
Biology, Genetic Diversity, and Conservation of Wild Bees in Tree Fruit Orchards


 ,
,  ,
, 
 ,
, 
Abstract
Simple Summary
Abstract

1. Introduction
2. Types of Bees Found in Orchards
2.1. Based on Nesting Behavior

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Nesting Behavior | Foraging Specialization | Social Behavior |
|---|---|---|---|
| Agapostemon virescens | Ground | Generalist | Solitary |
| Andrena barbara | Ground | Generalist | Solitary |
| Andrena barbilabris | Ground | Generalist | Solitary |
| Andrena bisalicis | Ground | Generalist | Solitary |
| Andrena bradleyi | Ground | Vaccinium specialist | Solitary |
| Andrena carlini | Ground | Generalist | Solitary |
| Andrena carolina | Ground | Vaccinium specialist | Solitary |
| Andrena cineraria | Ground | Generalist | Solitary |
| Andrena commoda | Ground | Generalist | Solitary |
| Andrena crataegi | Ground | Generalist | Solitary |
| Andrena cressonii | Ground | Generalist | Solitary |
| Andrena dunning | Ground | Generalist | Solitary |
| Andrena forbesii | Ground | Generalist | Solitary |
| Andrena hippotes | Ground | Generalist | Solitary |
| Andrena imitatrix | Ground | Generalist | Solitary |
| Andrena mandibularis | Ground | Generalist | Solitary |
| Andrena miserabilis | Ground | Generalist | Solitary |
| Andrena nasonii | Ground | Generalist | Solitary |
| Andrena nuda | Ground | Generalist | Solitary |
| Andrena perplexa | Ground | Generalist | Solitary |
| Andrena pruni | Ground | Generalist | Solitary |
| Andrena rugosa | Ground | Generalist | Solitary |
| Andrena tridens | Ground | Generalist | Solitary |
| Andrena vicina | Ground | Generalist | Solitary |
| Andrena violae | Ground | Viola specialist | Solitary |
| Andrena wilkella | Ground | Generalist | Solitary |
| Anthophora abrupta | Ground | Generalist | Solitary |
| Augochlora pura | Wood | Generalist | Solitary |
| Augochloropsis metallica | Ground | Generalist | Primitively Eusocial |
| Bombus balteatus | Cavity | Generalist | Eusocial |
| Bombus bifarius | Cavity | Generalist | Eusocial |
| Bombus bimaculatus | Cavity | Generalist | Eusocial |
| Bombus flavifrons | Cavity | Generalist | Eusocial |
| Bombus griseocollis | Cavity | Generalist | Eusocial |
| Bombus impatiens | Cavity | Generalist | Eusocial |
| Bombus perplexa | Cavity | Generalist | Eusocial |
| Bombus sylvicola | Cavity | Generalist | Eusocial |
| Bombus terrestris | Cavity | Generalist | Eusocial |
| Bombus vagans | Cavity | Generalist | Eusocial |
| Ceratina calcarata | Pith | Generalist | Sub-social |
| Ceratina dupla | Pith | Generalist | Sub-social |
| Ceratina strenua | Pith | Generalist | Sub-social |
| Colletes validus | Ground | Vaccinium specialist | Solitary |
| Habropoda laboriosa | Ground | Vaccinium specialist | Solitary |
| Halictus confusus | Ground | Generalist | Primitively Eusocial |
| Halictus ligatus | Ground | Generalist | Primitively Eusocial |
| Halictus rubicundus | Ground | Generalist | Sub-social |
| Hoplitis adunca | Tunnel | Boraginaceae specialist | Solitary |
| Hylaeus punctulatissimus | Ground | Lamiaceae specialist | Solitary |
| Lasioglossum admirandum | Ground | Generalist | Primitively Eusocial |
| Lasioglossum cressonii | Wood | Generalist | Primitively Eusocial |
| Lasioglossum foxii | Ground | Generalist | Solitary |
| Lasioglossum imitatum | Ground | Generalist | Primitively Eusocial |
| Lasioglossum pilosum | Ground | Generalist | Primitively Eusocial |
| Lasioglossum quebecense | Ground | Generalist | Solitary |
| Lasioglossum tegulare | Ground | Generalist | Primitively Eusocial |
| Lasioglossum truncatum | Ground | Generalist | Primitively Eusocial |
| Lasioglossum versans | Ground | Generalist | Primitively Eusocial |
| Lasioglossum versatum | Ground | Generalist | Primitively Eusocial |
| Lasioglossum vierecki | Ground | Generalist | Primitively Eusocial |
| Lasioglossum zephyrus | Ground | Generalist | Primitively Eusocial |
| Megachile addenda | Ground | Fabaceae specialist | Solitary |
| Melitta americana | Ground | Vaccinium specialist | Solitary |
| Osmia apicata | Tunnel | Onosma specialist | Solitary |
| Osmia atriventris | Tunnel | Generalist | Solitary |
| Osmia bicornis | Tunnel | Rosaceae specialist | Solitary |
| Osmia californica | Tunnel | Generalist | Solitary |
| Osmia cerinthidis | Tunnel | Boraginaceae specialist | Solitary |
