
Photoautotrophic Growth Rate Enhancement of Synechocystis sp. PCC6803 by Heterologous Production of 2-Oxoglutarate:Ferredoxin Oxidoreductase from Chlorobaculum tepidum


Abstract
Simple Summary
Abstract
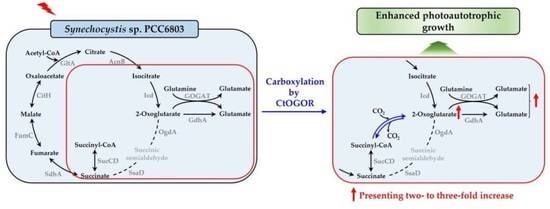
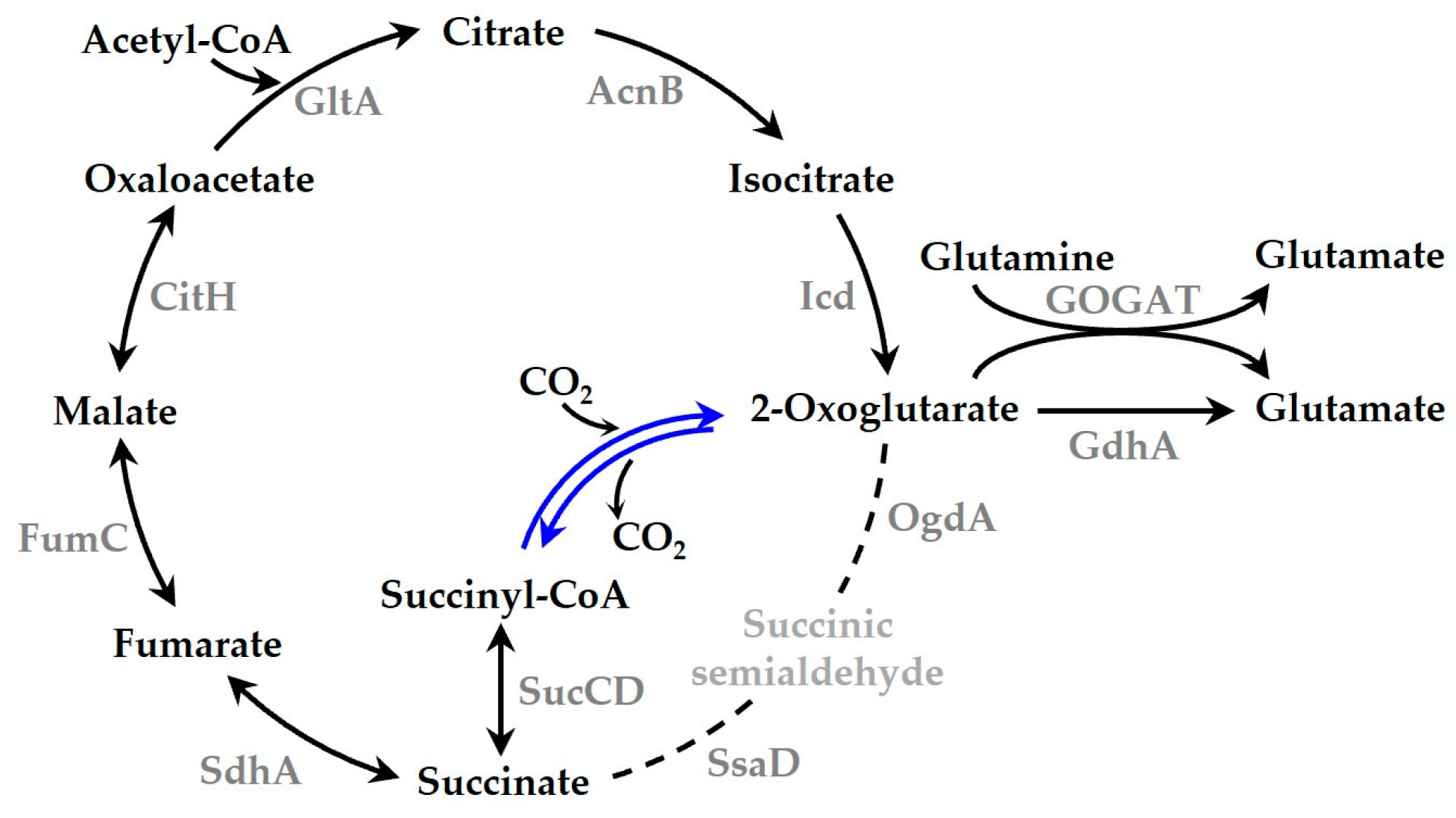
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Construction of Plasmids and Their Mobilization into Synechocystis
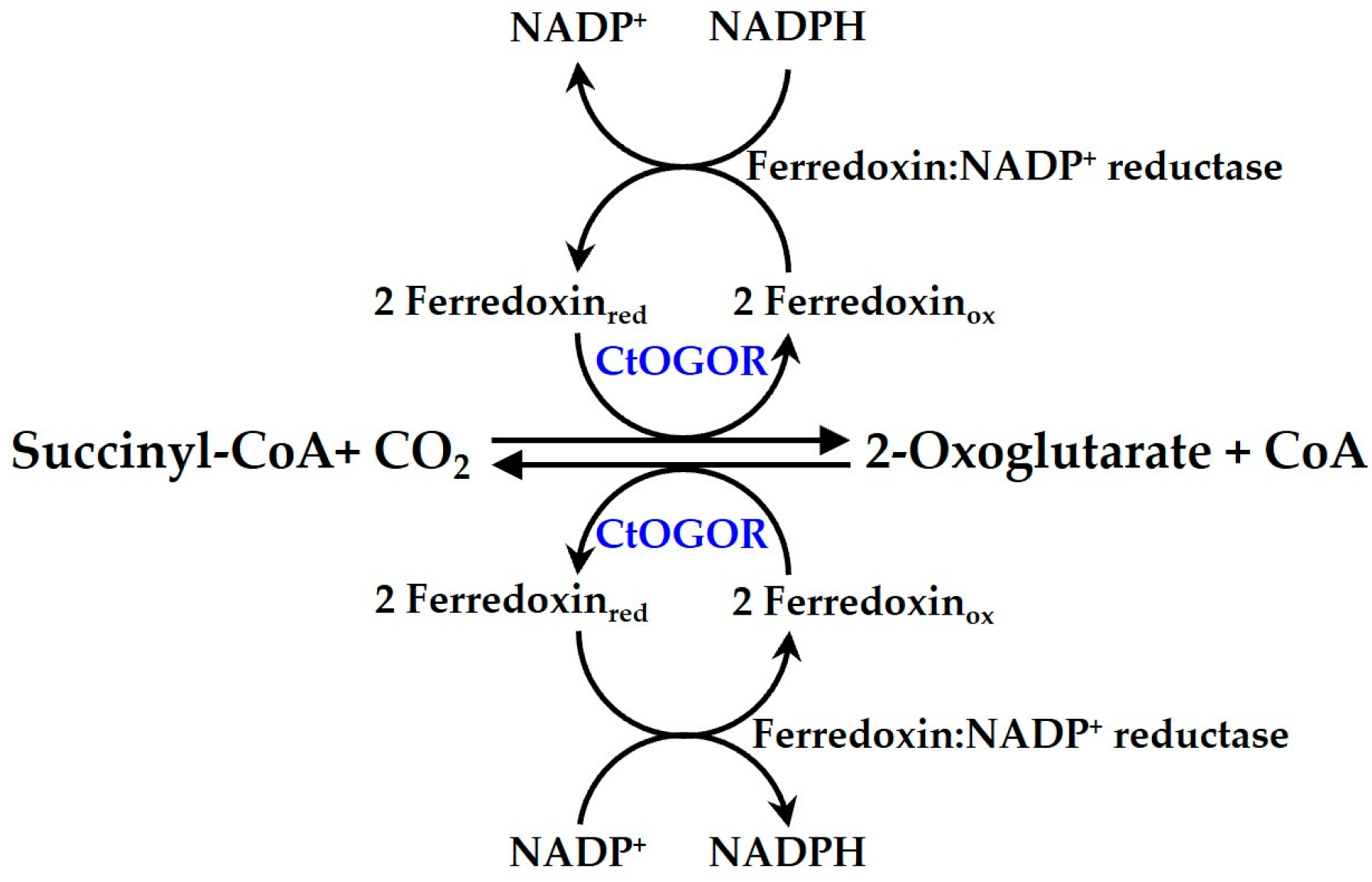
2.3. Purification of Recombinant Enzymes from E. coli
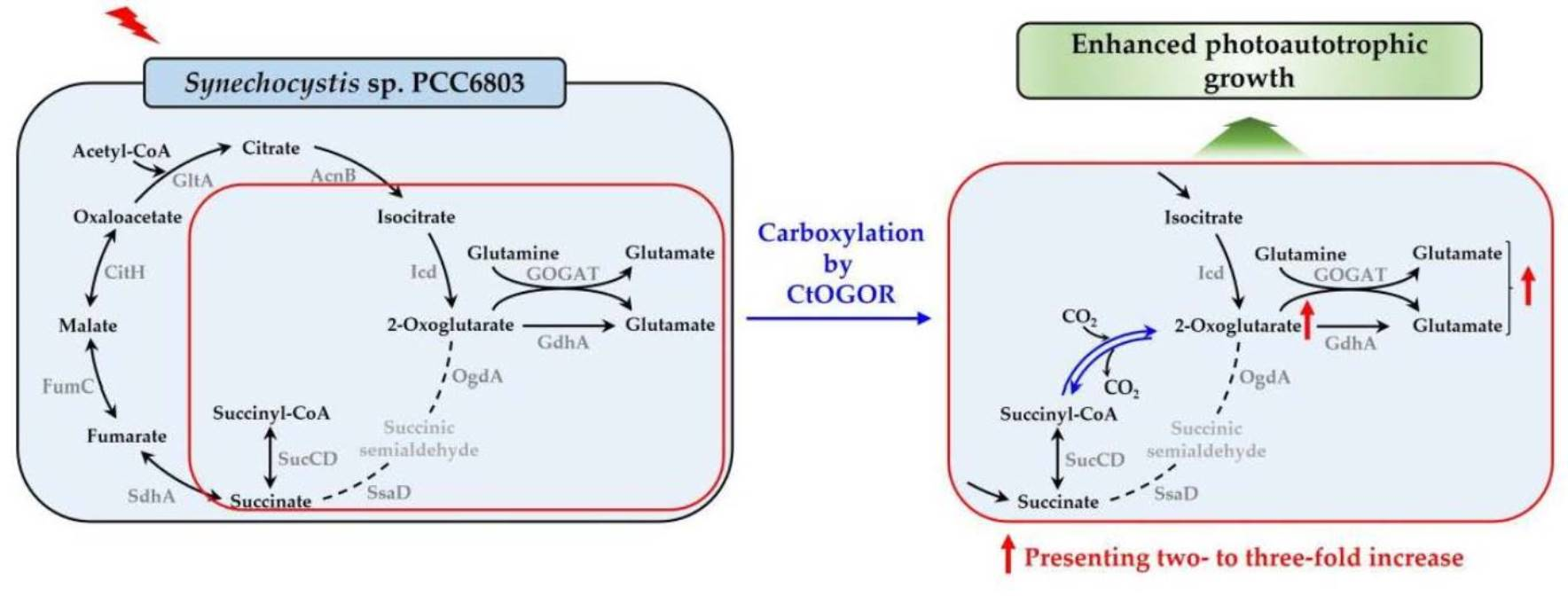
2.4. Reconstitution and Determination of Cofactors in CtOGOR
2.5. Kinetic Analyses of Enzymes
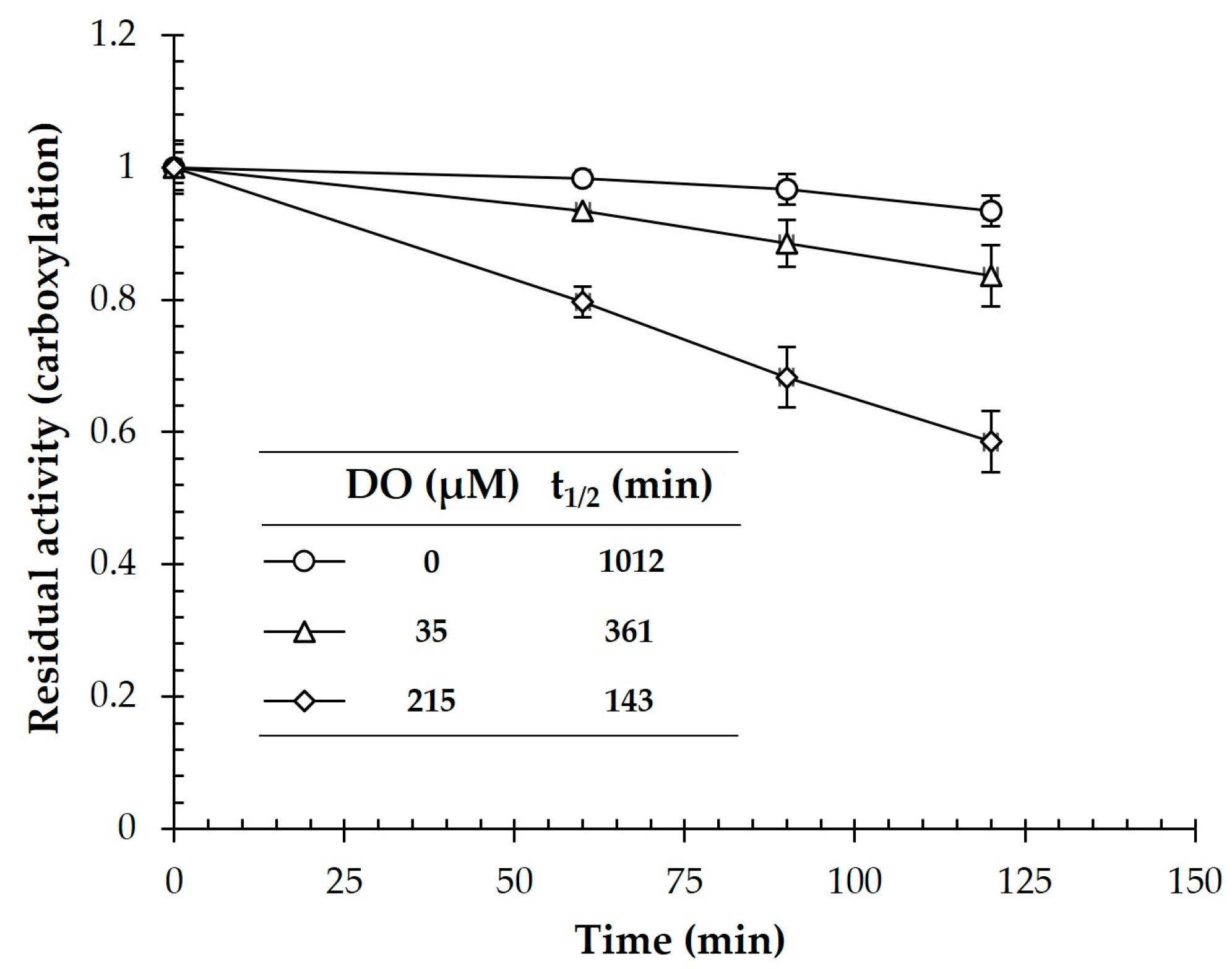
2.6. Assessment of O2 Stability of CtOGOR
2.7. Total RNA Extraction and Quantitative Reverse Transcription PCR (RT-qPCR)
2.8. Preparation and Spectral Analysis of the Membrane Fraction
2.9. Determination of Cellular Enzyme Activity
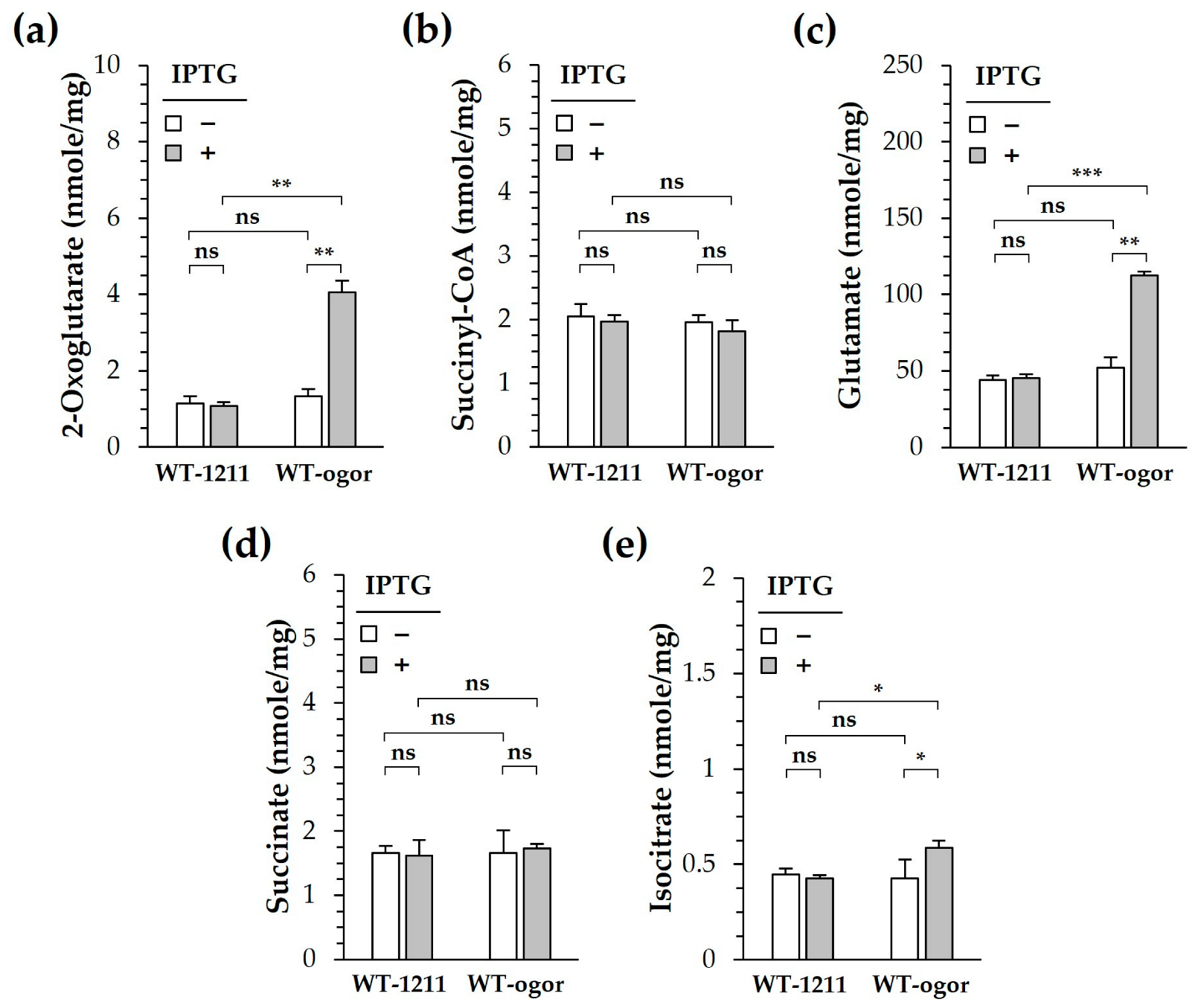
2.10. Determination of Metabolite Levels
3. Results
3.1. Biochemical Characterization of CtOGOR
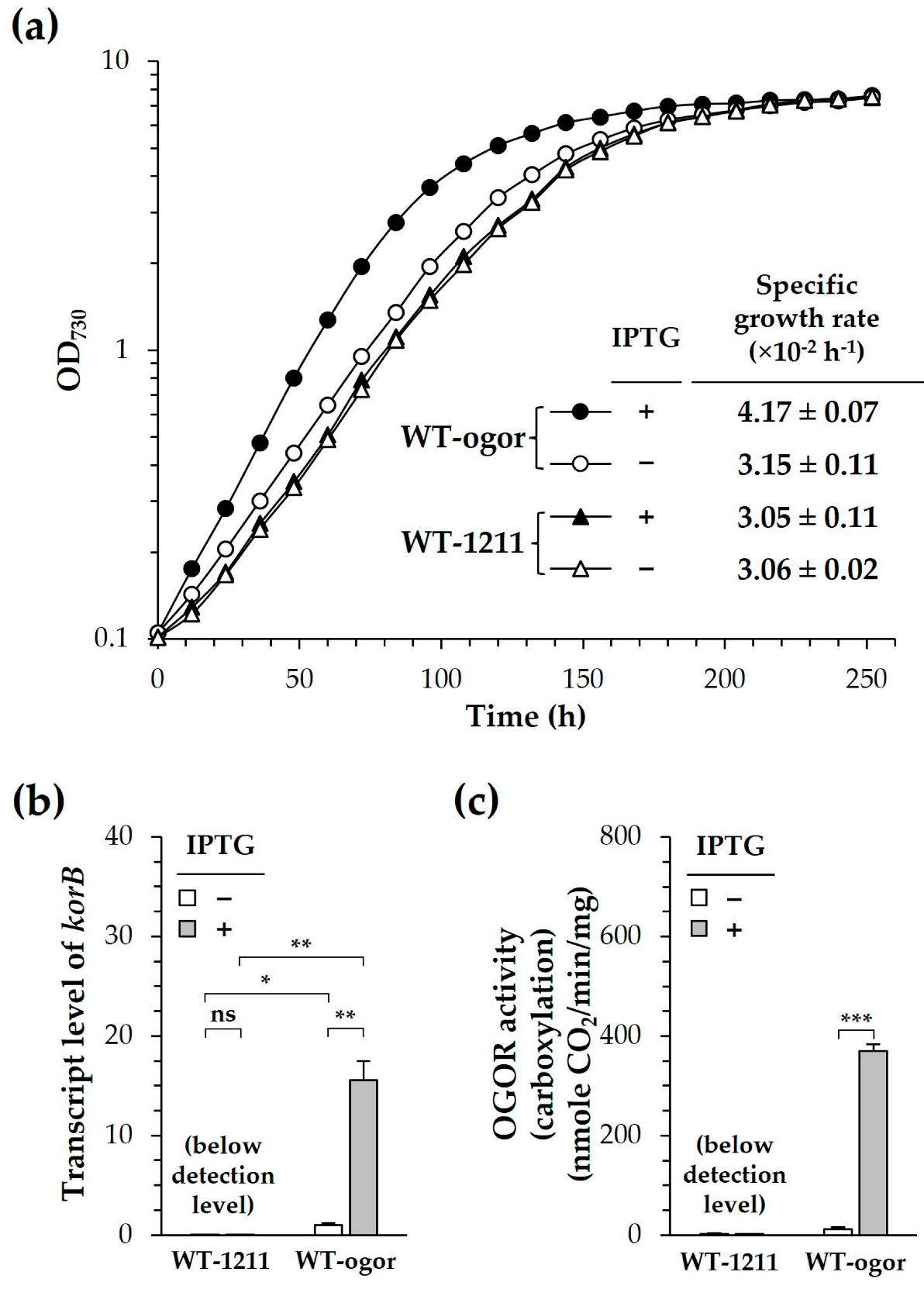
3.2. Heterologous Production of CtOGOR Enhanced Photoautotrophic Growth Rate of Synechocystis by Approximately >30%
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tang, K.H.; Blankenship, R.E. Both forward and reverse TCA cycles operate in green sulfur bacteria. J. Biol. Chem. 2010, 285, 35848–35854. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, G. Alternative pathways of carbon dioxide fixation: Insights into the early evolution of life? Annu. Rev. Microbiol. 2011, 65, 631–658. [Google Scholar] [CrossRef] [PubMed]
- Kerscher, L.; Oesterhelt, D. Purification and Properties of Two 2-Oxoacid: Ferredoxin Oxidoreductases from Halobacterium halobium. Eur. J. Biochem. 1981, 116, 587–594. [Google Scholar] [CrossRef] [PubMed]
