
PGPR-Soil Microbial Communities’ Interactions and Their Influence on Wheat Growth Promotion and Resistance Induction against Mycosphaerella graminicola


 ,
,  , , , ,
, , , ,  and
and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Microorganisms and Inoculum Preparation
2.2. Soil Physicochemical Analysis
2.3. Plant Material and Growth Conditions
2.4. Wheat Resistance Induction against M. graminicola and Wheat Biomasses
2.5. Taxonomic Analysis of Microbial Communities
2.6. Bioinformatic Analysis
2.7. Soil Microbial Metabolic Profiles
2.8. Data Analysis and Statistics
3. Results
3.1. Effects of PGPR Mixture on Wheat Biomass under Field Soil
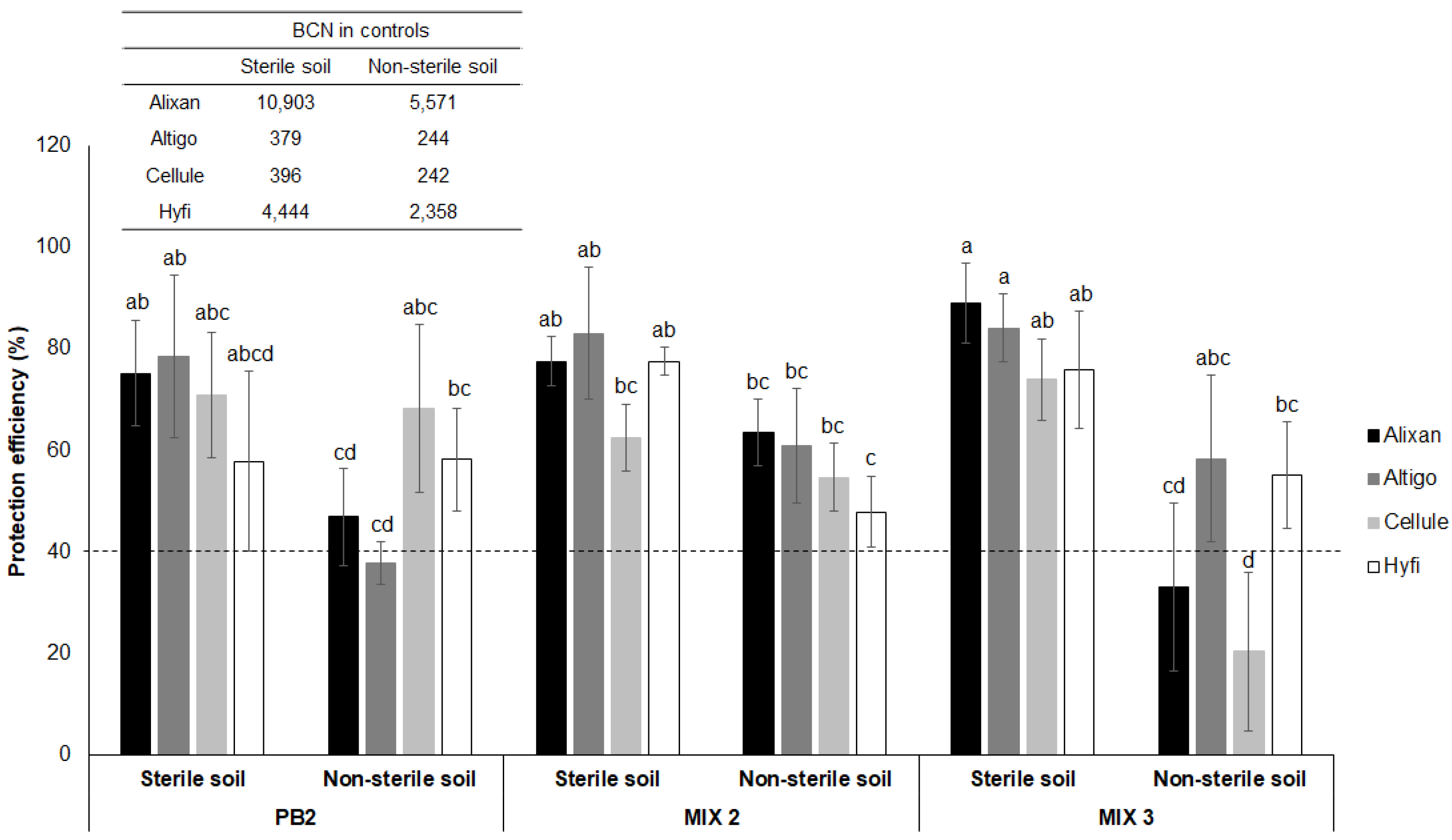
3.2. Resistance Induction in Wheat against M. graminicola
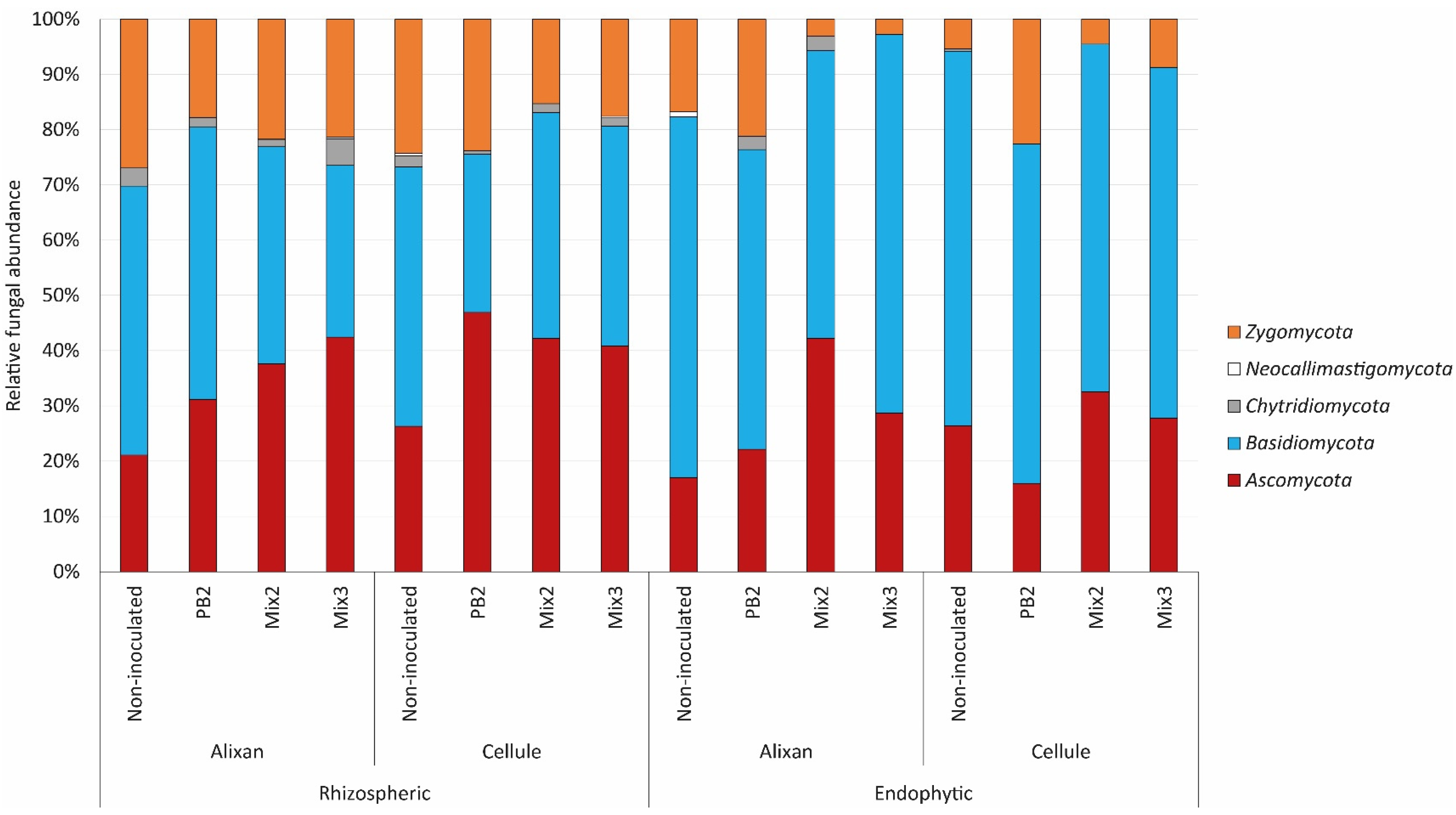
3.3. Influence of PGPR Inoculation on Microbial Communities
3.3.1. α-Diversity
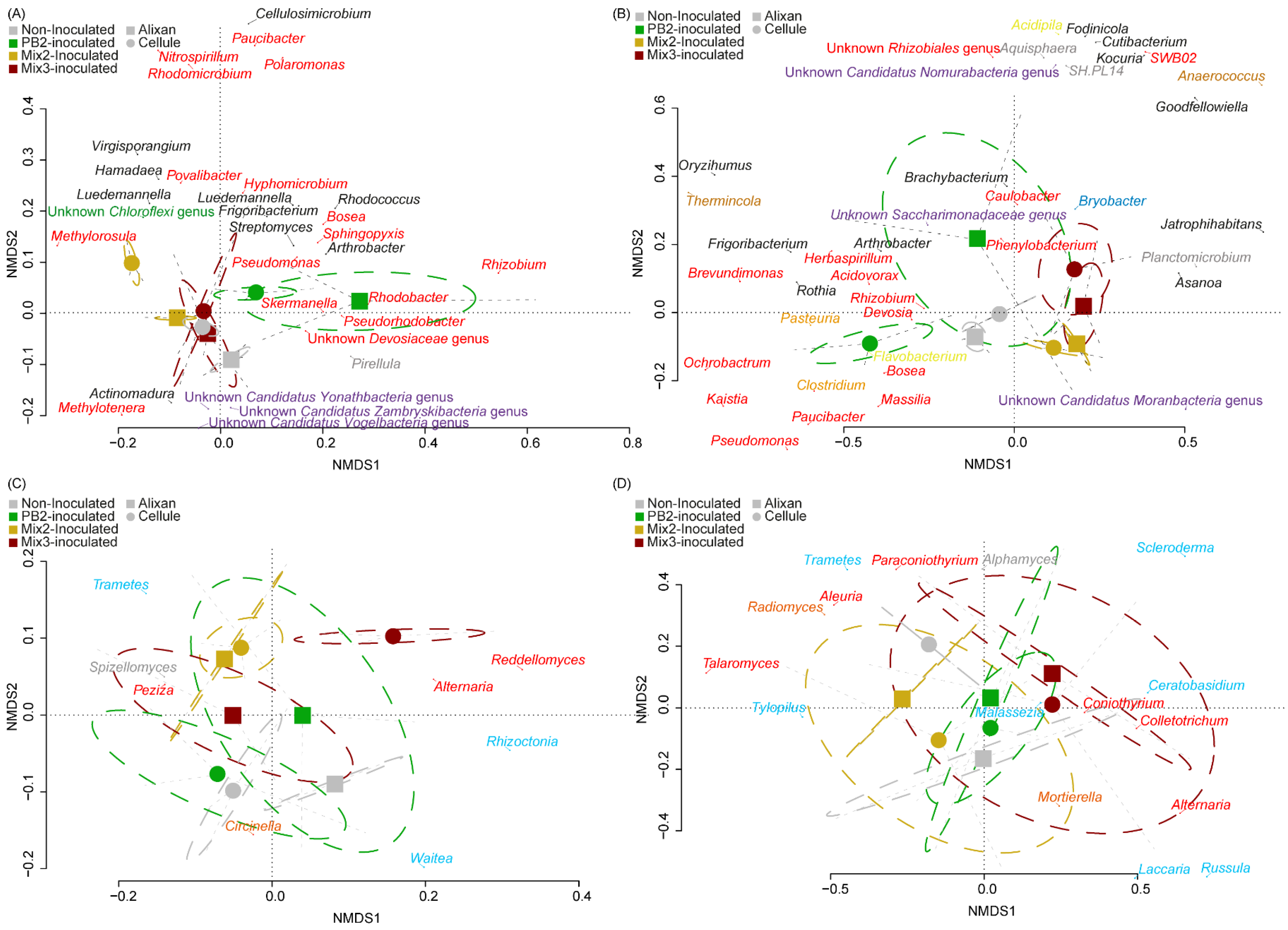
3.3.2. β-Diversity
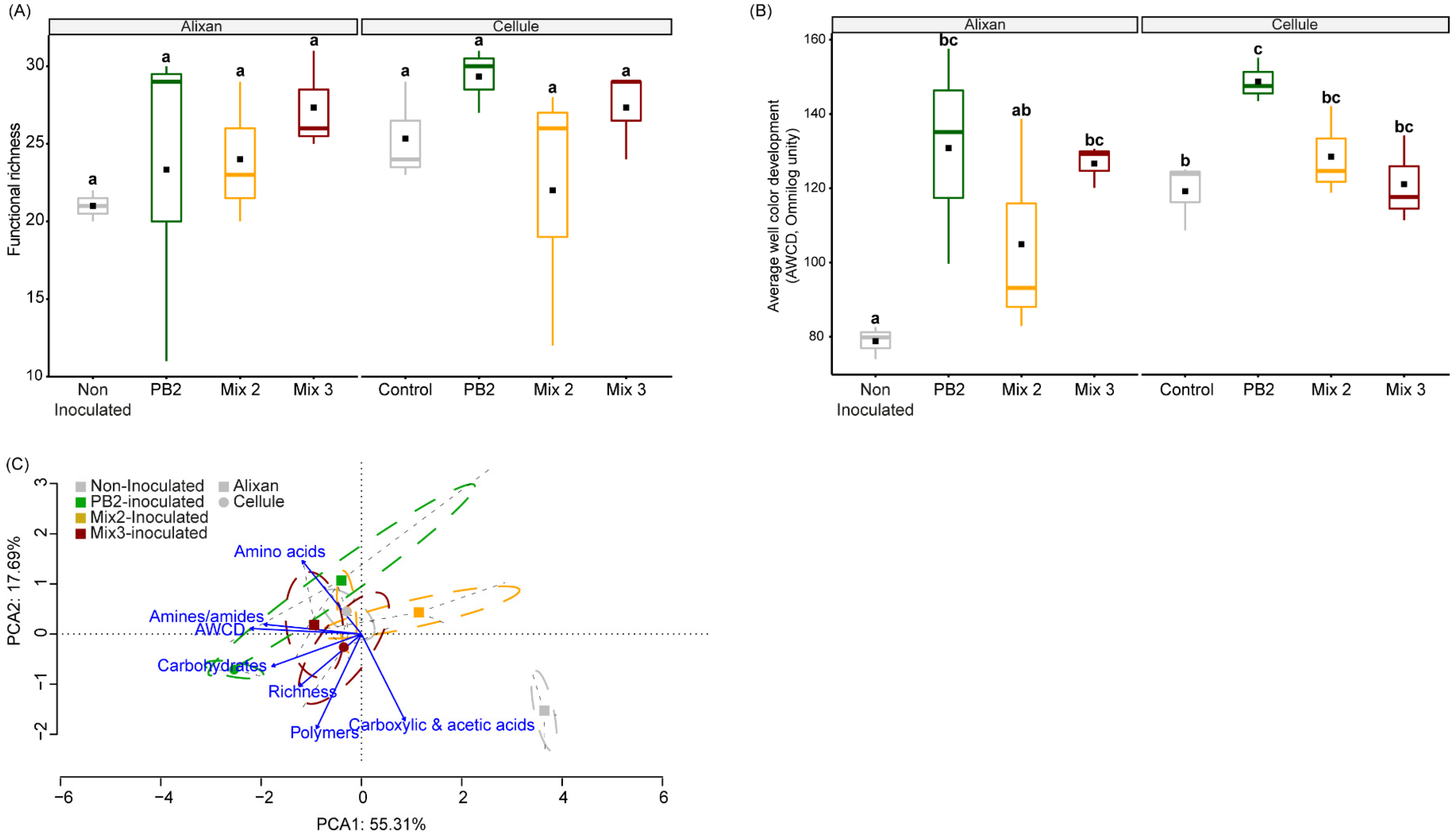
3.3.3. Microbial Community Function
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nihorimbere, V.; Ongena, M.; Smargiassi, M.; Thonart, P. Beneficial Effect of the Rhizosphere Microbial Community for Plant Growth and Health. Biotechnol. Agron. Soc. Environ. 2011, 15, 327–337. [Google Scholar]
