
Changes in Competitors, Stress Tolerators, and Ruderals (CSR) Ecological Strategies after the Introduction of Shrubs and Trees in Disturbed Semiarid Steppe Grasslands in Hulunbuir, Inner Mongolia


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
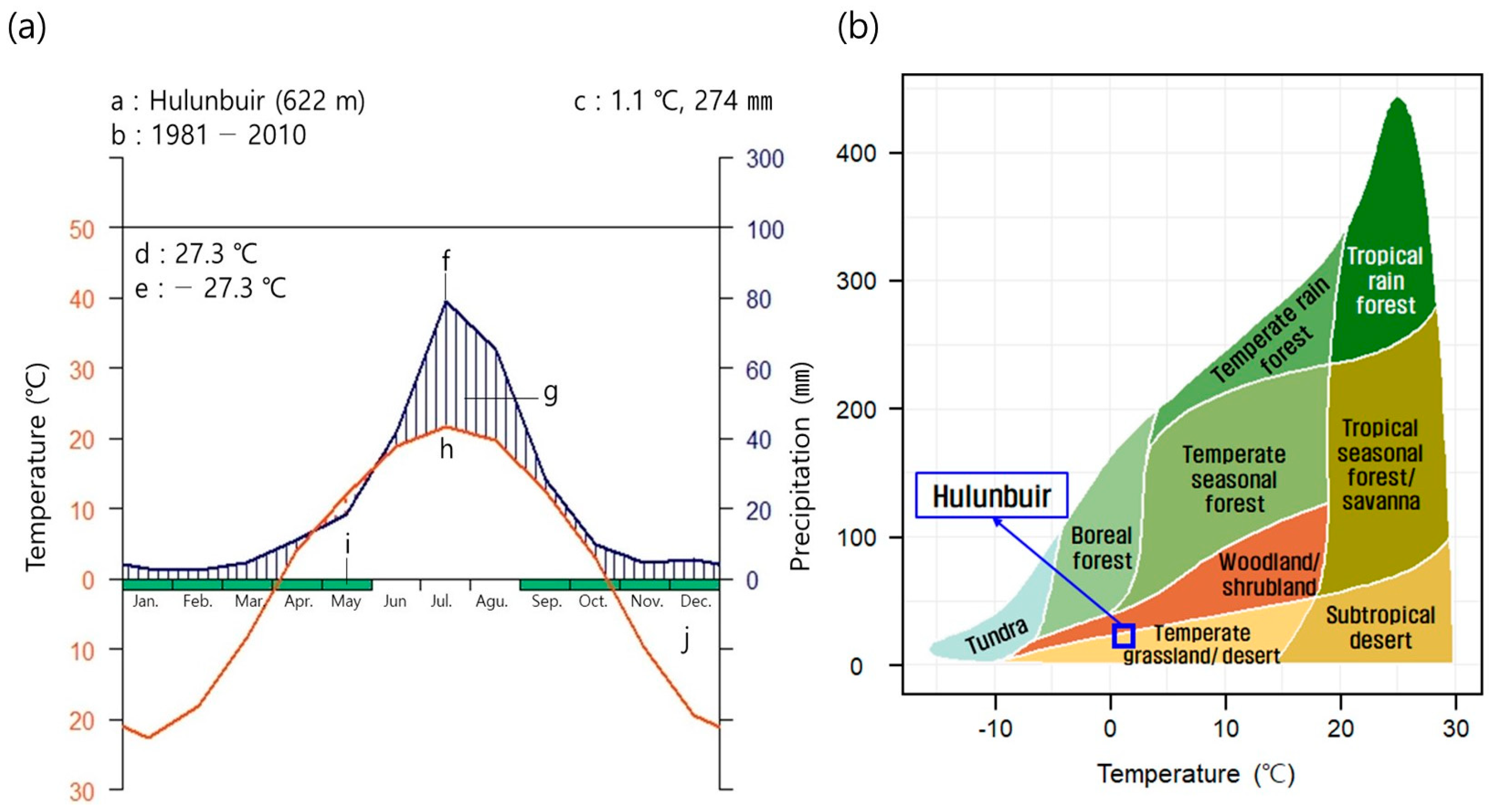
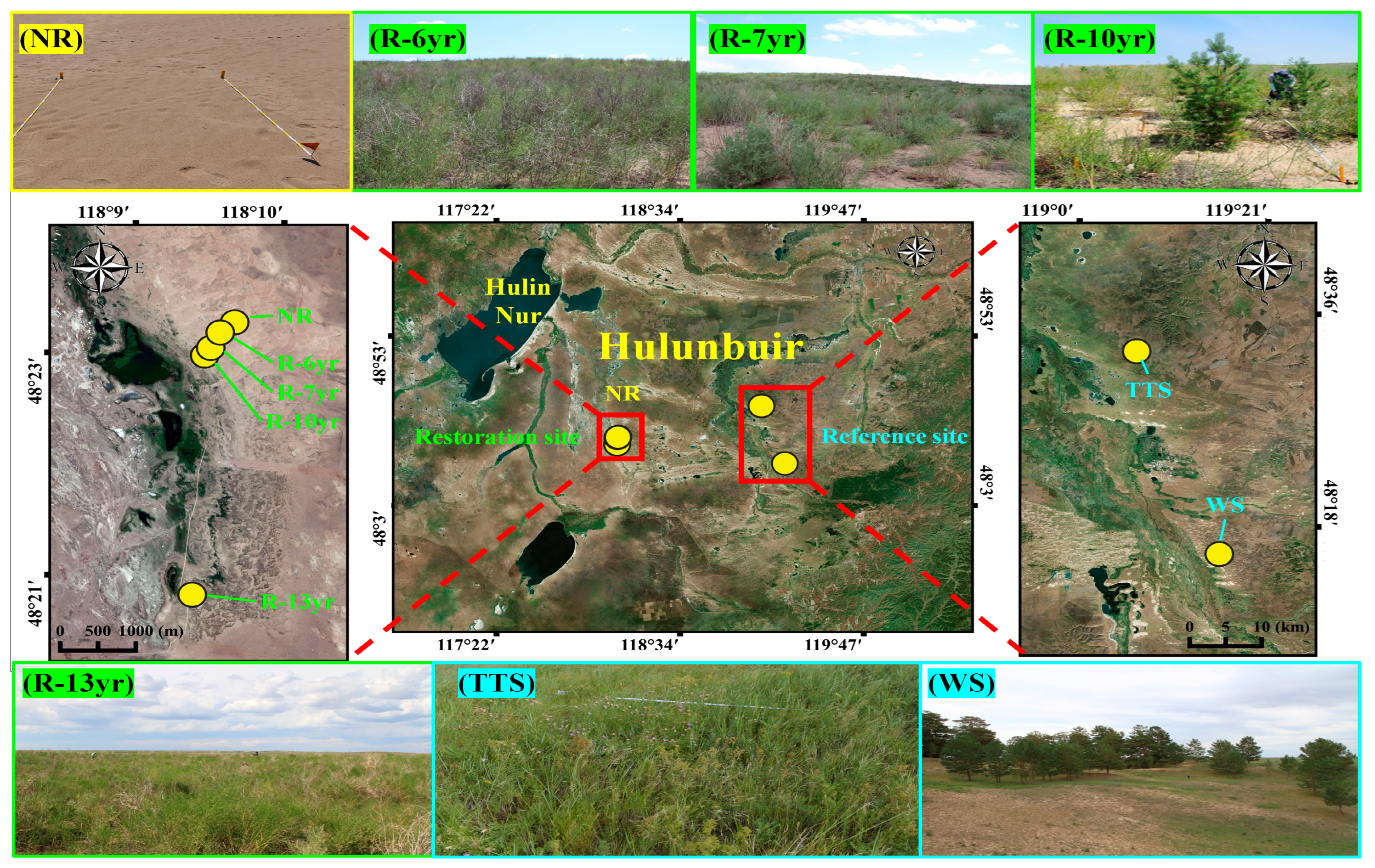
2.1. Overview of the Study Site
2.2. CSR Classification and Functional Group Diversity Comparison
2.3. Plant Species Composition and Diversity
3. Results
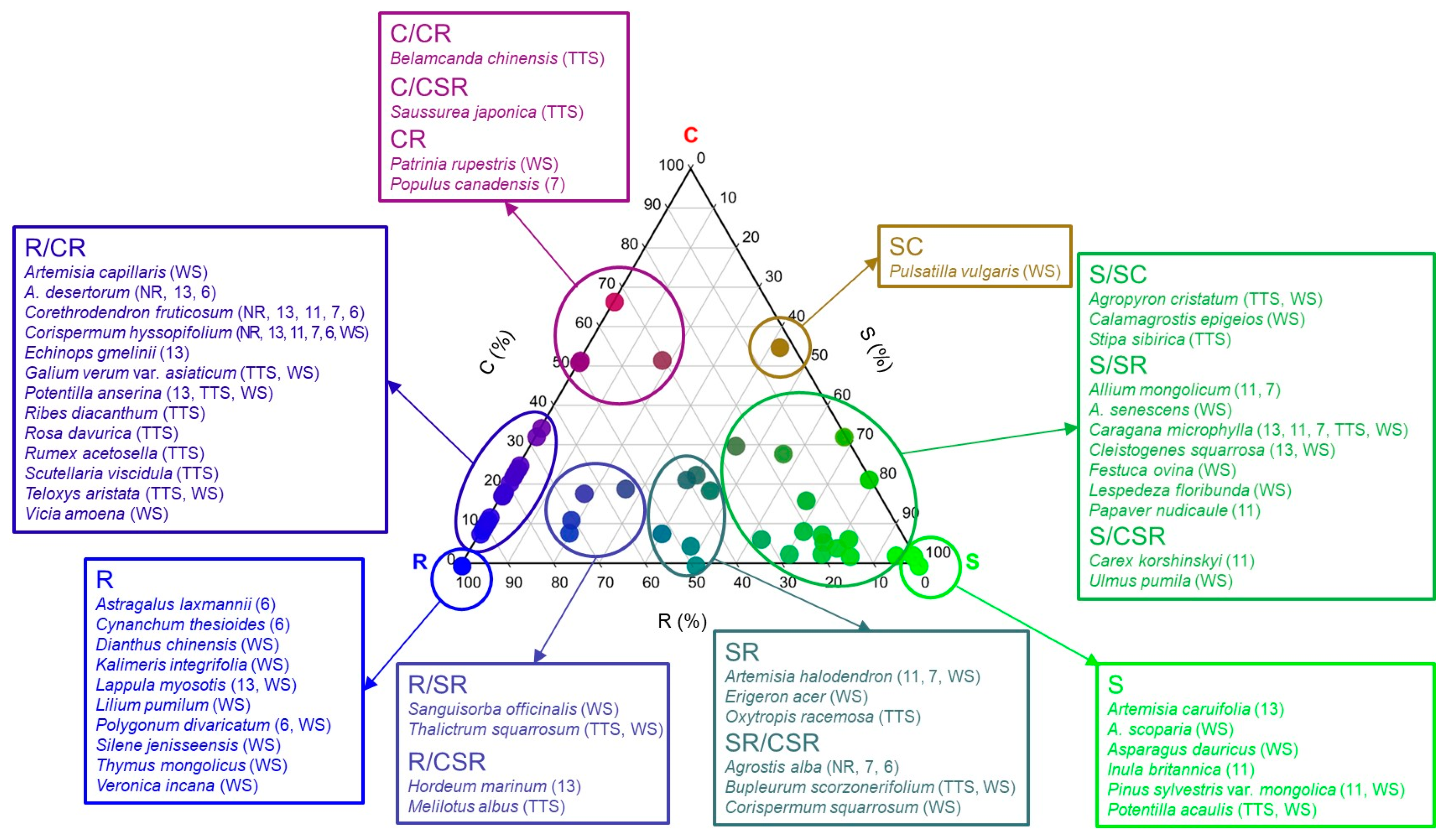
3.1. Plant CSR Ecological Strategies
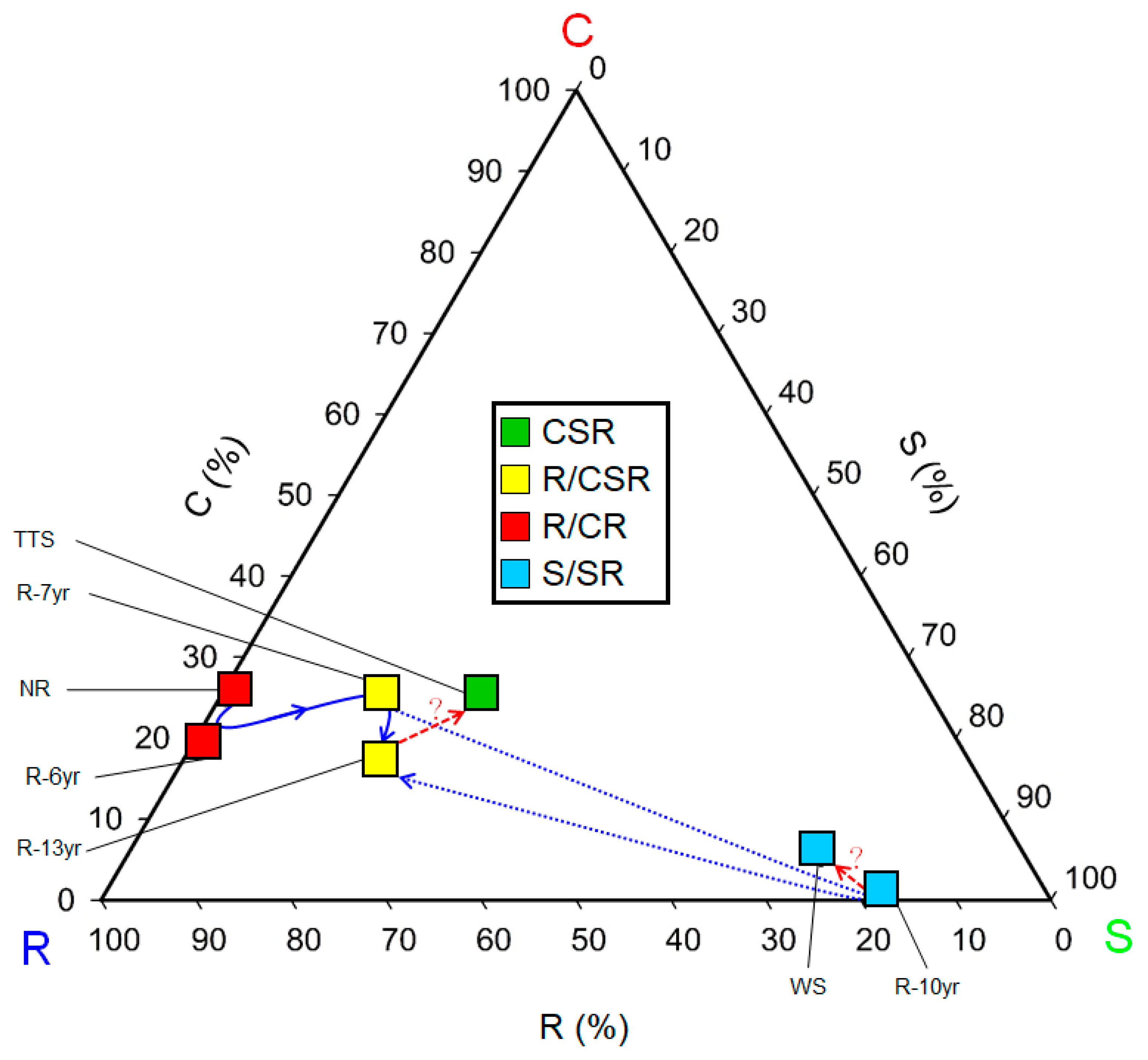
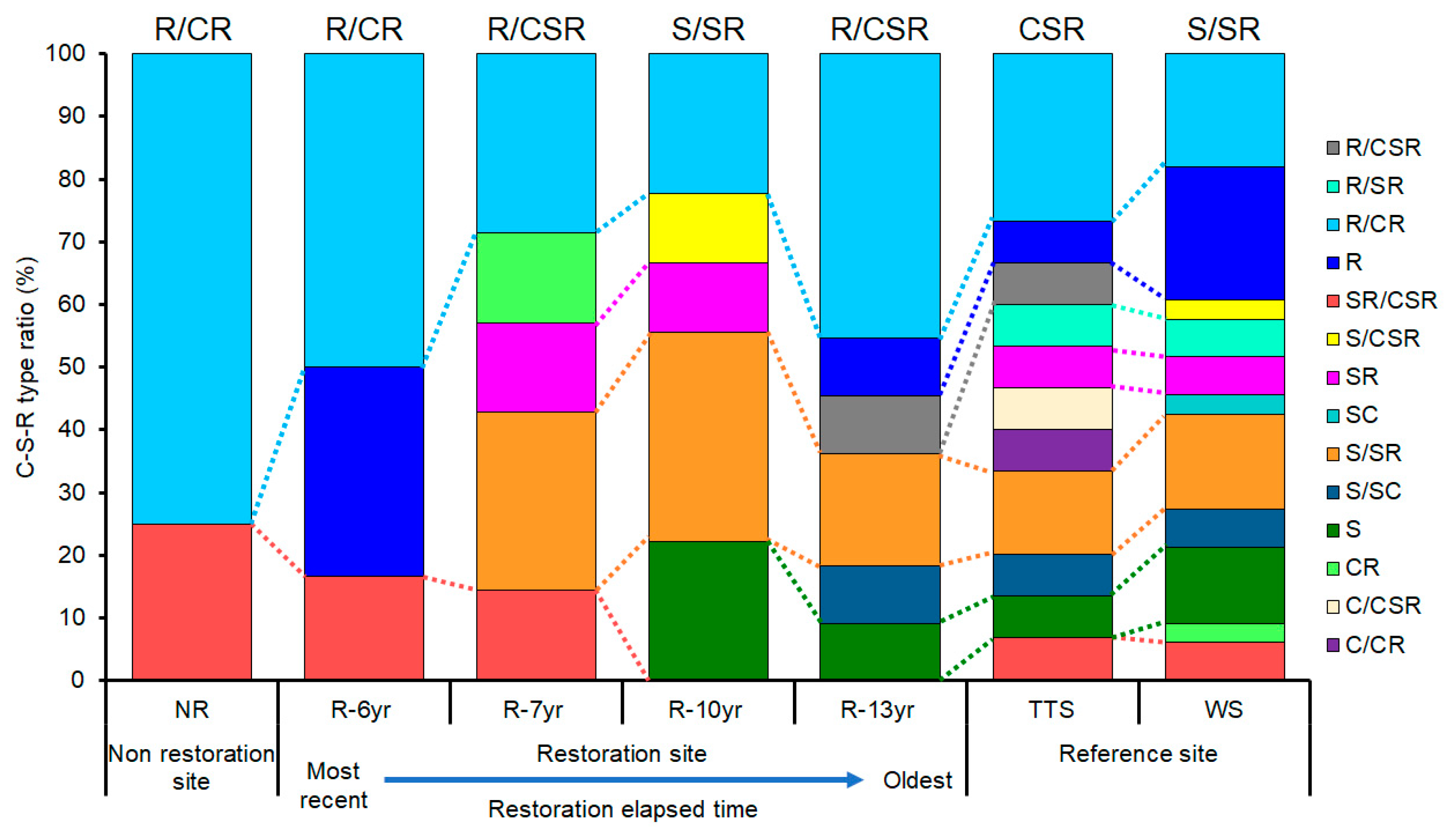
3.2. Changes in the Communities’ CSR Ecological Strategies
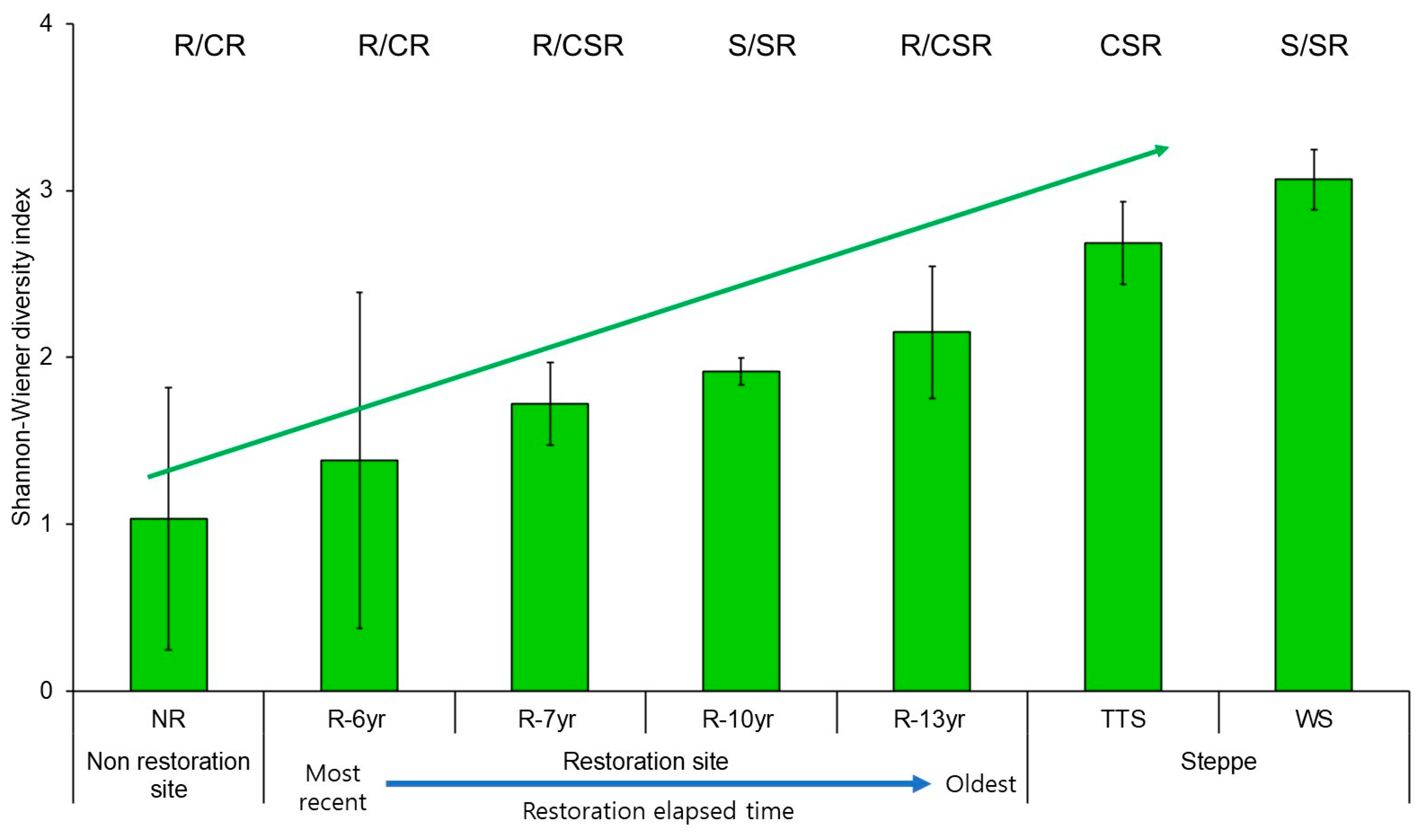
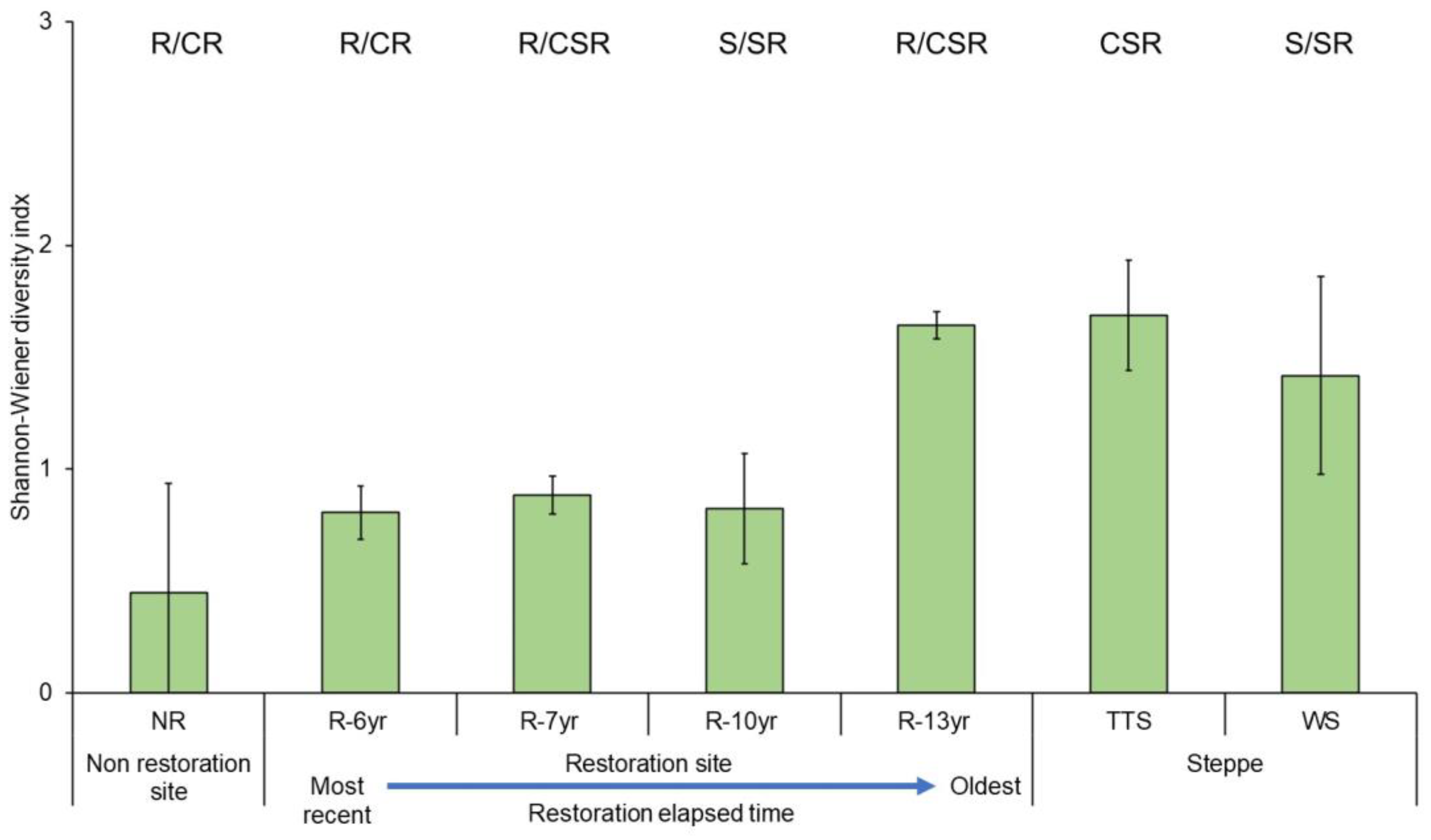
3.3. Changes in Eco-Functional Diversity
4. Discussion
4.1. Types of Plant CSR Ecological Strategies
4.2. Changes in Community CSR Ecological Strategies and Eco-Functional Diversity Following the Progress of Restoration
4.3. Changes in Plant Species Diversity According to the Progress of Restoration
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Park, B.K. Korea’s Grassland Research; Ewhapress: Seoul, Republic of Korea, 1985; 530p. [Google Scholar]