- Mai, X.; Adams, M.W.W. Characterization of a Fourth Type of 2-Keto Acid-Oxidizing Enzyme from a Hyperthermophilic Archaeon: 2-Ketoglutarate Ferredoxin Oxidoreductase from Thermococcus litoralis. J. Bacteriol. 1996, 178, 5890–5896. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Iwasaki, T.; Wakagi, T.; Oshima, T. 2-Oxoacid:Ferredoxin Oxidoreductase from the Thermoacidophilic Archaeon, Sulfolobus sp. Strain 7. J. Biochem. 1996, 120, 587–599. [Google Scholar] [CrossRef] [PubMed]
- Dörner, E.; Boll, M. Properties of 2-oxoglutarate:ferredoxin oxidoreductase from Thauera aromatica and its role in enzymatic reduction of the aromatic ring. J. Bacteriol. 2002, 184, 3975–3983. [Google Scholar] [CrossRef]
- Yan, Z.; Maruyama, A.; Arakawa, T.; Fushinobu, S.; Wakagi, T. Crystal structures of archaeal 2-oxoacid:ferredoxin oxidoreductases from Sulfolobus tokodaii. Sci. Rep. 2016, 6, 33061. [Google Scholar] [CrossRef]
- Chen, P.Y.; Li, B.; Drennan, C.L.; Elliott, S.J. A reverse TCA cycle 2-oxoacid:ferredoxin oxidoreductase that makes C-C bonds from CO2. Joule 2019, 3, 595–611. [Google Scholar] [CrossRef]
- Pieulle, L.; Magro, V.; Hatchikian, E.C. Isolation and Analysis of the Gene Encoding the Pyruvate-Ferredoxin Oxidoreductase of Desulfovibrio africanus, Production of the Recombinant Enzyme in Escherichia coli, and Effect of Carboxy-Terminal Deletions on Its Stability. J. Bacteriol. 1997, 179, 5684–5692. [Google Scholar] [CrossRef]
- Santos-Merino, M.; Singh, A.K.; Ducat, D.C. New Applications of Synthetic Biology Tools for Cyanobacterial Metabolic Engineering. Front. Bioeng. Biotechnol. 2019, 7, 33. [Google Scholar] [CrossRef]
- Spreitzer, R.J. Questions about the complexity of chloroplast ribulose-1,5-bisphosphate carboxylase/oxygenase. Photosynth. Res. 1999, 60, 29–42. [Google Scholar] [CrossRef]