| Osmia cornifrons | Tunnel | Generalist | Solitary |
| Osmia cornuta | Tunnel | Generalist | Solitary |
| Osmia lignaria | Tunnel | Generalist | Solitary |
| Osmia maxillaris | Tunnel | Fabaceae specialist | Solitary |
| Osmia maxschwarzi | Tunnel | Fabaceae specialist | Solitary |
| Osmia montana | Tunnel | Generalist | Solitary |
| Osmia pumila | Tunnel | Generalist | Solitary |
| Osmia scheherazade | Tunnel | Fabaceae specialist | Solitary |
| Osmia taurus | Tunnel | Generalist | Solitary |
| Osmia virga | Tunnel | Vaccinium specialist | Solitary |
| Peponapis pruinosa | Ground | Cucurbita specialist | Solitary |
| Xylocopa virginica | Wood | Generalist | Semi-social |
2.2. Based on Foraging Preferences
3. Bioecology of Major Species
3.1. Mating Behavior
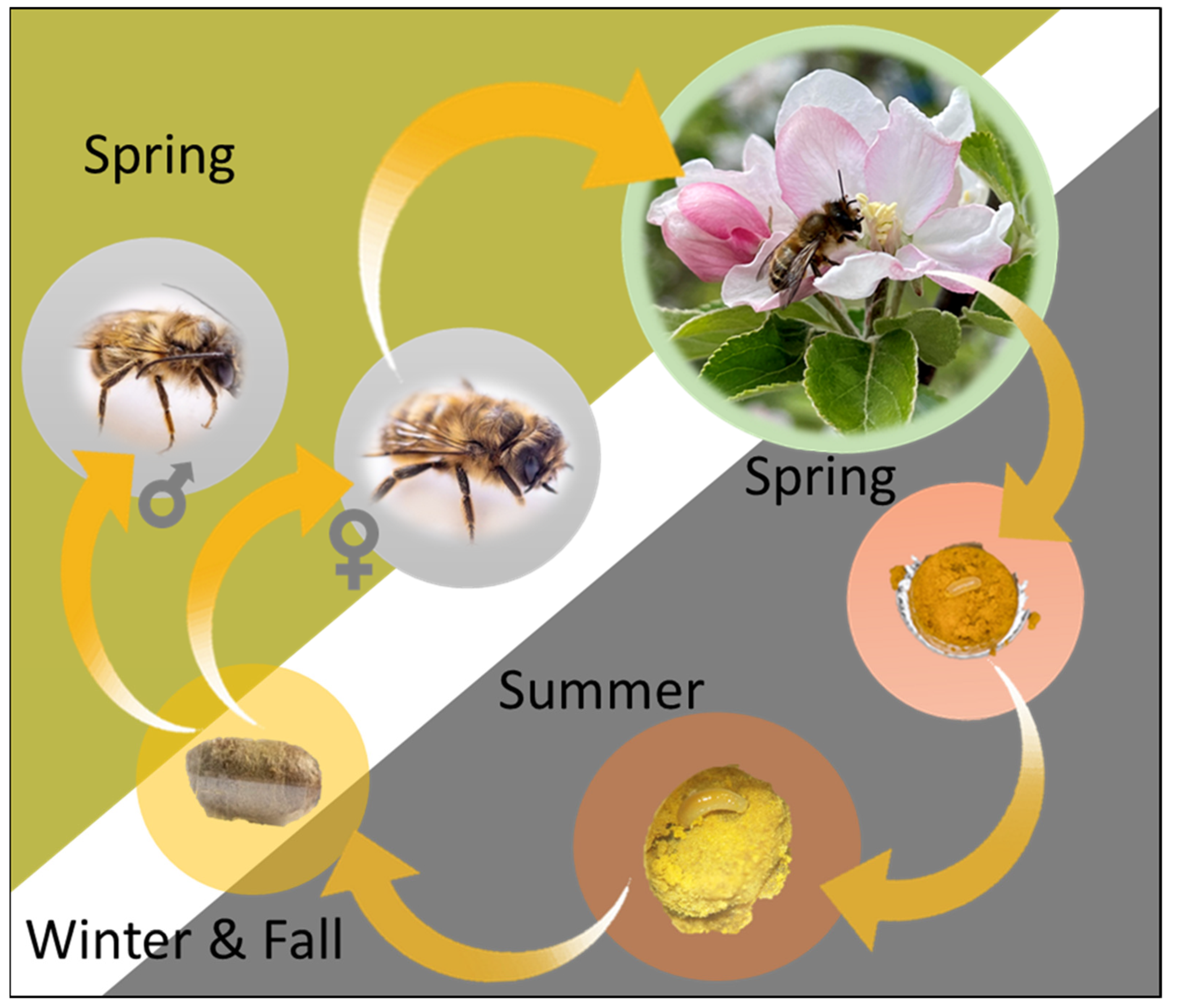
3.2. Life Cycle
3.3. Foraging Behavior of Orchard Bees
| Species | Max Reported Range (m) | Average Reported Range (m) | Bee Size (mm) | Number of Studies |
|---|---|---|---|---|
| Andrena cineraria | 300 | 300 | 10–14 | 1 |
| Andrena vaga | 600 | 555 | 11–15 | 2 |
| Andrena barbilabris | 530 | 515 | 10–12 | 2 |
| Anthophora abrupta | 12,500 | 12,500 | 14–17 | 1 |
| Bombus balteatus | 220.5 | 85.4 | 11–14 | 1 |
| Bombus bifarius | 220.5 | 110.25 | 8–14 | 1 |
| Bombus flavifrons | 202.5 | 23.8 | 9–12 | 1 |
| Bombus sylvicola | 290.5 | 74.7 | 10–14 | 1 |
| Bombus terrestris | 2800 | 1137.6 | 20–23 | 8 |
| Bombus terrestris (male) | 9900 | 9900 | 20–23 | 1 |
| Hoplitis adunca | 1400 | 1400 | 8–12 | 1 |
| Hylaeus punctulatissimus | 1100 | 1100 | 6–8 | 1 |
| Osmia lignaria | 600 | 600 | 11–12 | 1 |
3.4. Orchard Crop Preference (Food Sources for Adults and Their Offspring)
3.5. Time Matching with Flowering Period
4. Diversity of Bees in Orchards
Genetic Diversity of Orchard Bees
5. Environmental Threats and Multiple Stressors Affecting Orchard Bee Communities
5.1. Pesticide Exposure and Hazard
5.2. Arthropod Pests and Other Natural Enemies
5.3. Parasites and Diseases
5.4. Lack of Nesting Habitats
5.5. Lack of Diverse Floral Food Resources
6. Management and Conservation of Orchard Bees
6.1. Strategies for Enhancing Orchard Bee Nesting Habitats
6.1.1. Ground-Nesting Orchard Bees
6.1.2. Cavity and Tunnel-Nesting Orchard Bees
6.2. Proximity of Forest Habitat Improves Nesting Conditions for Orchard Bees
6.3. Building Artificial Nests for Orchard Bees
6.4. Establishment of Native Floral Plantings to Support Orchard Bee Communities
6.5. Protecting Orchard Bees from Pesticide Hazards
7. Conclusions and Recommendations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eilers, E.J.; Kremen, C.; Greenleaf, S.S.; Garber, S.K.; Klein, A.-M. Contribution of pollinator-mediated crops to nutrients in the human food supply. PLoS ONE 2011, 6, e21363. [Google Scholar] [CrossRef]
- Fenech, M.; Amaya, I.; Valpuesta, V.; Botella, M.A. Vitamin C content in fruits: Biosynthesis and regulation. Front. Plant Sci. 2019, 9, 2006. [Google Scholar] [CrossRef]
- Johnson, J.D.; Pettis, J.S. A survey of imidacloprid levels in water sources potentially frequented by honeybees (Apis mellifera) in the Eastern USA. Water Air Soil Pollut. 2014, 225, 2127. [Google Scholar] [CrossRef]
- Pitts-Singer, T.L.; Artz, D.R.; Peterson, S.S.; Boyle, N.K.; Wardell, G.I. Examination of a managed pollinator strategy for almond production using Apis mellifera (Hymenoptera: Apidae) and Osmia lignaria (Hymenoptera: Megachilidae). Environ. Entomol. 2018, 47, 364–377. [Google Scholar] [CrossRef]
- Da Santos, R.S.; Carneiro, L.T.; de Oliveira Santos, J.P.; da Silva, M.M.; de Oliveira Milfont, M.; Castro, C.C. Bee pollination services and the enhancement of fruit yield associated with seed number in self-incompatible tangelos. Sci. Hortic. 2021, 276, 109743. [Google Scholar] [CrossRef]
- Gallai, N.; Salles, J.-M.; Settele, J.; Vaissière, B.E. Economic valuation of the vulnerability of world agriculture confronted with pollinator decline. Ecol. Econ. 2009, 68, 810–821. [Google Scholar] [CrossRef]
- Losey, J.E.; Vaughan, M. The economic value of ecological services provided by insects. Bioscience 2006, 56, 311–323. [Google Scholar] [CrossRef]
- Wade, A.; Lin, C.-H.; Kurkul, C.; Regan, E.R.; Johnson, R.M. Combined toxicity of insecticides and fungicides applied to California almond orchards to honey bee larvae and adults. Insects 2019, 10, 20. [Google Scholar] [CrossRef]
