

Lactose Permease Scrambles Phospholipids


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Protein Expression and Purification of LacY and Its Mutants
2.3. Reconstitution of LacY into Liposomes
2.4. PI-PLC Assay
2.5. Scrambling of NBD-Lipids
3. Results and Discussion
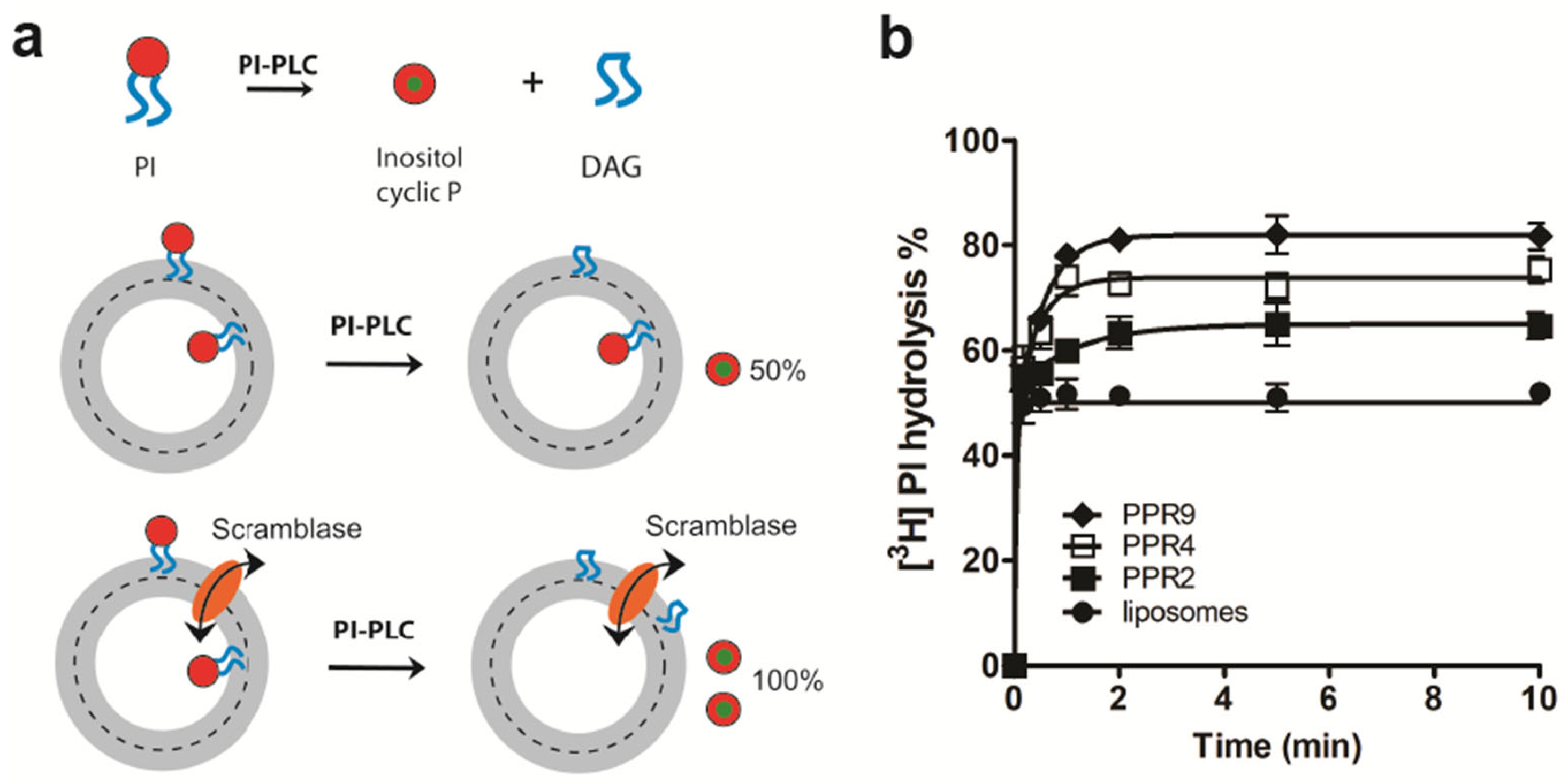
3.1. LacY Is a Phospholipid Scramblase
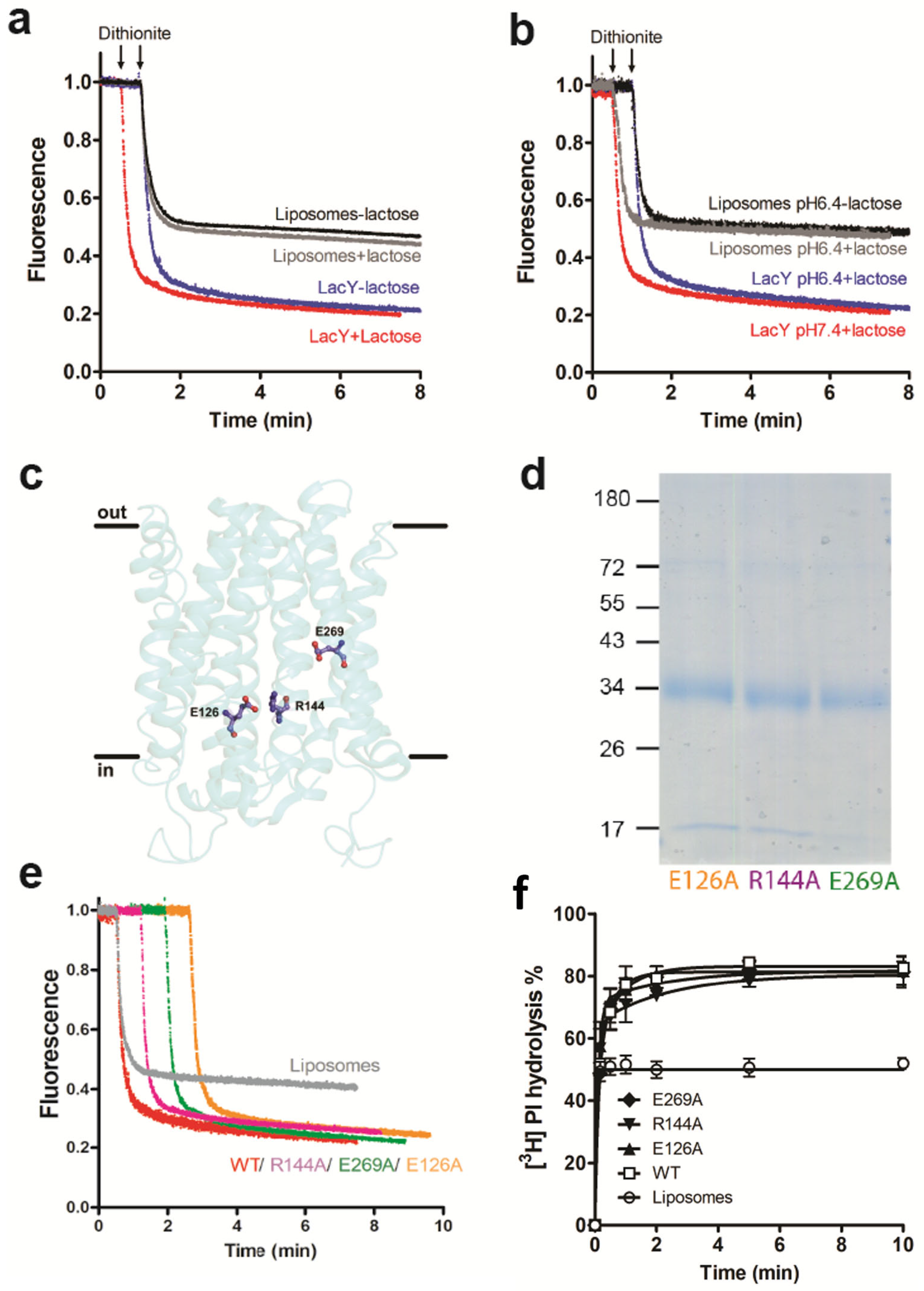
3.2. Glycerophospholipid Scrambling by LacY Is Independent of H+/Lactose Transport
3.3. Occluded Conformation of LacY Blocks Glycerophospholipid Scrambling
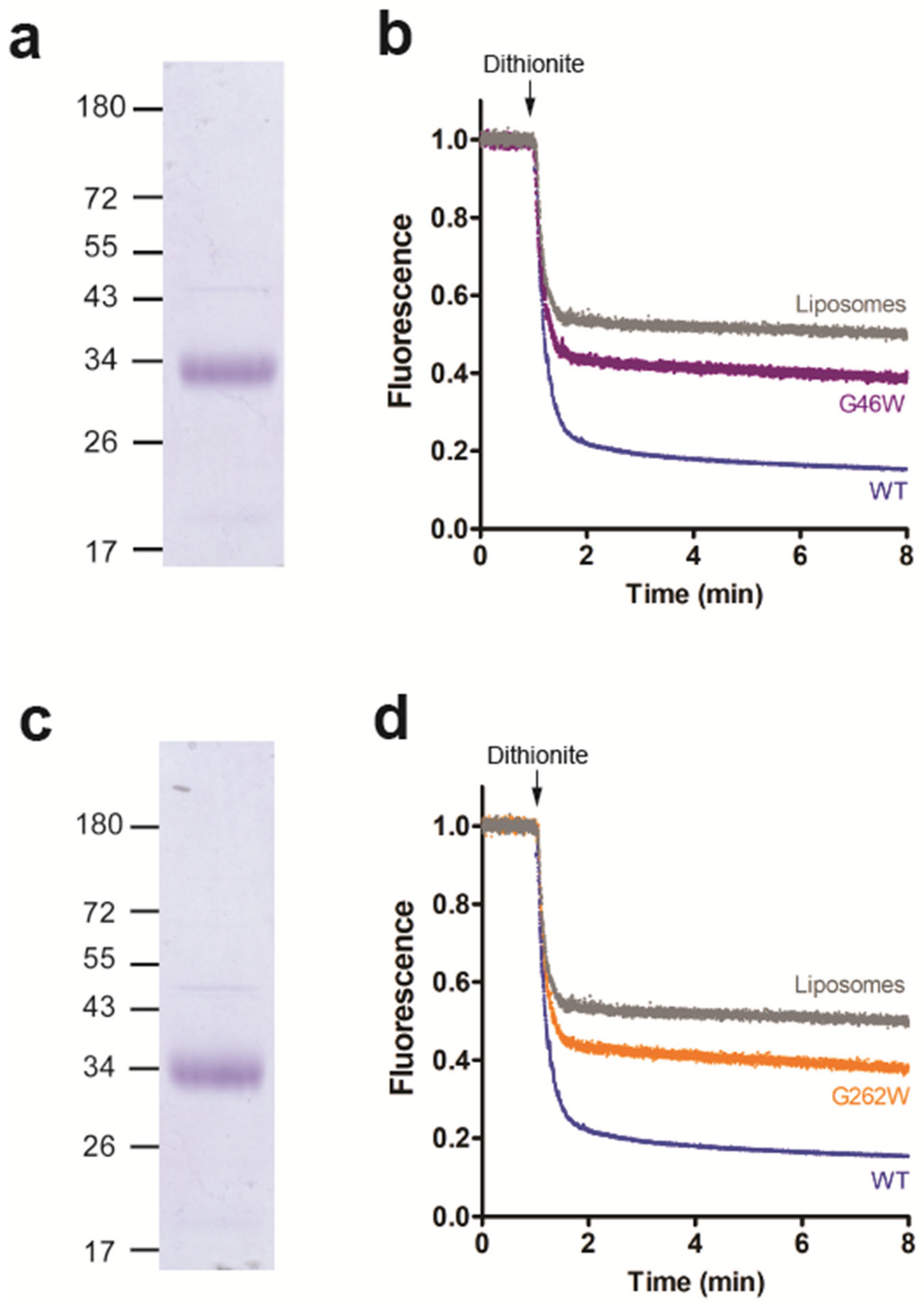
3.4. Amino Acid Residues G46 and G262 Are Important for LacY Scramblase Activity
4. Conclusions and Perspectives
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ballas, L.M.; Bell, R.M. Topography of Phosphatidylcholine, Phosphatidylethanolamine and Triacylglycerol Biosynthetic Enzymes in Rat Liver Microsomes. Biochim. Biophys. Acta 1980, 602, 578–590. [Google Scholar] [CrossRef] [PubMed]
- Kuchler, K.; Daum, G.; Paltauf, F. Subcellular and Submitochondrial Localization of Phospholipid-Synthesizing Enzymes in Saccharomyces cerevisiae. J. Bacteriol. 1986, 165, 901–910. [Google Scholar] [CrossRef] [PubMed]
- Henneberry, A.L.; Wright, M.M.; McMaster, C.R. The major sites of cellular phospholipid synthesis and molecular determinants of fatty acid and lipid head group specificity. Mol. Biol. Cell 2002, 13, 3148–3161. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, N.; Farine, L.; Pandey, K.; Menon, A.K.; Butikofer, P. Lipid topogenesis—35 years on. BBA-Mol. Cell Biol. Lipids 2016, 1861, 757–766. [Google Scholar] [CrossRef] [PubMed]