- McNevin, D.; von Caemmerer, S.; Farquhar, G. Determining RuBisCO activation kinetics and other rate and equilibrium constants by simultaneous multiple non-linear regression of a kinetic model. J. Exp. Bot. 2006, 57, 3883–3900. [Google Scholar] [CrossRef] [PubMed]
- Marcus, Y.; Altman-Gueta, H.; Wolff, Y.; Gurevitz, M. Rubisco mutagenesis provides new insight into limitations on photosynthesis and growth in Synechocystis PCC6803. J. Exp. Bot. 2011, 62, 4173–4182. [Google Scholar] [CrossRef] [PubMed]
- Marcus, Y.; Altman-Gueta, H.; Finkler, A.; Gurevitz, M. Mutagenesis at two distinct phosphate-binding sites unravels their differential roles in regulation of Rubisco activation and catalysis. J. Bacteriol. 2005, 187, 4222–4228. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Liang, F.; Lindberg, P.; Lindblad, P. Engineering photoautotrophic carbon fixation for enhanced growth and productivity. Sustain. Energy Fuels 2018, 2, 2583–2600. [Google Scholar] [CrossRef]
- Liang, F.; Lindblad, P. Synechocystis PCC 6803 overexpressing RuBisCO grow faster with increased photosynthesis. Metab. Eng. Commun. 2017, 4, 29–36. [Google Scholar] [CrossRef]
- Liang, F.; Lindblad, P. Effects of overexpressing photosynthetic carbon flux control enzymes in the cyanobacterium Synechocystis PCC 6803. Metab. Eng. 2016, 38, 56–64. [Google Scholar] [CrossRef]
- Nakajima, T.; Yoshikawa, K.; Toya, Y.; Matsuda, F.; Shimizu, H. Metabolic Flux Analysis of the Synechocystis sp. PCC 6803 ΔnrtABCD Mutant Reveals a Mechanism for Metabolic Adaptation to Nitrogen-Limited Conditions. Plant Cell Physiol. 2017, 58, 537–545. [Google Scholar] [CrossRef]
- Kratz, W.A.; Myers, J. Nutrition and Growth of Several Blue-Green Algae. Am. J. Bot. 1955, 42, 282–287. [Google Scholar] [CrossRef]
- Green, M.R.; Sambrook, J. Molecular Cloning: A Laboratory Manual, 4th ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2012. [Google Scholar]