- Belsky, J.; Joshi, N.K. Impact of biotic and abiotic stressors on managed and feral bees. Insects 2019, 10, 233. [Google Scholar] [CrossRef]
- Maeta, Y. Utilization of Wild Bees. Farming Jpn. 1990, 24, 13–22. [Google Scholar]
- Batra, S.W.T. Diversify Will Pollen Bees. Am. Bee J. 1994, 134, 591–594. [Google Scholar]
- Greer, L. Alternative Pollinators: Native Bees; ATTRA—National Center for Appropriate Technology: Fayetteville, AR, USA, 1999. [Google Scholar]
- Park, M.G.; Orr, M.C.; Danforth, B.N. The role of native bees in apple pollination. N. Y. Fruit Quart 2010, 18, 21–25. [Google Scholar]
- Moisset, B.B.; Buchmann, S.L. Bee Basics: An Introduction to Our Native Bees; Stritch, L., Nelson, J., Prendusi, T., Adams, L.D., Eds.; USDA Forest Service and Pollinator Partnership: Washington, DC, USA, 2011.
- Redhead, J.W.; Dreier, S.; Bourke, A.F.G.; Heard, M.S.; Jordan, W.C.; Sumner, S.; Wang, J.; Carvell, C. Effects of habitat composition and landscape structure on worker foraging distances of five bumble bee species. Ecol. Appl. 2016, 26, 726–739. [Google Scholar] [CrossRef]
- Hoehn, P.; Tscharntke, T.; Tylianakis, J.M.; Steffan-Dewenter, I. Functional group diversity of bee pollinators increases crop yield. Proc. R. Soc. B 2008, 275, 2283–2291. [Google Scholar] [CrossRef]
- Cameron, S.A.; Lozier, J.D.; Strange, J.P.; Koch, J.B.; Cordes, N.; Solter, L.F.; Griswold, T.L. Patterns of widespread decline in North American bumble bees. Proc Natl Acad Sci USA 2011, 108, 662–667. [Google Scholar] [CrossRef]
- Colla, S.R.; Packer, L. Evidence for decline in eastern North American bumblebees (Hymenoptera: Apidae), with special focus on Bombus affinis Cresson. Biodivers. Conserv. 2008, 17, 1379–1391. [Google Scholar] [CrossRef]
- Kerr, J.T.; Pindar, A.; Galpern, P.; Packer, L.; Potts, S.G.; Roberts, S.M.; Rasmont, P.; Schweiger, O.; Colla, S.R.; Richardson, L.L.; et al. Climate change impacts on bumblebees converge across continents. Science 2015, 349, 177–180. [Google Scholar] [CrossRef]
- Turley, N.E.; Biddinger, D.J.; Joshi, N.K.; López-Uribe, M.M. Six years of wild bee monitoring shows changes in biodiversity within and across years and declines in abundance. Ecol. Evol. 2022, 12, e9190. [Google Scholar] [CrossRef]
- Roulston, T.H.; Goodell, K. The Role of Resources and Risks in Regulating Wild Bee Populations. Annu. Rev. Entomol. 2011, 56, 293–312. [Google Scholar] [CrossRef]
- Goulson, D.; Hughes, W.O.H. Mitigating the Anthropogenic Spread of Bee Parasites to Protect Wild Pollinators. Biol. Conserv. 2015, 191, 10–19. [Google Scholar] [CrossRef]
- Bauer, D.M.; Wing, I.S. The Macroeconomic Cost of Catastrophic Pollinator Declines. Ecol. Econ. 2016, 126, 1–13. [Google Scholar] [CrossRef]
- Zattara, E.E.; Aizen, M.A. Worldwide Occurrence Records Suggest a Global Decline in Bee Species Richness. One Earth 2021, 4, 114–123. [Google Scholar] [CrossRef]
- Aizen, M.A.; Garibaldi, L.A.; Cunningham, S.A.; Klein, A.M. Long-Term Global Trends in Crop Yield and Production Reveal No Current Pollination Shortage but Increasing Pollinator Dependency. Curr. Biol. 2008, 18, 1572–1575. [Google Scholar] [CrossRef]
- Brown, M.J.F.; Dicks, L.; Paxton, R.J.; Baldock, K.C.R.; Barron, A.B.; Chauzat, M.P.; Freitas, B.M.; Goulson, D.; Jepsen, S.; Kremen, C.; et al. A Horizon Scan of Future Threats and Opportunities for Pollinators and Pollination. PeerJ 2016, 2016, e2249. [Google Scholar] [CrossRef]
- Isaacs, R.; Williams, N.; Ellis, J.; Pitts-Singer, T.L.; Bommarco, R.; Vaughan, M. Integrated Crop Pollination: Combining Strategies to Ensure Stable and Sustainable Yields of Pollination-Dependent Crops. Basic Appl. Ecol. 2017, 22, 44–60. [Google Scholar] [CrossRef]
- Bosch, J.; Kemp, W.P.; Trostle, G.E. Bee Population Returns and Cherry Yields in an Orchard Pollinated with Osmia lignaria (Hymenoptera: Megachilidae). J. Econ. Entomol. 2006, 99, 408–413. [Google Scholar] [CrossRef]
- Buchmann, S.L. Buzz Pollination in Angiosperms. In Handbook of Experimental Pollination Biology; Van Nostrand Reinhold Company: New York, NY, USA, 1983; pp. 73–113. ISBN 19840215543. [Google Scholar]
- Biddinger, D.J.; Rajotte, E.G.; Joshi, N.K.; Ritz, A. Wild Bees as Alternative Pollinators in Pennsylvania Apple Orchards. Fruit Times 2011, 30, 1–4. [Google Scholar]