- White, R.; Murray, S.; Rohweder, M. Pilot Analysis of Global Ecosystems: Grassland Ecosystems; World Resources Institute: Washington, DC, USA, 2000. [Google Scholar]
- Shantz, H.L. The place of grasslands in the Earth’s cover. Ecology 1954, 35, 143–145. [Google Scholar] [CrossRef]
- Eyre, S.R. Vegetation and Soils: A World Picture, 2nd ed.; Routledge: London, UK, 1968. [Google Scholar]
- Angerer, J.; Han, G.; Fujisaki, I.; Havstad, K. Climate change and ecosystems of Asia with emphasis on Inner Mongolia and Mongolia. Rangelands 2008, 30, 46–51. [Google Scholar] [CrossRef]
- Wrage, N.; Strodthoff, J.; Cuchillo, H.M.; Isselstein, J.; Kayser, M. Phytodiversity of temperate permanent grasslands: Ecosystem services for agriculture and livestock management for diversity conservation. Biodivers. Conserv. 2011, 20, 3317–3339. [Google Scholar] [CrossRef]
- Li, M.; Wang, X.; Chen, J. Assessment of grassland ecosystem services and analysis on its driving factors: A case study in Hulunbuir Grassland. Front. Ecol. Evol. 2022, 10, 841943. [Google Scholar] [CrossRef]
- Blair, J.; Nippert, J.; Briggs, J. Grassland ecology 14. Ecol. Environ. 2014, 389, 389–423. [Google Scholar] [CrossRef]
- Chen, Y.; Li, Y.; Zhao, X.; Awada, T.; Shang, W.; Han, J. Effects of grazing exclusion on soil properties and on ecosystem carbon and nitrogen storage in a sandy rangeland of Inner Mongolia, Northern China. Environ. Manag. 2012, 50, 622–632. [Google Scholar] [CrossRef]