- Eraso, J.M.; Kaplan, S. prrA, a Putative Response Regulator Involved in Oxygen Regulation of Photosynthesis Gene Expression in Rhodobacter sphaeroides. J. Bacteriol. 1994, 176, 32–43. [Google Scholar] [CrossRef]
- Ng, W.O.; Zentella, R.; Wang, Y.; Taylor, J.S.; Pakrasi, H.B. phrA, the major photoreactivating factor in the cyanobacterium Synechocystis sp. strain PCC 6803 codes for a cyclobutane-pyrimidine-dimer-specific DNA photolyase. Arch. Microbiol. 2000, 173, 412–417. [Google Scholar] [CrossRef] [PubMed]
- Simon, R.; Priefer, U.; Pühler, A. A Broad Host Range Mobilization System for In Vivo Genetic Engineering: Transposon Mutagenesis in Gram Negative Bacteria. Nat. Biotechnol. 1983, 1, 784–791. [Google Scholar] [CrossRef]
- Kim, E.-J.; Kim, J.-S.; Rhee, H.J.; Lee, J.K. Growth arrest of Synechocystis sp. PCC6803 by superoxide generated from heterologously expressed Rhodobacter sphaeroides chlorophyllide a reductase. FEBS Lett. 2009, 583, 219–223. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Kim, H.; Lee, J.K. Biochemical characterization of protoporphyrinogen dehydrogenase and protoporphyrin ferrochelatase of Vibrio vulnificus and the critical complex formation between these enzymes. Biochim. Biophys. Acta Gen. Subj. 2018, 1862, 2674–2687. [Google Scholar] [CrossRef]
- Kurihara, T.; Mihara, H.; Kato, S.; Yoshimura, T.; Esaki, N. Assembly of iron–sulfur clusters mediated by cysteine desulfurases, IscS, CsdB and CSD, from Escherichia coli. Biochim. Biophys. Acta 2003, 1647, 303–309. [Google Scholar] [CrossRef]
- Lovenberg, W.; Buchanan, B.B.; Rabinowitz, J.C. Studies on the Chemical Nature of Clostridial Ferredoxin. J. Biol. Chem. 1963, 238, 3899–3913. [Google Scholar] [CrossRef]
- Beinert, H. Semi-micro Methods for Analysis of Labile Sulfide and of Labile Sulfide plus Sulfane Sulfur in Unusually Stable Iron-Sulfur Proteins. Anal. Biochem. 1983, 131, 373–378. [Google Scholar] [CrossRef]