- Selim, S.; Roisin-Fichter, C.; Andry, J.-B.; Bogdanow, B.; Sambou, R. Real-Time PCR to Study the Effect of Timing and Persistence of Fungicide Application and Wheat Varietal Resistance on Mycosphaerella graminicola and Its Sterol 14α-Demethylation-Inhibitor-Resistant Genotypes. Pest Manag. Sci. 2014, 70, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Selim, S.; Jean-Baptiste, A.; Boris, B.; Céline, R.-F. Accuracy of Real-Time PCR to Study Mycosphaerella Graminicola Epidemic in Wheat: From Spore Arrival to Fungicide Efficiency. In Fungicides—Beneficial and Harmful Aspects; INTECH Open Access Publisher: London, UK, 2011; Volume 12, pp. 219–238. [Google Scholar]
- Benhamou, N. Elicitor-Induced Plant Defence Pathways. Trends Plant Sci. 1996, 1, 233–239. [Google Scholar] [CrossRef]
- Samain, E.; van Tuinen, D.; Jeandet, P.; Aussenac, T.; Selim, S. Biological Control of Septoria Leaf Blotch and Growth Promotion in Wheat by Paenibacillus Sp. Strain B2 and Curtobacterium Plantarum Strain EDS. Biol. Control 2017, 114, 87–96. [Google Scholar] [CrossRef]
- Samain, E.; Aussenac, T.; Selim, S. The Effect of Plant Genotype, Growth Stage, and Mycosphaerella graminicola Strains on the Efficiency and Durability of Wheat-Induced Resistance by Paenibacillus Sp. Strain B2. Front. Plant Sci. 2019, 10, 587. [Google Scholar] [CrossRef]
- Walters, D.; Walsh, D.; Newton, A.; Lyon, G. Induced Resistance for Plant Disease Control: Maximizing the Efficacy of Resistance Elicitors. Phytopathology 2005, 95, 1368–1373. [Google Scholar] [CrossRef]