- Pomorski, T.; Menon, A.K. Lipid flippases and their biological functions. Cell. Mol. Life Sci. 2006, 63, 2908–2921. [Google Scholar] [CrossRef] [PubMed]
- Sanyal, S.; Menon, A.K. Flipping Lipids: Why an’ What’s the Reason for? ACS Chem. Biol. 2009, 4, 895–909. [Google Scholar] [CrossRef] [PubMed]
- Montigny, C.; Lyons, J.; Champeil, P.; Nissen, P.; Lenoir, G. On the molecular mechanism of flippase- and scramblase-mediated phospholipid transport. BBA-Mol. Cell Biol. Lipids 2016, 1861, 767–783. [Google Scholar] [CrossRef]
- Sebastian, T.T.; Baldridge, R.D.; Xu, P.; Graham, T.R. Phospholipid flippases: Building asymmetric membranes and transport vesicles. BBA-Mol. Cell Biol. Lipids 2012, 1821, 1068–1077. [Google Scholar] [CrossRef]
- Pomorski, T.G.; Menon, A.K. Lipid somersaults: Uncovering the mechanisms of protein-mediated lipid flipping. Prog. Lipid Res. 2016, 64, 69–84. [Google Scholar] [CrossRef]
- Bishop, W.R.; Bell, R.M. Assembly of the Endoplasmic Reticulum Phospholipid Bilayer: The Phosphatidylcholine Transporter. Cell 1985, 42, 51–60. [Google Scholar] [CrossRef]
- Backer, J.M.; Dawidowicz, E.A. Reconstitution of a Phospholipid Flippase from Rat Liver Microsomes. Nature 1987, 327, 341–343. [Google Scholar] [CrossRef] [PubMed]
- Menon, A.K.; Watkins, W.E.; Hrafnsdottir, S. Specific proteins are required to translocate phosphatidylcholine bidirectionally across the endoplasmic reticulum. Curr. Biol. 2000, 10, 241–252. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, J.; Fujii, T.; Imao, T.; Ishihara, K.; Kuba, H.; Nagata, S. Calcium-dependent Phospholipid Scramblase Activity of TMEM16 Protein Family Members. J. Biol. Chem. 2013, 288, 13305–13316. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, J.; Umeda, M.; Sims, P.J.; Nagata, S. Calcium-dependent phospholipid scrambling by TMEM16F. Nature 2010, 468, 834–838. [Google Scholar] [CrossRef]