- Albrecht, M.; Schmid, B.; Hautier, Y.; Müller, C.B. Diverse Pollinator Communities Enhance Plant Reproductive Success. Proc. R. Soc. B 2012, 279, 4845–4852. [Google Scholar] [CrossRef]
- Bosch, J.; Sgolastra, F.; Kemp, W.P. Ecophysiology of Osmia Mason Bees Used as Crop Pollinators. In Bee Pollination in Agricultural Ecosystems; James, R., Pitts-Singer, T.L., Eds.; Oxford University Press: Oxford, UK, 2008; pp. 83–104. ISBN 9780199871575. [Google Scholar]
- Gruber, B.; Eckel, K.; Everaars, J.; Dormann, C.F. On Managing the Red Mason Bee (Osmia bicornis) in Apple Orchards. Apidologie 2011, 42, 564–576. [Google Scholar] [CrossRef]
- Kwon, H.; Kim, T.K.; Hong, S.M.; Se, E.K.; Cho, N.J.; Kyung, K.S. Effect of Household Processing on Pesticide Residues in Field-Sprayed Tomatoes. Food Sci. Biotechnol. 2015, 24, 1–6. [Google Scholar] [CrossRef]
- Horth, L.; Campbell, L.A. Supplementing Small Farms with Native Mason Bees Increases Strawberry Size and Growth Rate. J. Appl. Ecol. 2018, 55, 591–599. [Google Scholar] [CrossRef]
- Ahrenfeldt, E.J.; Sigsgaard, L.; Hansted, L.; Jensen, A.C.; Toldam-Andersen, T.B. Forage Quality and Quantity Affect Red Mason Bees and Honeybees Differently in Flowers of Strawberry Varieties. Entomol. Exp. Appl. 2019, 167, 763–773. [Google Scholar] [CrossRef]
- Eeraerts, M.; Vanderhaegen, R.; Smagghe, G.; Meeus, I. Pollination Efficiency and Foraging Behaviour of Honey Bees and Non-Apis Bees to Sweet Cherry. Agric. For. Entomol. 2020, 22, 75–82. [Google Scholar] [CrossRef]
- Batra, S.W.T. The Hornfaced Bee for Efficient Pollination of Small Farm Orchards. USDA Misc. Publ. 1982, 1422, 116–120. [Google Scholar]
- Bosch, J.; Kemp, W.P. Development and Emergence of the Orchard Pollinator Osmia lignaria (Hymenoptera: Megachilidae). Environ. Entomol. 2000, 29, 8–13. [Google Scholar] [CrossRef]
- Krunić, M.D.; Stanisavljević, L.Z. The Biology of European Orchard Bee Osmia Cornuta (Latr.) (Hymenoptera: Megachilidae); University of Belgrade: Beograd, Serbia, 2006; ISBN 9788670780354. [Google Scholar]
- Matsumoto, S.; Abe, A.; Maejima, T. Foraging Behavior of Osmia cornifrons in an Apple Orchard. Sci. Hortic. 2009, 121, 73–79. [Google Scholar] [CrossRef]
- Osterman, J.; Theodorou, P.; Radzevičiūtė, R.; Schnitker, P.; Paxton, R.J. Apple Pollination Is Ensured by Wild Bees When Honey Bees Are Drawn Away from Orchards by a Mass Co-Flowering Crop, Oilseed Rape. Agric. Ecosyst. Environ. 2021, 315, 9. [Google Scholar] [CrossRef]
- Schrader, M.N.; LaBerge, W.E. The Nest Biology of the Bees: Andrena (Melandrena) Regularis Malloch and Andrena (Melandrena) Carlini Cockerell (Hymenoptera: Andrenidae). In Biological Notes; Illinois Natural History Survey: Urbana, IL, USA, 1978. [Google Scholar]
- Miliczky, E.R.; Mayer, D.F.; Lunden, J.D. Notes on the Nesting Biology of Andrena (Melandrena) Nivalis Smith (Hymenoptera: Andrenidae). J. Kans. Entomol. Soc. 1990, 63, 166–174. [Google Scholar]
- Miliczky, E. Observations on the Nesting Biology of Andrena (Plastandrena) Prunorum Cockerell in Washington State (Hymenoptera: Andrenidae). J. Kans. Entomol. Soc. 2008, 81, 110–121. [Google Scholar] [CrossRef]
- Bossert, S.; Wood, T.J.; Patiny, S.; Michez, D.; Almeida, E.A.B.; Minckley, R.L.; Packer, L.; Neff, J.L.; Copeland, R.S.; Straka, J.; et al. Phylogeny, Biogeography and Diversification of the Mining Bee Family Andrenidae. Syst. Entomol. 2021, 47, 283–302. [Google Scholar] [CrossRef]
- Rozen, J.G. Biology Notes on the Bee Andrena Accepta Viereck (Hymenoptera, Andrenidae). J. N. Y. Entomol. Soc. 1973, 81, 54–61. [Google Scholar]
- Johnson, M.D. Observations on the Biology of Andrena (Melandrena) Dunningi Cockerell (Hymenoptera: Andrenidae). J. Kans. Entomol. Soc. 1981, 54, 32–40. [Google Scholar]
- Larsson, M.; Franzén, M. Critical Resource Levels of Pollen for the Declining Bee Andrena Hattorfiana (Hymenoptera, Andrenidae). Biol. Conserv. 2007, 134, 405–414. [Google Scholar] [CrossRef]
- Larkin, L.L.; Neff, J.L.; Simpson, B.B. The Evolution of a Pollen Diet: Host Choice and Diet Breadth of Andrena Bees (Hymenoptera: Andrenidae). Apidologie 2008, 39, 133–145. [Google Scholar] [CrossRef]
- Neff, J.L.; Simpson, B.B. Nesting and Foraging Behavior of Andrena (Callandrena) Rudbeckiae Robertson (Hymenoptera: Apoidea: Andrenidae) in Texas. J. Kans. Entomol. Soc. 1997, 70, 100–113. [Google Scholar]