- Van Veen, J.A.; Overbeek, L.S.V.; Elsas, J.D.V. Fate and Activity of Microorganisms Introduced into Soil. Microbiol. Mol. Biol. Rev. 1997, 61, 15. [Google Scholar]
- Ramos, B.; García, J.A.L.; Probanza, A.; Barrientos, M.L.; Gutierrez Mañero, F.J. Alterations in the Rhizobacterial Community Associated with European Alder Growth When Inoculated with PGPR Strain Bacillus licheniformis. Environ. Exp. Bot. 2003, 49, 61–68. [Google Scholar] [CrossRef]
- Samain, E.; Ernenwein, C.; Aussenac, T.; Selim, S. Effective and Durable Systemic Wheat-Induced Resistance by a Plant-Growth-Promoting Rhizobacteria Consortium of Paenibacillus Sp. Strain B2 and Arthrobacter Spp. Strain AA against Zymoseptoria Tritici and Drought Stress. Physiol. Mol. Plant Pathol. 2022, 119, 101830. [Google Scholar] [CrossRef]
- Selim, S.; Negrel, J.; Govaerts, C.; Gianinazzi, S.; van Tuinen, D. Isolation and Partial Characterization of Antagonistic Peptides Produced by Paenibacillus sp. Strain B2 Isolated from the Sorghum Mycorrhizosphere. Appl. Environ. Microbiol. 2005, 71, 6501–6507. [Google Scholar] [CrossRef]
- Cattelan, A.J.; Hartel, P.G.; Fuhrmann, J.J. Screening for Plant Growth–Promoting Rhizobacteria to Promote Early Soybean Growth. Soil Sci. Soc. Am. J. 1999, 69, 1670–1680. [Google Scholar] [CrossRef]
- Weller, D.M. Biological Control of Soilborne Plant Pathogens in the Rhizosphere with Bacteria. Annu. Rev. Phytopathol. 1988, 26, 379–407. [Google Scholar] [CrossRef]