- Fraccascia, P.; Sniekers, M.; Casteels, M.; Van Veldhoven, P.P. Presence of thiamine pyrophosphate in mammalian peroxisomes. BMC Biochem. 2007, 8, 10. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein Measurement with the Folin Phenol Reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Leader, D.P. A method of introducing the physiological carbon dioxide-bicarbonate buffer system to medical students. Biochem. Educ. 1979, 7, 37–38. [Google Scholar] [CrossRef]
- Lan, Y.; Mott, K.A. Determination of Apparent Km Values for Ribulose 1,5-Bisphosphate Carboxylase/Oxygenase (Rubisco) Activase Using the Spectrophotometric Assay of Rubisco Activity. Plant Physiol. 1991, 95, 604–609. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.-J.; Kim, J.-S.; Lee, I.-H.; Rhee, H.J.; Lee, J.K. Superoxide generation by chlorophyllide a reductase of Rhodobacter sphaeroides. J. Biol. Chem. 2008, 283, 3718–3730. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, R.E.; Erstad, S.M.; Ramos-Martinez, E.M.; Fimognari, L.; De Porcellinis, A.J.; Sakuragi, Y. An easy and efficient permeabilization protocol for in vivo enzyme activity assays in cyanobacteria. Microb. Cell Fact. 2016, 15, 186. [Google Scholar] [CrossRef] [PubMed]
- Jordan, P.M.; Laghai-Newton, A. [52] Purification of 5-aminolevulinate synthase. Methods Enzymol. 1986, 123, 435–443. [Google Scholar] [CrossRef] [PubMed]
- Page, L.E.; Liberton, M.; Pakrasi, H.B. Reduction of Photoautotrophic Productivity in the Cyanobacterium Synechocystis sp. Strain PCC 6803 by Phycobilisome Antenna Truncation. Appl. Environ. Microbiol. 2012, 78, 6349–6351. [Google Scholar] [CrossRef]
- Knoop, H.; Gründel, M.; Zilliges, Y.; Lehmann, R.; Hoffmann, S.; Lockau, W.; Steuer, R. Flux Balance Analysis of Cyanobacterial Metabolism: The Metabolic Network of Synechocystis sp. PCC 6803. PLoS Comput. Biol. 2013, 9, e1003081. [Google Scholar] [CrossRef]