- Tang, J.; Quan, Q.-M.; Chen, J.-Z.; Wu, T.; Huang, S.-Q. Pollinator Effectiveness and Importance between Female and Male Mining Bee (Andrena). Biol. Lett. 2019, 15, 20190479. [Google Scholar] [CrossRef]
- Burger, H.; Marquardt, M.; Babucke, K.; Heuel, K.C.; Ayasse, M.; Dotterl, S.; Galizia, C.G. Neural and Behavioural Responses of the Pollen-Specialist Bee Andrena Vaga to Salix Odours. J. Exp. Biol. 2021, 224, jeb242166. [Google Scholar] [CrossRef]
- Batra, S.W.T. Polyester-Making Bees and Other Innovative Insect Chemists. J. Chem. Educ. 1985, 62, 121–124. [Google Scholar] [CrossRef]
- Almeida, E.A.B. Colletidae Nesting Biology (Hymenoptera: Apoidea)*. Apidologie 2008, 39, 16–29. [Google Scholar] [CrossRef]
- Gardner, K.E.; Ascher, J.S. Notes on the Native Bee Pollinators in New York Apple Orchards. J. N. Y. Entomol. Soc. 2006, 114, 86–91. [Google Scholar] [CrossRef]
- MacIvor, J.S. Cavity-Nest Boxes for Solitary Bees: A Century of Design and Research. Apidologie 2017, 48, 311–327. [Google Scholar] [CrossRef]
- Ferrari, R.R.; Buschini, M.L.T.; Diniz, M.E.R.; Zhu, C.-D.; Melo, G.A.R. Discovery of Mourecotelles (Hymenoptera, Apidae, Colletinae) in Brazil: Nesting Biology and Pollen Preferences of a Remarkable New Species of the Genus. J. Hymenopt. Res. 2022, 89, 211–231. [Google Scholar] [CrossRef]
- Batra, S.W.T. Solitary Bees. Sci. Am. 1984, 250, 120–127. [Google Scholar] [CrossRef]
- Danforth, B.N.; Conway, L.; Ji, S. Phylogeny of Eusocial Lasioglossum Reveals Multiple Losses of Eusociality within a Primitively Eusocial Clade of Bees (Hymenoptera: Halictidae). Syst. Biol. 2003, 52, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Stockhammer, K.A. Nesting Habits and Life Cycle of a Sweat Bee, Augochlora Pura (Hymenoptera: Halictidae). J. Kans. Entomol. Soc. 1966, 39, 157–192. [Google Scholar]
- Winter, K.; Adams, L.; Thorp, R.; Inouye, D.; Day, L.; Ascher, J.; Buchmann, S. Importation of Non-Native Bumblebees into North America: Potential Consequences of Using Bombus Terrestris and Other Non-Native Bumble Bees for Greenhouse Crop Pollination in Canada, Mexico and the United States; North American Pollinator Protection Campaign: San Francisco, CA, USA, 2006; p. 33. [Google Scholar]
- Sampson, B.J.; Danka, R.G.; Stringer, S.J. Nectar Robbery by Bees Xylocopa virginica and Apis mellifera Contributes to the Pollination of Rabbiteye Blueberry. J. Econ. Entomol. 2004, 97, 735–740. [Google Scholar] [CrossRef]
- Dalmazzo, M.; Roig-Alsina, A. Nest Structure and Notes on the Social Behavior of Augochlora amphitrite (Schrottky) (Hymenoptera, Halictidae). J. Hymenopt. Res. 2012, 26, 17–29. [Google Scholar] [CrossRef]
- Cane, J.H. Dufour’s Gland Secretion in the Cell Linings of Bees (Hymenoptera: Apoidea). J. Chem. Ecol. 1981, 7, 403–410. [Google Scholar] [CrossRef]
- Mitra, A. Function of the Dufour’s Gland in Solitary and Social Hymenoptera. J. Hymenopt. Res. 2013, 35, 33–58. [Google Scholar] [CrossRef]
- Cane, J.H.; Griswold, T.; Parker, F.D. Substrates and Materials Used for Nesting by North American Osmia Bees (Hymenoptera: Apiformes: Megachilidae). Ann. Entomol. Soc. Am. 2007, 100, 350–358. [Google Scholar] [CrossRef]
- Bosch, J.; Kemp, W.P. Effect of Wintering Duration and Temperature on Survival and Emergence Time in Males of the Orchard Pollinator Osmia lignaria (Hymenoptera: Megachilidae). Environ. Entomol. 2003, 32, 711–716. [Google Scholar] [CrossRef]
- Stanley, C. Blue Orchard Bee. In Fact Sheet Series: Insects—Beneficials; Utah State University Cooperative Extension: Logan, UT, USA, 2012; Volume ENT-162-12, p. 4. [Google Scholar]
- Torchio, P.F. In-Nest Biologies and Development of Immature Stages of Three Osmia Species (Hymenoptera: Megachilidae). Ann. Entomol. Soc. Am. 1989, 82, 599–615. [Google Scholar] [CrossRef]
- Kraemer, M.E.; Favi, F.D.; Niedziela, C.E. Nesting and Pollen Preference of Osmia Lignaria Lignaria (Hymenoptera: Megachilidae) in Virginia and North Carolina Orchards. Environ. Entomol. 2014, 43, 932–941. [Google Scholar] [CrossRef] [PubMed]
- Bosch, J.; Kemp, W.P. Developing and Establishing Bee Species as Crop Pollinators: The Example of Osmia Spp. (Hymenoptera: Megachilidae) and Fruit Trees. Bull. Entomol. Res. 2002, 92, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Wolf, A.T.; Ascher, J.S. Bees of Wisconsin (Hymenoptera: Apoidea: Anthophila). Great Lakes Entomol. 2009, 41, 129–168. [Google Scholar]