- Jones, M.L.; Rivett, D.W.; Pascual-García, A.; Bell, T. Relationships between Community Composition, Productivity and Invasion Resistance in Semi-Natural Bacterial Microcosms. eLife 2021, 10, e71811. [Google Scholar] [CrossRef] [PubMed]
- Selim, S.; Martin-Laurent, F.; Rouard, N.; Gianinazzi, S.; van Tuinen, D. Impact of a New Biopesticide Produced by Paenibacillus Sp. Strain B2 on the Genetic Structure and Density of Soil Bacterial Communities. Pest Manag. Sci. 2007, 63, 269–275. [Google Scholar] [CrossRef]
- Samain, E.; Ernenwein, C.; Aussenac, T.; Selim, S. Efficacity and Durability of Paenibacillus Sp. Strain B2 in Co-Inoculation with Arthrobacter Sp. SSM-004 and Microbacterium Sp. SSM-001 for Growth Promoting and Resistance Induction in Wheat against Mycosphaerella Graminicola and Drought Stress. Plant Pathol. Microbiol. 2022, 13, 1000603. [Google Scholar]
- Budi, S.W.; Tuinen, D.V.; Martinotti, G.; Gianinazzi, S. Isolation from the Sorghum Bicolor Mycorrhizosphere of a Bacterium Compatible with Arbuscular Mycorrhiza Development and Antagonistic towards Soilborne Fungal Pathogens. Appl. Environ. Microbiol. 1999, 65, 5148–5150. [Google Scholar] [CrossRef]
- Arvalis Institut du Végétal Blé Tendre—Maladies; Comportement Vis-à-Vis Des Maladies. Choisir 2017. Available online: https://www.arvalis.fr/sites/default/files/imported_files/premiers_resultats_bth_alsace_1263678181362291022.pdf (accessed on 3 November 2023).