- Mills, L.A.; McCormick, A.J.; Lea-Smith, D.J. Current knowledge and recent advances in understanding metabolism of the model cyanobacterium Synechocystis sp. PCC 6803. Biosci. Rep. 2020, 40, BSR20193325. [Google Scholar] [CrossRef]
- Muro-Pastor, M.I.; Florencio, F.J. Purification and properties of NADP-isocitrate dehydrogenase from the unicellular cyanobacterium Synechocystis sp. PCC 6803. Eur. J. Biochem. 1992, 203, 99–105. [Google Scholar] [CrossRef]
- Cassier-Chauvat, C.; Chauvat, F. Function and Regulation of Ferredoxins in the Cyanobacterium, Synechocystis PCC6803: Recent Advances. Life 2014, 4, 666–680. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, X.; Spengler, K.; Terberger, K.; Boehm, M.; Appel, J.; Barske, T.; Timm, S.; Battchikova, N. Pyruvate:ferredoxin oxidoreductase and low abundant ferredoxins support aerobic photomixotrophic growth in cyanobacteria. eLife 2022, 11, e71339. [Google Scholar] [CrossRef] [PubMed]
- Schorsch, M.; Kramer, M.; Goss, T.; Eisenhut, M.; Robinson, N.; Osman, D.; Wilde, A.; Sadaf, S.; Brückler, H.; Walder, L.; et al. A unique ferredoxin acts as a player in the low-iron response of photosynthetic organisms. Proc. Natl. Acad. Sci. USA 2018, 115, E12111–E12120. [Google Scholar] [CrossRef] [PubMed]
- Kihara, S.; Hartzler, D.A.; Savikhin, S. Oxygen concentration inside a functioning photosynthetic cell. Biophys. J. 2014, 106, 1882–1889. [Google Scholar] [CrossRef] [PubMed]
- Durall, C.; Rukminasari, N.; Lindblad, P. Enhanced growth at low light intensity in the cyanobacterium Synechocystis PCC 6803 by overexpressing phosphoenolpyruvate carboxylase. Algal Res. 2016, 16, 275–281. [Google Scholar] [CrossRef]