- Wang, Y.C.; Menon, A.K.; Maki, Y.; Liu, Y.S.; Iwasaki, Y.; Fujita, M.; Guerrero, P.A.; Silva, D.V.; Seeberger, P.H.; Murakami, Y.; et al. Genome-wide CRISPR screen reveals CLPTM1L as a lipid scramblase required for efficient glycosylphosphatidylinositol biosynthesis. Proc. Natl. Acad. Sci. USA 2022, 119, e2115083119. [Google Scholar] [CrossRef]
- Ernst, O.P.; Menon, A.K. Phospholipid scrambling by rhodopsin. Photochem. Photobiol. Sci. 2015, 14, 1922–1931. [Google Scholar] [CrossRef]
- Ploier, B.; Caro, L.N.; Morizumi, T.; Pandey, K.; Pearring, J.N.; Goren, M.A.; Finnemann, S.C.; Graumann, J.; Arshavsky, V.Y.; Dittman, J.S.; et al. Dimerization deficiency of enigmatic retinitis pigmentosa-linked rhodopsin mutants. Nat. Commun. 2016, 7, 12832. [Google Scholar] [CrossRef]
- Menon, I.; Huber, T.; Sanyal, S.; Banerjee, S.; Barre, P.; Canis, S.; Warren, J.D.; Hwa, J.; Sakmar, T.P.; Menon, A.K. Opsin Is a Phospholipid Flippase. Curr. Biol. 2011, 21, 149–153. [Google Scholar] [CrossRef]
- Suzuki, J.; Denning, D.P.; Imanishi, E.; Horvitz, H.R.; Nagata, S. Xk-Related Protein 8 and CED-8 Promote Phosphatidylserine Exposure in Apoptotic Cells. Science 2013, 341, 403–406. [Google Scholar] [CrossRef]
- Kaback, H.R.; Guan, L. It takes two to tango: The dance of the permease. J. Gen. Physiol. 2019, 151, 878–886. [Google Scholar] [CrossRef]
- Brenner, S. A night at the operon. Nature 1997, 386, 235. [Google Scholar] [CrossRef]
- Buchel, D.E.; Gronenborn, B.; Muller-Hill, B. Sequence of the Lactose Permease Gene. Nature 1980, 283, 541–545. [Google Scholar] [CrossRef] [PubMed]