- Bearchell, S.J.; Fraaije, B.A.; Shaw, M.W.; Fitt, B.D. Wheat Archive Links Long-Term Fungal Pathogen Population Dynamics to Air Pollution. Proc. Natl. Acad. Sci. USA 2005, 102, 5438–5442. [Google Scholar] [CrossRef]
- Alahmad, A.; Decocq, G.; Spicher, F.; Kheirbeik, L.; Kobaissi, A.; Tetu, T.; Dubois, F.; Duclercq, J. Cover Crops in Arable Lands Increase Functional Complementarity and Redundancy of Bacterial Communities. J. Appl. Ecol. 2019, 56, 651–664. [Google Scholar] [CrossRef]
- Herlemann, D.P.; Labrenz, M.; Jürgens, K.; Bertilsson, S.; Waniek, J.J.; Andersson, A.F. Transitions in Bacterial Communities along the 2000 Km Salinity Gradient of the Baltic Sea. ISME J. 2011, 5, 1571–1579. [Google Scholar] [CrossRef]
- Toju, H.; Tanabe, A.S.; Yamamoto, S.; Sato, H. High-Coverage ITS Primers for the DNA-Based Identification of Ascomycetes and Basidiomycetes in Environmental Samples. PLoS ONE 2012, 7, e40863. [Google Scholar] [CrossRef] [PubMed]
- Escudié, F.; Auer, L.; Bernard, M.; Mariadassou, M.; Cauquil, L.; Vidal, K.; Maman, S.; Hernandez-Raquet, G.; Combes, S.; Pascal, G. FROGS: Find, Rapidly, OTUs with Galaxy Solution. Bioinformatics 2018, 34, 1287–1294. [Google Scholar] [CrossRef] [PubMed]
- Martin, M. Cutadapt Removes Adapter Sequences from High-Throughput Sequencing Reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Mahé, F.; Rognes, T.; Quince, C.; de Vargas, C.; Dunthorn, M. Swarm: Robust and Fast Clustering Method for Amplicon-Based Studies. PeerJ 2014, 2, e593. [Google Scholar] [CrossRef]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A Versatile Open Source Tool for Metagenomics. PeerJ 2016, 18, e2584. [Google Scholar] [CrossRef]
- Deshpande, V.; Wang, Q.; Greenfield, P.; Charleston, M.; Porras-Alfaro, A.; Kuste, C.R.; Cole, J.R.; Midgley, D.; Tran-Dinh, N. Fungal Identification Using a Bayesian Classifier and the Warcup Training Set of Internal Transcribed Spacer Sequences. Mycologia 2015, 108, 1–5. [Google Scholar] [CrossRef]
- Langrand, J.; Lounès-Hadj Sahraou, A.; Duclercq, J.; Raveau, R.; Laruelle, F.; Bert, V.; Fontaine, J. Coriander (Coriandrum sativum) Cultivation Combined with Arbuscular Mycorrhizal Fungi Inoculation and Steel Slag Application Influences Trace Elements-Polluted Soil Bacterial Functioning. Plants 2023, 12, 618. [Google Scholar] [CrossRef]
- Weber, K.P.; Grove, J.A.; Gehder, M.; Anderson, W.A.; Legge, R.L. Data Transformations in the Analysis of Community-Level Substrate Utilization Data from Microplates. J. Microbiol. Methods 2007, 69, 461–469. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R.B. Community Ecology Package. R Pachage Version 2013, 2, 1–295. [Google Scholar]
- Mendiburu, F. De Agricolae: Statistical Procedures for Agricultural Research. 2021. Available online: https://rdrr.io/cran/agricolae/ (accessed on 13 September 2022).