- Durall, C.; Lindberg, P.; Yu, J.; Lindblad, P. Increased ethylene production by overexpressing phosphoenolpyruvate carboxylase in the cyanobacterium Synechocystis PCC 6803. Biotechnol. Biofuels 2020, 13, 16. [Google Scholar] [CrossRef]
- Fathy, W.; Essawy, E.; Tawfik, E.; Khedr, M.; Abdelhameed, M.S.; Hammouda, O.; Elsayed, K. Recombinant overexpression of the Escherichia coli acetyl-CoA carboxylase gene in Synechocystis sp. boosts lipid production. J. Basic Microbiol. 2021, 61, 330–338. [Google Scholar] [CrossRef]
- Tripathy, J.N.; Hirasawa, M.; Sutton, R.B.; Dasgupta, A.; Vaidyanathan, N.; Zabet-Moghaddam, M.; Florencio, F.J.; Srivastava, A.P.; Knaff, D.B. A loop unique to ferredoxin-dependent glutamate synthases is not absolutely essential for ferredoxin-dependent catalytic activity. Photosynth. Res. 2015, 123, 129–139. [Google Scholar] [CrossRef]
- Chávez, S.; Candau, P. An NAD-specific glutamate dehydrogenase from cyanobacteria. Identification and properties. FEBS Lett. 1991, 285, 35–38. [Google Scholar] [CrossRef]
- Wang, X.; Lei, G.; Wu, X.; Wang, F.; Lai, C.; Li, Z. Expression, purification and characterization of sll1981 protein from cyanobacterium Synechocystis sp. PCC6803. Protein Expr. Purif. 2017, 139, 21–28. [Google Scholar] [CrossRef]
- Forchhammer, K.; Selim, K.A. Carbon/nitrogen homeostasis control in cyanobacteria. FEMS Microbiol. Rev. 2020, 44, 33–53. [Google Scholar] [CrossRef]
- Fokina, O.; Chellamuthu, V.R.; Forchhammer, K.; Zeth, K. Mechanism of 2-oxoglutarate signaling by the Synechococcus elongatus PII signal transduction protein. Proc. Natl. Acad. Sci. USA 2010, 107, 19760–19765. [Google Scholar] [CrossRef] [PubMed]
- Wolk, C.P. Physiology and Cytological Chemistry of Blue-Green Algae. Bacteriol. Rev. 1973, 37, 32–101. [Google Scholar] [CrossRef] [PubMed]
- Walker, M.C.; van der Donk, W.A. The many roles of glutamate in metabolism. J. Ind. Microbiol. Biotechnol. 2016, 43, 419–430. [Google Scholar] [CrossRef] [PubMed]
