.-104 | 5.16 × 10−3 | 4.74% | 1.94 (TT) | 1.72 (AT) | 1.44 (AA) | 9.29 × 10−2 |
| C18:3n-3 | F2b.-304 | 1.26 × 10−2 | 3.92% | 1.04 (CC) | 0.61 (CT) | 0.94 (TT) | 5.1 × 10−2 |
| C20:3n-6 | F2b.-304 | 2.92 × 10−4 | 7.16% | 3.21 (CC) | 2.5 (CT) | 1.77 (TT) | 5.54 × 10−3 |
| C18:2n-6 | F2b.-700 | 2.3 × 10−3 | 5.52% | 23.95 (GG) | 30.72 (GT) | 27.78 (TT) | 4.37 × 10−2 |
| C20:3n-3 | F2b.-785 | 3.75 × 10−4 | 7.11% | 0.3 (AA) | 0.29 (AT) | 0.21 (TT) | 7.12 × 10−3 |
| C22:4n-6 | F2b.-785 | 1.89 × 10−3 | 5.80% | 0.76 (AA) | 0.76 (AT) | 0.56 (TT) | 3.59 × 10−2 |
| C22:5n-3 | F2b.-785 | 4.85 × 10−3 | 4.88% | 1.88 (AA) | 1.84 (AT) | 1.4 (TT) | 9.22 × 10−2 |
| cpsnp Set | C18:3n-3 | C18:4n-3 | C20:3n-3 | C20:4n-3 | C20:5n-3 | C22:5n-3 |
|---|---|---|---|---|---|---|
| E5b.-330 | NA | NA | NA | 4.53% | NA | NA |
| E5b.172 | 6.9% | NA | 3.96% | 7.12% | NA | 3.7% |
| E5b.782 | NA | NA | 2.15% | NA | NA | NA |
| E5b.cpSNPs | 11.31% | 1.08% | 11.84% | 13.71% | 4.43% | 9.17% |
| F2b.-104 | NA | NA | NA | NA | NA | 4.74% |
| F2b.-304 | 3.92% | NA | NA | NA | NA | NA |
| F2b.-785 | NA | NA | 7.11% | NA | NA | 4.88% |
| F2b.751 | 5% | NA | 13% | 11% | NA | 11% |
| F2b.1197 | NA | NA | 4.3% | 5% | NA | 5% |
| F2b.cpSNPs | 21.41% | 8.69% | 20.64% | 17.91% | 12.11% | 14.42% |
| E5b.cpSNPs & F2b.cpSNPs | 18.12% | 15.74% | 31.70% | 20.64% | 20.41% | 21.79% |
| cpSNP Set | C18:2n-6 | C18:3n-6 | C20:3n-6 | C20:4n-6 | C22:4n-6 | C22:5n-6 |
|---|---|---|---|---|---|---|
| E5b.-330 | NA | NA | NA | NA | NA | NA |
| E5b.172 | 2.86% | NA | 3.37% | 2.1% | 3.44% | 4.07% |
| E5b.782 | NA | NA | 4.48% | NA | 1.83% | 1.83% |
| E5b.cpSNPs | 13.22% | 1.29% | 13.37% | 3.45% | 9.56% | 11.65% |
| F2b.-104 | NA | NA | 5.66% | NA | NA | NA |
| F2b.-304 | NA | NA | 7.16% | NA | NA | NA |
| F2b.-785 | NA | NA | NA | NA | 5.80% | NA |
| F2b.751 | NA | NA | 11% | NA | 7% | 9% |
| F2b.1197 | 4% | NA | 4% | NA | NA | 3% |
| F2b.cpSNPs | 10.08% | 9.96% | 16.99% | 8.25% | 12.63% | 14.15% |
| E5b.cpSNPs & F2b.cpSNPs | 12.37% | 9.93% | 28.65% | 30.87% | 43.74% | 35.35% |
| PUFAs (%) | fads2a | fads2b | elovl5a | elovl5b | ||||
|---|---|---|---|---|---|---|---|---|
| r | FDR p Value | R | FDR p Value | r | FDR p Value | r | FDR p Value | |
| C18:3n-3 | −0.091 | 0.558 | −0.105 | 0.498 | 0.043 | 0.784 | 0.285 | 0.061 |
| C18:4n-3 | 0.113 | 0.465 | 0.350 * | 0.02 | 0.25 | 0.102 | 0.370 * | 0.013 |
| C20:3n-3 | 0.410 * | 0.006 | 0.421 * | 0.004 | 0.169 | 0.273 | 0.023 | 0.88 |
| C20:4n-3 | 0.377 * | 0.012 | 0.380 * | 0.011 | 0.228 | 0.137 | 0.189 | 0.22 |
| C20:5n-3 | 0.410 * | 0.006 | 0.186 | 0.227 | 0.087 | 0.574 | −0.058 | 0.708 |
| C22:5n-3 | 0.456 ** | 0.002 | 0.362 * | 0.016 | 0.063 | 0.682 | −0.087 | 0.573 |
| C18:2n-6 | 0.019 | 0.9 | 0.119 | 0.443 | −0.044 | 0.778 | 0.208 | 0.175 |
| C18:3n-6 | 0.151 | 0.329 | 0.028 | 0.858 | −0.317 | 0.036 * | −0.349 | 0.02 |
| C20:3n-6 | 0.144 | 0.352 | 0.244 | 0.111 | 0.099 | 0.521 | 0.21 | 0.171 |
| C20:4n-6 | 0.354 * | 0.018 | 0.434 * | 0.003 | 0.201 | 0.191 | −0.015 | 0.924 |
| C22:4n-6 | 0.367 * | 0.014 | 0.481 ** | 0.001 | 0.181 | 0.24 | 0.068 | 0.661 |
| C22:5n-6 | 0.352 * | 0.019 | 0.458 ** | 0.002 | 0.149 | 0.334 | 0.121 | 0.435 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Xu, Z.-M.; Wang, Q.; Li, Q.-S.; Sun, X.-Q.; Li, J.-T. The Promoter SNPs Were Associated with Both the Contents of Poly-Unsaturated Fatty Acids (PUFAs) and the Expressions of PUFA-Related Genes in Common Carp. Biology 2023, 12, 524. https://doi.org/10.3390/biology12040524
Zhang Y, Xu Z-M, Wang Q, Li Q-S, Sun X-Q, Li J-T. The Promoter SNPs Were Associated with Both the Contents of Poly-Unsaturated Fatty Acids (PUFAs) and the Expressions of PUFA-Related Genes in Common Carp. Biology. 2023; 12(4):524. https://doi.org/10.3390/biology12040524
Chicago/Turabian StyleZhang, Yan, Zi-Ming Xu, Qi Wang, Qing-Song Li, Xiao-Qing Sun, and Jiong-Tang Li. 2023. "The Promoter SNPs Were Associated with Both the Contents of Poly-Unsaturated Fatty Acids (PUFAs) and the Expressions of PUFA-Related Genes in Common Carp" Biology 12, no. 4: 524. https://doi.org/10.3390/biology12040524
APA StyleZhang, Y., Xu, Z.-M., Wang, Q., Li, Q.-S., Sun, X.-Q., & Li, J.-T. (2023). The Promoter SNPs Were Associated with Both the Contents of Poly-Unsaturated Fatty Acids (PUFAs) and the Expressions of PUFA-Related Genes in Common Carp. Biology, 12(4), 524. https://doi.org/10.3390/biology12040524