- Chanway, C.P.; Turkington, R.; Holl, F.B. Ecological Implications of Specificity between Plants and Rhizosphere Micro-Organisms. Adv. Ecol. Res. 1991, 21, 121–169. [Google Scholar] [CrossRef]
- Hastings, A.; Harrison, S. Metapopulation Dynamics and Genetics. Ann. Rev. Ecol. Syst. 1994, 25, 167–188. [Google Scholar] [CrossRef]
- Levins, R. Extinction. Lect. Math. Life Sci. 1976, 2, 75–107. [Google Scholar]
- Sturz, A.V.; Novak, J. Endophytic communities of Rhizobacteria and the Strategies Requiredto Creare Yield Enhancing Associations with Crops. Appl. Soil Ecol. 2000, 15, 183–190. [Google Scholar] [CrossRef]
- Trabelsi, D.; Mhamdi, R. Microbial Inoculants and Their Impact on Soil Microbial Communities: A Review. BioMed Res. Int. 2013, 2013, 863240. [Google Scholar] [CrossRef] [PubMed]
- Ciccillo, F.; Fiore, A.; Bevivino, A.; Dalmastri, C.; Tabacchioni, S.; Chiarini, L. Effects of Two Different Application Methods of Burkholderia Ambifaria MCI 7 on Plant Growth and Rhizospheric Bacterial Diversity. Environ. Microbiol. 2002, 4, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Schreiter, S.; Ding, G.-C.; Heuer, H.; Neumann, G.; Sandmann, M.; Grosch, R.; Kropf, S.; Smalla, K. Effect of the Soil Type on the Microbiome in the Rhizosphere of Field-Grown Lettuce. Front. Microbiol. 2014, 5, 144. [Google Scholar] [CrossRef]
- Gholami, A.; Shahsavani, S.; Nezarat, S. The Effect of Plant Growth Promoting Rhizobacteria (PGPR) on Germination, Seedling Growth and Yield of Maize. Int. J. Biol. Life Sci. 2009, 1, 35–40. [Google Scholar]
- Backer, R.; Rokem, J.S.; Ilangumaran, G.; Lamont, J.; Praslickova, D.; Ricci, E.; Subramanian, S.; Smith, D.L. Plant Growth-Promoting Rhizobacteria: Context, Mechanisms of Action, and Roadmap to Commercialization of Biostimulants for Sustainable Agriculture. Front. Plant Sci. 2018, 9, 1473. [Google Scholar] [CrossRef]
- Nelson, L.M. Plant Growth Promoting Rhizobacteria (PGPR): Prospects for New Inoculants. Crop Manag. 2004, 3, 301–305. [Google Scholar] [CrossRef]
- Sturz, A.V.; Christie, B.R. Endophytic Bacteria of Red Clover as Agents of Allelopathic Clover-Maize Syndromes. Soil Biol. Biochem. 1996, 28, 583–588. [Google Scholar] [CrossRef]
- Sturz, A.V.; Christie, B.R. Endophytic Bacterial Systems Governing Red Clover Growth and Development. Ann. Appl. Biol. 1995, 126, 285–290. [Google Scholar] [CrossRef]
- Gagliardi, J.V.; Buyer, J.S.; Angle, J.S.; Russek-Cohen, E. Structural and Functional Analysis of Whole-Soil Microbial Communities for Risk and Efficacy Testing Following Microbial Inoculation of Wheat Roots in Diverse Soils. Soil Biol. 2001, 33, 25–40. [Google Scholar] [CrossRef]
- Bashan, Y.; Kloepper, J.W.; De-Bashan, L.E.; Nannipieri, P. A Need for Disclosure of the Identity of Microorganisms, Constituents, and Application Methods When Reporting Tests with Microbe-Based or Pesticide-Based Products. Biol. Fertil. Soils 2016, 52, 283–284. [Google Scholar] [CrossRef]
- Mawarda, P.C.; Le Roux, X.; Van Elsas, J.D.; Salles, J.F. Deliberate Introduction of Invisible Invaders: A Critical Appraisal of the Impact of Microbial Inoculants on Soil Microbial Communities. Soil Biol. Biochem. 2020, 148, 107874. [Google Scholar] [CrossRef]
- Mitter, E.K.; Tosi, M.; Obregón, D.; Dunfield, K.E.; Germida, J.J. Rethinking Crop Nutrition in Times of Modern Microbiology: Innovative Biofertilizer Technologies. Front. Sustain. Food Syst. 2021, 5, 606815. [Google Scholar] [CrossRef]
- Ardanov, P.; Sessitsch, A.; Häggman, H.; Kozyrovska, N.; Pirttilä, A.M. Methylobacterium-Induced Endophyte Community Changes Correspond with Protection of Plants against Pathogen Attack. PLoS ONE 2012, 7, e46802. [Google Scholar] [CrossRef]
- Karimi, B.; Maron, P.A.; Chemidlin-Prevost Boure, N.; Bernard, N.; Gilbert, D.; Ranjard, L. Microbial Diversity and Ecological Networks as Indicators of Environmental Quality. Environ. Chem. Lett. 2017, 15, 265–281. [Google Scholar] [CrossRef]