- Chen, L.; Gao, J.; Ji, Y.; Bai, Z.; Shi, M.; Liu, H. Effects of particulate matter of various sizes derived from suburban farmland, woodland and grassland on air quality of the central district in Tianjin, China. Aerosol Air Qual. Res. 2014, 14, 829–839. [Google Scholar] [CrossRef]
- Montanarella, L.; Scholes, R.; Brainich, A. The Assessment Report on Land Degradation and Restoration; Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services (IPBES): Bonn, Germany, 2018. [Google Scholar]
- Li, J.; Zheng, Z.; Xie, H.; Zhao, N.; Gao, Y. Increased soil nutrition and decreased light intensity drive species loss after eight years grassland enclosures. Sci. Rep. 2017, 7, 44525. [Google Scholar] [CrossRef]
- Deák, B.; Rádai, Z.; Lukács, K.; Kelemen, A.; Kiss, R.; Bátori, Z.; Péter János, K.; Valkó, O. Fragmented dry grasslands preserve unique components of plant species and phylogenetic diversity in agricultural landscapes. Biodivers. Conserv. 2020, 29, 4091–4110. [Google Scholar] [CrossRef]
- Oikawa, T.; Ito, A. Modeling carbon dynamics of terrestrial ecosystem in monsoon Asia. In Present and Future of Modeling Global Environmental Change: Towards Integrated Modeling; Matsuno, T., Kida, H., Eds.; Terra Scientific Publishing: Tokyo, Japan, 2001; pp. 207–219. [Google Scholar]