- Lerman, S.B.; Milam, J. Bee Fauna and Floral Abundance within Lawn-Dominated Suburban Yards in Springfield, MA. Ann. Entomol. Soc. Am. 2016, 109, 713–723. [Google Scholar] [CrossRef]
- Wood, T.J.; Roberts, S.P.M. An Assessment of Historical and Contemporary Diet Breadth in Polylectic Andrena Bee Species. Biol. Conserv. 2017, 215, 72–80. [Google Scholar] [CrossRef]
- Dar, S.A.; Sofi, M.A.; El-Sharnouby, M.; Hassan, M.; Rashid, R.; Mir, S.H.; Naggar, Y.A.; Salah, M.; Gajger, I.T.; Sayed, S. Nesting Behaviour and Foraging Characteristics of Andrena Cineraria (Hymenoptera: Andrenidae). Saudi. J. Biol. Sci. 2021, 28, 4147–4154. [Google Scholar] [CrossRef]
- Scott, Z.; Ginsberg, H.S.; Alm, S.R. Native Bee Diversity and Pollen Foraging Specificity in Cultivated Highbush Blueberry (Ericaceae: Vaccinium corymbosum) in Rhode Island. Environ. Entomol. 2016, 45, 1432–1438. [Google Scholar] [CrossRef]
- Weaver, S.A.; Mallinger, R.E. A Specialist Bee and Its Host Plants Experience Phenological Shifts at Different Rates in Response to Climate Change. Ecology 2022, 103, e3658. [Google Scholar] [CrossRef]
- Zurbuchen, A.; Landert, L.; Klaiber, J.; Müller, A.; Hein, S.; Dorn, S. Maximum Foraging Ranges in Solitary Bees: Only Few Individuals Have the Capability to Cover Long Foraging Distances. Biol. Conserv. 2010, 143, 669–676. [Google Scholar] [CrossRef]
- Fowler, J. Specialist Bees of the Northeast: Host Plants and Habitat Conservation. Northeast. Nat. (Steuben) 2016, 23, 305–320. [Google Scholar] [CrossRef]
- Haider, M.; Dorn, S.; Sedivy, C.; Müller, A. Phylogeny and Floral Hosts of a Predominantly Pollen Generalist Group of Mason Bees (Megachilidae: Osmiini). Biol. J. Linnean Soc. 2014, 111, 78–91. [Google Scholar] [CrossRef]
- Müller, A. Osmia (Orientosmia) Maxschwarzi Sp. n., a New Palaearctic Osmiine Bee with Extraordinarily Long Mouthparts (Hymenoptera, Apiformes, Megachilidae). Mitt. Schweiz. Entomol. Ges. 2012, 85, 27–35. [Google Scholar]
- Bastin, F.; Savarit, F.; Lafon, G.; Sandoz, J.C. Age-Specific Olfactory Attraction between Western Honey Bee Drones (Apis Mellifera) and Its Chemical Basis. PLoS ONE 2017, 12, e0185949. [Google Scholar] [CrossRef]
- Seeley, T.D.; Mikheyev, A.S. Reproductive Decisions by Honey Bee Colonies: Tuning Investment in Male Production in Relation to Success in Energy Acquisition. Insectes Sociaux 2003, 50, 134–138. [Google Scholar] [CrossRef]
- Seeley, T.D.; Morse, R.A. The Nest of the Honey Bee (Apis mellifera L.). Insectes Sociaux 1976, 23, 495–512. [Google Scholar] [CrossRef]
- Seeley, T.D.; Morse, R.A. Nest Site Selection by the Honey Bee, Apis mellifera. Insectes Sociaux 1978, 25, 323–337. [Google Scholar] [CrossRef]
- Remolina, S.C.; Hughes, K.A. Evolution and Mechanisms of Long Life and High Fertility in Queen Honey Bees. Age 2008, 30, 185. [Google Scholar] [CrossRef]
- Del Castillo, R.C.; Sanabria-Urbán, S.; Serrano-Meneses, M.A. Trade-offs in the Evolution of Bumblebee Colony and Body Size: A Comparative Analysis. Ecol. Evol. 2015, 5, 3926. [Google Scholar] [CrossRef]
- Williams, N.M.; Mola, J.M.; Stuligross, C.; Harrison, T.; Page, M.L.; Brennan, R.M.; Rosenberger, N.M.; Rundlöf, M. Fantastic Bees and Where to Find Them: Locating the Cryptic Overwintering Queens of a Western Bumble Bee. Ecosphere 2019, 10, e02949. [Google Scholar] [CrossRef]
- Kells, A.R.; Goulson, D. Preferred Nesting Sites of Bumblebee Queens (Hymenoptera: Apidae) in Agroecosystems in the UK. Biol. Conserv. 2003, 109, 165–174. [Google Scholar] [CrossRef]
- Goulson, D. Bumble Bees: Their Behaviour and Ecology; Oxford University Press: Oxford, UK, 2003; ISBN 9780198526063. [Google Scholar]
- Page, M.L.; Nicholson, C.C.; Brennan, R.M.; Britzman, A.T.; Greer, J.; Hemberger, J.; Kahl, H.; Müller, U.; Peng, Y.; Rosenberger, N.M.; et al. A Meta-Analysis of Single Visit Pollination Effectiveness Comparing Honeybees and Other Floral Visitors. Am. J. Bot. 2021, 108, 2196–2207. [Google Scholar] [CrossRef] [PubMed]
- Michener, C.D. The Bees of the World, 2nd ed.; Johns Hopkins University Press: Baltimore, MD, USA, 2007; ISBN 9780801885730. [Google Scholar]
- Portman, Z.M.; Tepedino, V.J. Convergent Evolution of Pollen Transport Mode in Two Distantly Related Bee Genera (Hymenoptera: Andrenidae and Melittidae). Apidologie 2017, 48, 461–472. [Google Scholar] [CrossRef]