- Ajilogba, C.F.; Olanrewaju, O.S.; Babalola, O.O. Plant Growth Stage Drives the Temporal and Spatial Dynamics of the Bacterial Microbiome in the Rhizosphere of Vigna Subterranea. Front. Microbiol. 2022, 13, 825377. [Google Scholar] [CrossRef]
- Çakmakçı, R.; Dönmez, F.; Aydın, A.; Şahin, F. Growth Promotion of Plants by Plant Growth-Promoting Rhizobacteria under Greenhouse and Two Different Field Soil Conditions. Soil Biol. Biochem. 2006, 38, 1482–1487. [Google Scholar] [CrossRef]
- Liu, X.; Jiang, X.; He, X.; Zhao, W.; Cao, Y.; Guo, T.; Li, T.; Ni, H.; Tang, X. Phosphate-Solubilizing Pseudomonas Sp. Strain P34-L Promotes Wheat Growth by Colonizing the Wheat Rhizosphere and Improving the Wheat Root System and Soil Phosphorus Nutritional Status. J. Plant Growth Regul. 2019, 38, 1314–1324. [Google Scholar] [CrossRef]
- Yanni, Y.G.; Dazzo, F.B.; Squartini, A.; Zanardo, M.; Zidan, M.I.; Elsadany, A.E.Y. Assessment of the Natural Endophytic Association between Rhizobium and Wheat and Its Ability to Increase Wheat Production in the Nile Delta. Plant Soil 2016, 407, 367–383. [Google Scholar] [CrossRef]
- Podolich, O.V.; Ardanov, P.; Zaets, I.; Pirttilä, A.M.; Kozyrovska, N. Reviving of the Endophytic Bacterial Community as a Putative Mechanism of Plant Resistance. Plant Soil 2015, 388, 367–377. [Google Scholar] [CrossRef]
- Podolich, O.V.; Ardanov, P.; Voznyuk, T.M.; Kovalchuk, M.V.; Danylchenko, O.V.; Laschevskyi, V.V.; Lyastchenko, S.A.; Kozyrovska, N.O. Endophytic Bacteria from Potato in Vitro Activated by Exogenic Non-Pathogenic Bacteria. Biopolym. Cell 2007, 23, 21–27. [Google Scholar] [CrossRef][Green Version]
- Yin, C.; Schlatter, D.C.; Kroese, D.R.; Paulitz, T.C.; Hagerty, C.H. Responses of Soil Fungal Communities to Lime Application in Wheat Fields in the Pacific Northwest. Front. Microbiol. 2021, 12, 576763. [Google Scholar] [CrossRef]
- Ou, T.; Zhang, M.; Gao, H.; Wang, F.; Xu, W.; Liu, X.; Wang, L.; Wang, R.; Xie, J. Study on the Potential for Stimulating Mulberry Growth and Drought Tolerance of Plant Growth-Promoting Fungi. Int. J. Mol. Sci. 2023, 24, 4090. [Google Scholar] [CrossRef] [PubMed]
- Man, Y.; Wang, J.; Tam, N.F.Y.; Wan, X.; Huang, W.; Zheng, Y.; Yang, Y. Responses of Rhizosphere and Bulk Substrate Microbiome to Wastewater-Borne Sulfonamides in Constructed Wetlands with Different Plant Species. Sci. Total Environ. 2020, 706, 135955. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Kour, D.; Yadav, A.N.; Saxena, R.; Rai, P.K.; Jyoti, A.; Tomar, R.S. Biodiversity of Methylotrophic Microbial Communities and Their Potential Role in Mitigation of Abiotic Stresses in Plants. Biologia 2019, 74, 287–308. [Google Scholar] [CrossRef]
- Turzhanova, A.; Khapilina, O.N.; Tumenbayeva, A.; Shevtsov, V.; Raiser, O.; Kalendar, R. Genetic Diversity of Alternaria Species Associated with Black Point in Wheat Grains. PeerJ 2020, 8, e9097. [Google Scholar] [CrossRef]
- Yin, C.; Hulbert, S.H.; Schroeder, K.L.; Mavrodi, O.; Mavrodi, D.; Dhingra, A.; Schillinger, W.F.; Paulitz, T.C. Role of Bacterial Communities in the Natural Suppression of Rhizoctonia Solani Bare Patch Disease of Wheat (Triticum aestivum L.). Appl. Environ. Microbiol. 2013, 79, 7428–7438. [Google Scholar] [CrossRef]
- Jones, E.E.; Rabeendran, N.; Stewart, A. Biocontrol of Sclerotinia sclerotiorum Infection of Cabbage by Coniothyrium minitans and Trichoderma spp. Biocontrol Sci. Technol. 2014, 24, 1363–1382. [Google Scholar] [CrossRef]
- Ojaghian, M.R. Biocontrol of Potato White Mold Using Coniothyrium minitans and Resistance of Potato Cultivars to Sclerotinia sclerotiorum. Plant Pathol. J. 2010, 26, 346–352. [Google Scholar] [CrossRef]
- Sun, Q.; Zhang, P.; Zhao, Z.; Li, X.; Sun, X.; Jiang, W. Continuous Wheat/Soybean Cropping Influences Soybean Yield and Rhizosphere Microbial Community Structure and Function. Agronomy 2022, 13, 28. [Google Scholar] [CrossRef]