- Akiyama, T.; Kawamura, K. Grassland degradation in China: Methods of monitoring, management, and restoration. Grassl. Sci. 2007, 53, 1–17. [Google Scholar] [CrossRef]
- Zhang, Z.; Huisingh, D. Combating desertification in China: Monitoring, control, management and revegetation. J. Clean. Prod. 2018, 182, 765–775. [Google Scholar] [CrossRef]
- Yang, B.; Gong, J.R.; Zhang, Z.H.; Wang, B.; Zhu, C.C.; Shi, J.; Liu, M.; Liu, Y.; Li, X. Stabilization of carbon sequestration in a Chinese desert steppe benefits from increased temperatures and from precipitation outside the growing season. Sci. Total Environ. 2019, 691, 263–277. [Google Scholar] [CrossRef] [PubMed]
- Li, W.J.; Ali, S.H.; Zhang, Q. Property rights and grassland degradation: A study of the Xilingol Pasture, Inner Mongolia, China. J. Environ. Manag. 2007, 85, 461–470. [Google Scholar] [CrossRef]
- Kang, L.; Han, X.; Zhang, Z.; Sun, O.J. Grassland ecosystems in China: Review of current knowledge and research advancement. Philos. Trans. R. Soc. B Biol. Sci. 2007, 362, 997–1008. [Google Scholar] [CrossRef] [PubMed]
- Squires, V.R.; Lu, X.; Lu, Q.; Wang, T.; Yang, Y. Chapter 7: Case study1 Hulunbeier grassland, Inner Mongolia. In Rangeland Degradation and Recovery in China’s Pastoral Lands; CAB Internationl: London, UK, 2009; pp. 91–102. [Google Scholar] [CrossRef]