- Konzmann, S.; Hilgendorf, F.; Niester, C.; Rech, A.R.; Lunau, K. Morphological Specialization of Heterantherous Rhynchanthera grandiflora (Melastomataceae) Accommodates Pollinator Diversity. Plant Biol. 2020, 22, 583–590. [Google Scholar] [CrossRef]
- Miller, A.E.; Brosi, B.J.; Magnacca, K.; Daily, G.C.; Pejchar, L. Pollen Carried by Native and Nonnative Bees in the Large-Scale Reforestation of Pastureland in Hawai‘i: Implications for Pollination. Pac. Sci. 2015, 69, 67–79. [Google Scholar] [CrossRef]
- Ogilvie, J.E.; Forrest, J.R.K. Interactions between Bee Foraging and Floral Resource Phenology Shape Bee Populations and Communities. Curr. Opin. Insect Sci. 2017, 21, 75–82. [Google Scholar] [CrossRef]
- Schlindwein, C.; Pick, R.A.; Martins, C.F. Evaluation of Oligolecty in the Brazilian Bee Ptilothrix plumata (Hymenoptera, Apidae, Emphorini). Apidologie 2009, 40, 106–116. [Google Scholar] [CrossRef][Green Version]
- Bosch, J.; Kemp, W.P. How to Manage the Blue Orchard Bee as an Orchard Pollinator. In Sustainable Agricultural Network handbook series; Sustainable Agricultural Network: Beltsville, MD, USA, 2001; ISBN 1888626062. [Google Scholar]
- Simons, M.A.; Smith, A.R. Ovary Activation Does Not Correlate with Pollen and Nectar Foraging Specialization in the Bumblebee Bombus impatiens. PeerJ 2018, 2018, e4415. [Google Scholar] [CrossRef]
- Saunders, M.E. Insect Pollinators Collect Pollen from Wind-Pollinated Plants: Implications for Pollination Ecology and Sustainable Agriculture. Insect Conserv. Divers. 2018, 11, 13–31. [Google Scholar] [CrossRef]
- Kratochwil, A.; Beil, M.; Schwabe, A. Complex Structure of Pollinator-Plant Interaction-Webs: Random, Nested, with Gradients or Modules? Apidologie 2009, 40, 634–650. [Google Scholar] [CrossRef]
- Nooten, S.S.; Odanaka, K.; Rehan, S.M. Characterization of Wild Bee Communities in Apple and Blueberry Orchards. Agric For Entomol 2020, 22, 157–168. [Google Scholar] [CrossRef]
- Graham, J.R.; Willcox, E.; Ellis, J.D. The Potential Management of a Ground-Nesting, Solitary Bee: Anthophora Abrupta (Hymenoptera: Apidae). Fla. Entomol. 2015, 98, 528–535. [Google Scholar] [CrossRef]
- Pyke, G.H.; Thomson, J.D.; Inouye, D.W.; Miller, T.J. Effects of Climate Change on Phenologies and Distributions of Bumble Bees and the Plants They Visit. Ecosphere 2016, 7, e01267. [Google Scholar] [CrossRef]
- Moisan-Deserres, J.; Girard, M.; Chagnon, M.; Fournier, V. Pollen Loads and Specificity of Native Pollinators of Lowbush Blueberry. J. Econ. Entomol. 2014, 107, 1156–1162. [Google Scholar] [CrossRef]
- Averill, A.L.; Sylvia, M.M.; Hahn, N.; Couto, A.V. Bees (Hymenoptera: Apoidea) Foraging on American Cranberry in Massachusetts. Northeast. Nat. 2018, 25, 502–512. [Google Scholar] [CrossRef]
- Conrad, T.; Stöcker, C.; Ayasse, M. The Effect of Temperature on Male Mating Signals and Female Choice in the Red Mason Bee, Osmia bicornis (L.). Ecol. Evol. 2017, 7, 8966–8975. [Google Scholar] [CrossRef]
- Conrad, T.; Ayasse, M. The Role of Vibrations in Population Divergence in the Red Mason Bee, Osmia bicornis. Curr. Biol. 2015, 25, 2819–2822. [Google Scholar] [CrossRef]
- Seidelmann, K.; Rolke, D. Advertisement of Unreceptivity—Perfume Modifications of Mason Bee Females (Osmia bicornis and O. Cornuta) and a Non-Existing Antiaphrodisiac. PLoS ONE 2019, 14, e0215925. [Google Scholar] [CrossRef]
- Reyes, M.; Crauser, D.; Prado, A.; le Conte, Y. Flight Activity of Honey Bee (Apis mellifera) Drones. Apidologie 2019, 50, 669–680. [Google Scholar] [CrossRef]
- Short, C.A.; Lucky, A. Pure Gold-Green Sweat Bee Augochlora Pura (Say 1837) (Insecta: Hymenoptera: Halictidae: Halictinae); University of Florida: Gainesville, FL, USA, 2018. [Google Scholar]
- Buckley, P.R. The Honey Bee (Apis mellifera): Cost Benefit Ratio of the Honey Bee; Lincoln University: Canterbury, New Zealand, 2003. [Google Scholar]
- Zhang, G.; st. Clair, A.L.; Dolezal, A.G.; Toth, A.L.; O’Neal, M.E. Can Native Plants Mitigate Climate-Related Forage Dearth for Honey Bees (Hymenoptera: Apidae)? J. Econ. Entomol. 2022, 115, 1–9. [Google Scholar] [CrossRef]
- Winston, M.L. The Biology of the Honey Bee, 1st ed.; Harvard University Press: Cambridge, MA, USA, 1987; ISBN 9780674074088. [Google Scholar]