- Kozdrój, J.; Trevors, J.T.; van Elsas, J.D. Influence of Introduced Potential Biocontrol Agents on Maize Seedling Growth and Bacterial Community Structure in the Rhizosphere. Soil Biol. Biochem. 2004, 36, 1775–1784. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Analysis Target | Cultivar | Modality | Richness | Shannon | ||||
|---|---|---|---|---|---|---|---|---|
| Mean | SD | Sign | Mean | SD | Sign | |||
| Rhizosphere | Alixan | Non-inoculated | 2454.7 | 222.23 | b | 6.435 | 0.01 | f |
| PB2 | 1776.7 | 442.10 | c | 6.134 | 0.27 | bcf | ||
| Mix2 | 2664.3 | 293.14 | b | 6.298 | 0.16 | f | ||
| Mix3 | 2445.0 | 846.29 | b | 6.273 | 0.14 | cf | ||
| Cellule | Non-inoculated | 2904.3 | 399.74 | b | 6.316 | 0.01 | f | |
| PB2 | 2735.7 | 225.90 | b | 6.330 | 0.11 | f | ||
| Mix2 | 2793.0 | 146.43 | b | 6.271 | 0.12 | cf | ||
| Mix3 | 2583.3 | 605.81 | b | 6.258 | 0.14 | cf | ||
| Endosphere | Alixan | Non-inoculated | 848.0 | 173.35 | a | 5.626 | 0.08 | ad |
| PB2 | 606.7 | 524.89 | a | 5.242 | 0.52 | de | ||
| Mix2 | 1069.3 | 178.21 | a | 5.922 | 0.08 | abc | ||
| Mix3 | 820.7 | 350.44 | a | 5.820 | 0.27 | ab | ||
| Cellule | Non-inoculated | 818.5 | 95.46 | a | 5.681 | 0.07 | a | |
| PB2 | 533.0 | 137.96 | a | 5.172 | 0.31 | e | ||
| Mix2 | 977.0 | 327.11 | a | 5.828 | 0.21 | ab | ||
| Mix3 | 670.3 | 422.44 | a | 5.649 | 0.35 | a | ||
| Analysis Target | Cultivar | Modality | Richness | Shannon | ||||
|---|---|---|---|---|---|---|---|---|
| Mean | SD | Sign | Mean | SD | Sign | |||
| Rhizosphere | Alixan | Non-inoculated | 270.67 | 13.32 | c | 4.34 | 0.18 | d |
| PB2 | 241.00 | 18.52 | bc | 3.22 | 1.29 | bf | ||
| Mix2 | 217.67 | 13.05 | b | 4.16 | 0.34 | de | ||
| Mix3 | 223.67 | 23.12 | b | 3.80 | 0.63 | def | ||
| Cellule | Non-inoculated | 246.67 | 33.08 | bc | 4.37 | 0.06 | d | |
| PB2 | 239.00 | 7.81 | b | 3.30 | 1.15 | bef | ||
| Mix2 | 233.00 | 8.89 | b | 4.16 | 0.15 | de | ||
| Mix3 | 237.67 | 46.69 | b | 4.12 | 0.29 | def | ||
| Endosphere | Alixan | Non-inoculated | 26.67 | 9.45 | a | 2.15 | 0.03 | ac |
| PB2 | 25.33 | 15.28 | a | 2.15 | 0.35 | ac | ||
| Mix2 | 28.33 | 6.35 | a | 2.74 | 0.14 | ab | ||
| Mix3 | 17.33 | 6.35 | a | 2.11 | 0.70 | ac | ||
| Cellule | Non-inoculated | 23.00 | 8.49 | a | 2.55 | 0.42 | abc | |
| PB2 | 23.67 | 2.31 | a | 1.77 | 0.41 | c | ||
| Mix2 | 28.00 | 8.89 | a | 2.61 | 0.29 | abc | ||
| Mix3 | 18.00 | 11.27 | a | 2.01 | 0.46 | ac | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Samain, E.; Duclercq, J.; Ait Barka, E.; Eickermann, M.; Ernenwein, C.; Mazoyon, C.; Sarazin, V.; Dubois, F.; Aussenac, T.; Selim, S. PGPR-Soil Microbial Communities’ Interactions and Their Influence on Wheat Growth Promotion and Resistance Induction against Mycosphaerella graminicola. Biology 2023, 12, 1416. https://doi.org/10.3390/biology12111416
Samain E, Duclercq J, Ait Barka E, Eickermann M, Ernenwein C, Mazoyon C, Sarazin V, Dubois F, Aussenac T, Selim S. PGPR-Soil Microbial Communities’ Interactions and Their Influence on Wheat Growth Promotion and Resistance Induction against Mycosphaerella graminicola. Biology. 2023; 12(11):1416. https://doi.org/10.3390/biology12111416
Chicago/Turabian StyleSamain, Erika, Jérôme Duclercq, Essaïd Ait Barka, Michael Eickermann, Cédric Ernenwein, Candice Mazoyon, Vivien Sarazin, Frédéric Dubois, Thierry Aussenac, and Sameh Selim. 2023. "PGPR-Soil Microbial Communities’ Interactions and Their Influence on Wheat Growth Promotion and Resistance Induction against Mycosphaerella graminicola" Biology 12, no. 11: 1416. https://doi.org/10.3390/biology12111416
APA StyleSamain, E., Duclercq, J., Ait Barka, E., Eickermann, M., Ernenwein, C., Mazoyon, C., Sarazin, V., Dubois, F., Aussenac, T., & Selim, S. (2023). PGPR-Soil Microbial Communities’ Interactions and Their Influence on Wheat Growth Promotion and Resistance Induction against Mycosphaerella graminicola. Biology, 12(11), 1416. https://doi.org/10.3390/biology12111416