- Batunacun; Hu, Y.; Biligejifu; Liu, J.; Zhen, L. Spatial distribution and variety of grass species on the Ulan bator—Xilinhot transect of Mongolian plateau. J. Nat. Resour. 2015, 30, 24–36. [Google Scholar] [CrossRef]
- McIntosh, R.P. The Background of Ecology: Concept and Theory; Cambridge University Press: Cambridge, UK, 1986. [Google Scholar]
- Bai, X.; Zhao, W.; Wang, J.; Ferreira, C.S.S. Reducing plant community variability and improving resilience for sustainable restoration of temperate grassland. Environ. Res. 2022, 207, 112149. [Google Scholar] [CrossRef]
- Fry, E.L.; Savage, J.; Hall, A.L.; Oakley, S.; Pritchard, W.J.; Ostle, N.J.; Pywell, R.F.; Bullock, J.M.; Bardgett, R.D. Soil multifunctionality and drought resistance are determined by plant structural traits in restoring grassland. Ecology 2018, 99, 2260–2271. [Google Scholar] [CrossRef]
- Barbour, R.K.K.; Burk, P.; Pitts, J.; Gilliam, F.; Schwartz, M. Terrestrial Plant Ecology; Hongreung Publusing Company: Seoul, Republic of Korea, 2015; 614p. [Google Scholar]
- Hunt, R.; Hodgson, J.G.; Thompson, K.; Bungener, P.; Dunnett, N.P.; Askew, A.P. A new practical tool for deriving a functional signature for herbaceous vegetation. Appl. Veg. Sci. 2004, 7, 163–170. [Google Scholar] [CrossRef]