- Newman, M.J.; Foster, D.L.; Wilson, T.H.; Kaback, H.R. Purification and Reconstitution of Functional Lactose Carrier from Escherichia coli. J. Biol. Chem. 1981, 256, 11804–11808. [Google Scholar] [CrossRef] [PubMed]
- Abramson, J.; Smirnova, I.; Kasho, V.; Verner, G.; Kaback, H.R.; Iwata, S. Structure and mechanism of the lactose permease of Escherichia coli. Science 2003, 301, 610–615. [Google Scholar] [CrossRef]
- Kaback, H.R.; Smirnova, I.; Kasho, V.; Nie, Y.L.; Zhou, Y.G. The Alternating Access Transport Mechanism in LacY. J. Membr. Biol. 2011, 239, 85–93. [Google Scholar] [CrossRef]
- Kaback, H.R. A chemiosmotic mechanism of symport. Proc. Natl. Acad. Sci. USA 2015, 112, 1259–1264. [Google Scholar] [CrossRef]
- Ilgu, H.; Jeckelmann, J.M.; Gapsys, V.; Ucurum, Z.; de Groot, B.L.; Fotiadis, D. Insights into the molecular basis for substrate binding and specificity of the wild-type L-arginine/agmatine antiporter AdiC. Proc. Natl. Acad. Sci. USA 2016, 113, 10358–10363. [Google Scholar] [CrossRef]
- Wang, L.; Iwasaki, Y.; Andra, K.K.; Pandey, K.; Menon, A.K.; Butikofer, P. Scrambling of natural and fluorescently tagged phosphatidylinositol by reconstituted G protein–coupled receptor and TMEM16 scramblases. J. Biol. Chem. 2018, 293, 18318–18327. [Google Scholar] [CrossRef]
- Ploier, B.; Menon, A.K. A Fluorescence-based Assay of Phospholipid Scramblase Activity. JoVE-J. Vis. Exp. 2016, 115, e54635. [Google Scholar]
- Chang, Q.L.; Gummadi, S.N.; Menon, A.K. Chemical modification identifies two populations of glycerophospholipid flippase in rat liver ER. Biochemistry 2004, 43, 10710–10718. [Google Scholar] [CrossRef]