- Cameron, J.C.; Pakrasi, H.B. Glutathione in Synechocystis 6803: A closer look into the physiology of a ΔgshB mutant. Plant Signal. Behav. 2011, 6, 89–92. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Content | Number per αβ-Protomer * |
|---|---|
| Fe | 7.51 ± 0.51 |
| S ** | 7.65 ± 0.12 |
| TPP | 0.77 ± 0.07 |
| Direction of Reaction | Substrate | Km (mM) | kcat (s−1) * | kcat/Km (s−1 mM−1) |
|---|---|---|---|---|
| Carboxylation | Succinyl-CoA | 0.024 ± 0.002 | 2.6 ± 0.1 | 110.2 ± 3.5 |
| CO2 ** | 0.468 ± 0.045 | 2.7 ± 0.1 | 6.5 ± 0.2 | |
| Decarboxylation | 2-Oxoglutarate | 1.689 ± 0.092 | 8.4 ± 0.1 | 5.0 ± 0.1 |
| CoA | 0.042 ± 0.015 | 8.1 ± 0.2 | 191.7 ± 2.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.; Oh, E.K.; Kim, E.-J.; Lee, J.K. Photoautotrophic Growth Rate Enhancement of Synechocystis sp. PCC6803 by Heterologous Production of 2-Oxoglutarate:Ferredoxin Oxidoreductase from Chlorobaculum tepidum. Biology 2023, 12, 59. https://doi.org/10.3390/biology12010059
Kim J, Oh EK, Kim E-J, Lee JK. Photoautotrophic Growth Rate Enhancement of Synechocystis sp. PCC6803 by Heterologous Production of 2-Oxoglutarate:Ferredoxin Oxidoreductase from Chlorobaculum tepidum. Biology. 2023; 12(1):59. https://doi.org/10.3390/biology12010059
Chicago/Turabian StyleKim, June, Eun Kyoung Oh, Eui-Jin Kim, and Jeong K. Lee. 2023. "Photoautotrophic Growth Rate Enhancement of Synechocystis sp. PCC6803 by Heterologous Production of 2-Oxoglutarate:Ferredoxin Oxidoreductase from Chlorobaculum tepidum" Biology 12, no. 1: 59. https://doi.org/10.3390/biology12010059
APA StyleKim, J., Oh, E. K., Kim, E.-J., & Lee, J. K. (2023). Photoautotrophic Growth Rate Enhancement of Synechocystis sp. PCC6803 by Heterologous Production of 2-Oxoglutarate:Ferredoxin Oxidoreductase from Chlorobaculum tepidum. Biology, 12(1), 59. https://doi.org/10.3390/biology12010059