- Pierce, S.; Negreiros, D.; Cerabolini, B.E.L.; Kattge, J.; Díaz, S.; Kleyer, M.; Shipley, B.; Wright, S.J.; Soudzilovskaia, N.A.; Onipchenko, V.G.; et al. A global method for calculating plant CSR ecological strategies applied across biomes world-wide. Funct. Ecol. 2017, 31, 444–457. [Google Scholar] [CrossRef]
- Grime, J.P.; Pierce, S. The Evolutionary Strategies that Shape Ecosystems; Wiley-Blackwell: Chichester, UK, 2012. [Google Scholar]
- Yu, J.; Hou, G.; Zhou, T.; Shi, P.; Zong, N.; Sun, J. Variation of plant CSR strategies across a precipitation gradient in the alpine grasslands on the northern Tibet Plateau. Sci. Total Environ. 2022, 838, 156512. [Google Scholar] [CrossRef]
- Zhu, Y.; Shan, D.; Wang, B.; Shi, Z.; Yang, X.; Liu, Y. Floristic features and vegetation classification of the Hulun Buir steppe in North China: Geography and climate-driven steppe diversification. Glob. Ecol. Conserv. 2019, 20, e00741. [Google Scholar] [CrossRef]
- Walter, H.; Harnickell, E.; Mueller-Dombois, D. Climate Diagram Maps of the Individual Countries and the Ecological Climatic Regions of the Earth: Supplement to the Vegetation Monographs; Springer: Berlin, Germany, 1975; Volume 8, pp. 1–36. [Google Scholar]
- China Meteorological Data Service Centre. 2023. Available online: http://data.cma.cn/ (accessed on 1 August 2023).