- Goren, M.A.; Morizumi, T.; Menon, I.; Joseph, J.S.; Dittman, J.S.; Cherezov, V.; Stevens, R.C.; Ernst, O.P.; Menon, A.K. Constitutive phospholipid scramblase activity of a G protein-coupled receptor. Nat. Commun. 2014, 5, 5115. [Google Scholar] [CrossRef] [PubMed]
- Brunner, J.D.; Lim, N.K.; Schenck, S.; Duerst, A.; Dutzler, R. X-ray structure of a calcium-activated TMEM16 lipid scramblase. Nature 2014, 516, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.E.; Wang, Y.C.; Du, X.M.; Zhang, T.Z.; Mak, H.Y.; Hancock, S.E.; McEwen, H.; Pandzic, E.; Whan, R.M.; Aw, Y.C.; et al. TMEM41B and VMP1 are scramblases and regulate the distribution of cholesterol and phosphatidylserine. J. Cell Biol. 2021, 220, e202103105. [Google Scholar] [CrossRef] [PubMed]
- Adlakha, J.; Hong, Z.P.; Li, P.Q.; Reinisch, K.M. Structural and biochemical insights into lipid transport by VPS13 proteins. J. Cell Biol. 2022, 221, e202202030. [Google Scholar] [CrossRef]
- Butikofer, P.; Lin, Z.W.; Chiu, D.T.Y.; Lubin, B.; Kuypers, F.A. Transbilayer Distribution and Mobility of Phosphatidylinositol in Human Red Blood Cells. J. Biol. Chem. 1990, 265, 16035–16038. [Google Scholar] [CrossRef]
- Vidugiriene, J.; Menon, A.K. Early Lipid Intermediates in Glycosyl-Phosphatidylinositol Anchor Assembly Are Synthesized in the Er and Located in the Cytoplasmic Leaflet of the Er Membrane Bilayer. J. Cell Biol. 1993, 121, 987–996. [Google Scholar] [CrossRef]
- Jahn, H.; Bartoš, L.; Dearden, G.I.; Dittman, J.S.; Holthuis, J.C.M.; Vácha, R.; Menon, A.K. Phospholipids are imported into mitochondria by VDAC, a dimeric beta barrel scramblase. bioRxiv 2023. [Google Scholar] [CrossRef]
- Liu, Z.Y.; Madej, M.G.; Kaback, H.R. Helix Dynamics in LacY: Helices II and IV. J. Mol. Biol. 2010, 396, 617–626. [Google Scholar] [CrossRef]