- Whittaker, R.H. Classification of natural communities. Bot. Rev. 1962, 28, 1–239. [Google Scholar] [CrossRef]
- Whittaker, R.H. Communities and Ecosystems, 2nd ed.; Cornell University Press: New York, NY, USA, 1970. [Google Scholar]
- Kottek, M.; Grieser, J.; Beck, C.; Rudolf, B.; Rubel, F. World Map of the Köppen-Geiger climate classification updated. Meteorol. Z. 2006, 15, 259–263. [Google Scholar] [CrossRef]
- Park, K.H.; Qu, Z.Q.; Wan, Q.Q.; Ding, G.D.; Wu, B. Effects of enclosures on vegetation recovery and succession in Hulunbeier steppe, China. For. Sci. Technol. 2013, 9, 25–32. [Google Scholar] [CrossRef]
- Hodgson, J.G.; Wilson, P.J.; Hunt, R.; Grime, J.P.; Thompson, K. Allocation C-S-R plant functional types: A soft approach to a hard problem. Oikos 1999, 85, 282–294. [Google Scholar] [CrossRef]
- Gu, A.; Wang, Z. Atlas of Rangeland Plants in Northern China; China Agricultural Science and Technology Press: Beijing, China, 2009; 524p. [Google Scholar]
- Gu, A.; Wang, Z. Atlas of Rangeland Plants in Northern China (Supplement); China Agricultural Science and Technology Press: Beijing, China, 2011; 363p. [Google Scholar]
- Flora of China. 2023. Available online: http://www.efloras.org/ (accessed on 1 August 2023).
- Pierce, S.; Brusa, G.; Vagge, I.; Cerabolini, B.E.L. Allocations CSR plant functional types: The use of leaf economics and size traits to classify woody and herbaceous vascular plants. Funct. Ecol. 2013, 27, 1002–1010. [Google Scholar] [CrossRef]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Grime, J.P. Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory. Am. Nat. 1977, 111, 1169–1194. [Google Scholar] [CrossRef]
- Grime, J.P. Manipulation of plant species and communities. In Ecology and Design in Landscape; Bradshaw, A.D., Goode, D.A., Thorpe, E., Eds.; Blackwell Scientific Publications: Oxford/London, UK, 1986; pp. 175–194. [Google Scholar]
- Grime, J.P. Plant Strategies and Vegetation Processes and Ecosystem Properties, 2nd ed.; John Wiley and Sons: Hoboken, NJ, USA, 2001; 2011419p. [Google Scholar]
- Grime, J.P.; Hunt, R. Relative growth-rate: Its range and adaptive significance in a local flora. J. Ecol. 1975, 63, 393–422. [Google Scholar] [CrossRef]
- Tilman, D. Plant Strategies and the Dynamics and Structure of Plant Communities; Princeton University Press: Princeton, NJ, USA, 1998. [Google Scholar]
- Liu, Y.; Bao, G.; Song, H.; Cai, Q.; Sun, J. Precipitation reconstruction from Hailar pine (Pinus sylvestris var. mongolica) tree rings in the Hailar region, Inner Mongolia, China back to 1865 AD. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2009, 282, 81–87. [Google Scholar] [CrossRef]
- Song, L.; Zhu, J.; Zhang, J.; Zhang, T.; Wang, K.; Wang, G.; Liu, J. Effect of drought and topographic position on depth of soil water extraction of Pinus sylvestris L. var. mongolica Litv. trees in a semiarid sandy region, Northeast China. Forests 2019, 10, 370. [Google Scholar] [CrossRef]
- Bessle, H.; Oelmann, Y.; Roscher, C.; Buchmann, N.; Scherer-Lorenzen, M.; Schulze, E.D.; Temperton, V.M.; Wilcke, W.; Engels, C. Nitrogen uptake by grassland communities: Contribution of N2 fixation, facilitation, complementarity, and species dominance. Plant Soil 2012, 358, 301–322. [Google Scholar] [CrossRef]
- Tang, L.; Mao, L.; Shu, J.; Li, C.; Shen, C.; Zhou, Z. Atlas of Quaternary Pollen and Spores in China; Springer: Beijing, China, 2020; 577p. [Google Scholar]
- Kim, J.H.; Choi, S.S.; An, I.J.; Lee, S.H.; Lee, E.J.; You, Y.H.; Kim, B.J.; Han, D.U.; Park, S.K.; Joo, S.B. Palatability and livestock preferences of restored plants in steppe restoration areas, Hulunbuir, Inner Mongolia, China. Proc. Natl. Inst. Ecol. Repub. Korea 2021, 2, 170–179. [Google Scholar] [CrossRef]
- Pierce, S.; Vagge, I.; Brusa, G. The intimacy between sexual traits and Grime’s CSR strategies for orchids coexisting in semi-natural calcareous grassland at the Olive Lawn. Plant Ecol. 2014, 215, 495–505. [Google Scholar] [CrossRef]
- Critchley, C.N.R. Ecological assessment of plant communities by reference to species traits and habitat preferences. Biodivers. Conserv. 2000, 9, 87–105. [Google Scholar] [CrossRef]
- Caccianiga, M.; Luzzaro, A.; Pierce, S.; Ceriani, R.M.; Cerabolini, B. The functional basis of a primary succession resolved by CSR classification. Oikos 2006, 112, 10–20. [Google Scholar] [CrossRef]