- Kumar, H.; Kasho, V.; Smirnova, I.; Finer-Moore, J.S.; Kaback, H.R.; Stroud, R.M. Structure of sugar-bound LacY. Proc. Natl. Acad. Sci. USA 2014, 111, 1784–1788. [Google Scholar] [CrossRef]
- Khelashvili, G.; Menon, A.K. Phospholipid Scrambling by G Protein–Coupled Receptors. Annu. Rev. Biophys. 2022, 51, 39–61. [Google Scholar] [CrossRef]
- Falzone, M.E.; Feng, Z.; Alvarenga, O.E.; Pan, Y.G.; Lee, B.; Cheng, X.L.; Fortea, E.; Scheuring, S.; Accardi, A. TMEM16 scramblases thin the membrane to enable lipid scrambling. Nat. Commun. 2022, 13, 2604. [Google Scholar] [CrossRef] [PubMed]
- Bethel, N.P.; Grabe, M. Atomistic insight into lipid translocation by a TMEM16 scramblase. Proc. Natl. Acad. Sci. USA 2016, 113, 14049–14054. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Rocha-Roa, C.; Schilling, M.A.; Reinisch, K.M.; Vanni, S. Lipid scrambling is a general feature of protein insertases. bioRxiv 2023. [Google Scholar] [CrossRef]
- Bartoš, L.; Menon, A.K.; Vácha, R. Insertases Scramble Lipids: Molecular Simulations of MTCH2. bioRxiv 2023. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Bütikofer, P. Lactose Permease Scrambles Phospholipids. Biology 2023, 12, 1367. https://doi.org/10.3390/biology12111367
Wang L, Bütikofer P. Lactose Permease Scrambles Phospholipids. Biology. 2023; 12(11):1367. https://doi.org/10.3390/biology12111367
Chicago/Turabian StyleWang, Lei, and Peter Bütikofer. 2023. "Lactose Permease Scrambles Phospholipids" Biology 12, no. 11: 1367. https://doi.org/10.3390/biology12111367
APA StyleWang, L., & Bütikofer, P. (2023). Lactose Permease Scrambles Phospholipids. Biology, 12(11), 1367. https://doi.org/10.3390/biology12111367