- Pierce, S.; Ceriani, R.M.; De Andreis, R.; Luzzaro, A.; Cerabolini, B. The leaf economics spectrum of Poaceae reflects variation in survival strategies. Plant Biosyst. 2007, 141, 337–343. [Google Scholar] [CrossRef]
- Zhou, T.; Hou, G.; Sun, J.; Zong, N.; Shi, P. Degradation shifts plant communities from S-to R-strategy in an alpine meadow, Tibetan Plateau. Sci. Total Environ. 2021, 800, 149572. [Google Scholar] [CrossRef]
- Shimoda, S.; Mo, W.; Oikawa, T. The effects of characteristics of Asian monsoon climate on interannual CO2 exchange in a humid temperate C3/C4 co-occurring grassland. Sci. Online Lett. Atmos. 2005, 1, 169–172. [Google Scholar] [CrossRef][Green Version]
- Zhang, W. Changes in species diversity and canopy cover in steppe vegetation in Inner Mongolia under protection from grazing. Biodivers. Conserv. 1998, 7, 1365–1381. [Google Scholar] [CrossRef]
- Lv, S.H.; Feng, C.S.; Gao, J.I.; Lu, X.S. Study on enclosing effects and biodiversity variation of desertification grassland in Hulunbeir Steppe. Acta Agrestia Sin. 2008, 16, 442–447. [Google Scholar] [CrossRef]
- Quan, Q.; Nianpeng, H.; Zhen, Z.; Yunhai, Z.; Yang, G. Nitrogen enrichment and grazing accelerate vegetation restoration in degraded grassland patches. Ecol. Eng. 2015, 75, 172–177. [Google Scholar] [CrossRef]
- Yao, M.; Rui, J.; Li, J.; Wang, J.; Cao, W.; Li, X. Soil bacterial community shifts driven by restoration time and steppe types in the degraded steppe of Inner Mongolia. Catena 2018, 165, 228–236. [Google Scholar] [CrossRef]
- Amartuvshin, N.; Kim, J.B.; Cho, N.H.; Seo, B.S.; Kang, S.K. Local and regional steppe vegetation palatability at grazing hotspot areas in Mongolia. J. Ecol. Environ. 2022, 46, 76–84. [Google Scholar] [CrossRef]
- Zhu, M.J.; Ren, A.Z.; Wen, W.; Gao, Y.B. Diversity and taxonomy of endophytes from Leymus chinensis in the Inner Mongolia steppe of China. FEMS Microbiol. Lett. 2013, 340, 135–145. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic of Site | Site * Name | Topology Type | Elevation (m) | Planted Method | Introduced or Dominant Plant (Life-Form) | Abundance State (Coverage, %) |
|---|---|---|---|---|---|---|
| Non-restoration | NR | Shifted sandy dune | 601 | - | Corethrodendron fruticosum B.H.Choi & H.Ohashi (Shrub) | 3.1 |
| Restoration | R-6yr | Sand dune | 590 | Shrub planting + Seed spray | C. fruticosum (Shrub) | 42.7 |
| R-7yr | 590 | Tree + Shrub planting | Populus canadensis Moench (Tree) C. fruticosum (Shrub) | 43.7 | ||
| R-10yr | 575 | Pinus sylvestris var. mongolica Litv. (Tree) Caragana microphylla Lam. (Shrub) | 43.9 | |||
| R-13yr | 581 | Shrub planting + Seed spray | C. fruticosum (Shrub) | 74.7 | ||
| Reference | TTS | Fixed sandy dune | 693 | - | Galium verum var. asiaticum (Nakai) T. Yamaz. (Herb) | 77.1 |
| WS | 729 | - | P. sylvestris var. mongolica (Tree) | 51.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, E.-J.; Lee, S.-H.; Kim, S.-H.; Park, J.-H.; You, Y.-H. Changes in Competitors, Stress Tolerators, and Ruderals (CSR) Ecological Strategies after the Introduction of Shrubs and Trees in Disturbed Semiarid Steppe Grasslands in Hulunbuir, Inner Mongolia. Biology 2023, 12, 1479. https://doi.org/10.3390/biology12121479
Kim E-J, Lee S-H, Kim S-H, Park J-H, You Y-H. Changes in Competitors, Stress Tolerators, and Ruderals (CSR) Ecological Strategies after the Introduction of Shrubs and Trees in Disturbed Semiarid Steppe Grasslands in Hulunbuir, Inner Mongolia. Biology. 2023; 12(12):1479. https://doi.org/10.3390/biology12121479
Chicago/Turabian StyleKim, Eui-Joo, Seung-Hyuk Lee, Se-Hee Kim, Jae-Hoon Park, and Young-Han You. 2023. "Changes in Competitors, Stress Tolerators, and Ruderals (CSR) Ecological Strategies after the Introduction of Shrubs and Trees in Disturbed Semiarid Steppe Grasslands in Hulunbuir, Inner Mongolia" Biology 12, no. 12: 1479. https://doi.org/10.3390/biology12121479
APA StyleKim, E.-J., Lee, S.-H., Kim, S.-H., Park, J.-H., & You, Y.-H. (2023). Changes in Competitors, Stress Tolerators, and Ruderals (CSR) Ecological Strategies after the Introduction of Shrubs and Trees in Disturbed Semiarid Steppe Grasslands in Hulunbuir, Inner Mongolia. Biology, 12(12), 1479. https://doi.org/10.3390/biology12121479