- Khan, K.; Ghramh, H.A. Nutritional Efficacy of Different Diets Supplemented with Microalga Arthrospira platensis (Spirulina) in Honey Bees (Apis mellifera). J. King Saud Univ. Sci. 2022, 34, 101819. [Google Scholar] [CrossRef]
- Greenleaf, S.S.; Williams, N.M.; Winfree, R.; Kremen, C. Bee Foraging Ranges and Their Relationship to Body Size. Oecologia 2007, 153, 589–596. [Google Scholar] [CrossRef]
- Wolf, S.; Moritz, R.F.A. Foraging Distance in Bombus terrestris L. (Hymenoptera: Apidae). Apidologie 2008, 39, 419–427. [Google Scholar] [CrossRef]
- Couvillon, M.J.; Schürch, R.; Ratnieks, F.L.W. Dancing Bees Communicate a Foraging Preference for Rural Lands in High-Level Agri-Environment Schemes. Curr. Biol. 2014, 24, 1212–1215. [Google Scholar] [CrossRef] [PubMed]
- Wright, I.R.; Roberts, S.P.M.; Collins, B.E. Evidence of Forage Distance Limitations for Small Bees (Hymenoptera: Apidae). Eur. J. Entomol. 2015, 112, 303–310. [Google Scholar] [CrossRef]
- Osborne, J.L.; Martin, A.P.; Carreck, N.L.; Swain, J.L.; Knight, M.E.; Goulson, D.; Hale, R.J.; Sanderson, R.A. Bumblebee Flight Distances in Relation to the Forage Landscape. J. Anim. Ecol. 2008, 77, 406–415. [Google Scholar] [CrossRef] [PubMed]
- Walther-Hellwig, K.; Frankl, R. Foraging Habitats and Foraging Distances of Bumblebees, Bombus spp. (Hym., Apidae), in an Agricultural Landscape. J. Appl. Entomol. 2000, 124, 299–306. [Google Scholar] [CrossRef]
- Carreck, N.; Williams, I.H.; Oakley, J. Enhancing Farmland for Insect Pollinators Using Flower Mixtures. Asp. Appl. Biol. 1999, 54, 101–108. [Google Scholar]
- Darvill, B.; Knight, M.E.; Goulson, D. Use of Genetic Markers to Quantify Bumblebee Foraging Range and Nest Density. Oikos 2004, 107, 471–478. [Google Scholar] [CrossRef]
- Knight, M.E.; Martin, A.P.; Bishop, S.; Osborne, J.L.; Hale, R.J.; Sanderson, R.A.; Goulson, D. An Interspecific Comparison of Foraging Range and Nest Density of Four Bumblebee (Bombus) Species. Mol. Ecol. 2005, 14, 1811–1820. [Google Scholar] [CrossRef]
- Kraus, F.B.; Wolf, S.; Moritz, R.F.A. Male Flight Distance and Population Substructure in the Bumblebee Bombus terrestris. J. Anim. Ecol. 2009, 78, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Rust, R.W. Spatial and Temporal Heterogeneity of Pollen Foraging in Osmia Lignaria Propinqua (Hymenoptera: Megachilidae). Environ. Entomol. 1990, 19, 332–338. [Google Scholar] [CrossRef]
- Geib, J.C.; Strange, J.P.; Galen, A. Bumble Bee Nest Abundance, Foraging Distance, and Host-Plant Reproduction: Implications for Management and Conservation. Ecol. Appl. 2015, 25, 768–778. [Google Scholar] [CrossRef] [PubMed]
- Vinson, S.B.; Frankie, G.W.; Williams, H.J. Nest Liquid Resources of Several Cavity Nesting Bees in the Genus Centris and the Identification of a Preservative, Levulinic Acid. J. Chem. Ecol. 2006, 32, 2013–2021. [Google Scholar] [CrossRef] [PubMed]
- Eckhardt, M.; Haider, M.; Dorn, S.; Müller, A. Pollen Mixing in Pollen Generalist Solitary Bees: A Possible Strategy to Complement or Mitigate Unfavourable Pollen Properties? J. Anim. Ecol. 2014, 83, 588–597. [Google Scholar] [CrossRef]
- Heller, S.; Joshi, N.K.; Leslie, T.; Rajotte, E.G.; Biddinger, D.J. Diversified Floral Resource Plantings Support Bee Communities after Apple Bloom in Commercial Orchards. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef]
- Joshi, N.K.; Naithani, K.; Biddinger, D.J. Nest Modification Protects Immature Stages of the Japanese Orchard Bee (Osmia cornifrons) from Invasion of a Cleptoparasitic Mite Pest. Insects 2020, 11, 65. [Google Scholar] [CrossRef]
- Joshi, N.K.; Leslie, T.; Rajotte, E.G.; Kammerer, M.A.; Otieno, M.; Biddinger, D.J. Comparative Trapping Efficiency to Characterize Bee Abundance, Diversity, and Community Composition in Apple Orchards. Ann. Entomol. Soc. Am. 2015, 108, 785–799. [Google Scholar] [CrossRef]
- Quinet, M.; Warzée, M.; Vanderplanck, M.; Michez, D.; Lognay, G.; Jacquemart, A.L. Do Floral Resources Influence Pollination Rates and Subsequent Fruit Set in Pear (Pyrus Communis L.) and Apple (Malus x Domestica Borkh) Cultivars? Eur. J. Agron. 2016, 77, 59–69. [Google Scholar] [CrossRef]
- Zhu, Y.; Yang, G.; Yang, H.; Guo, L.; Xu, B.; Li, Z.; Han, S.; Zhu, X.; Li, Z.; Jones, G. Forecasting Regional Apple First Flowering Using the Sequential Model and Gridded Meteorological Data with Spatially Optimized Calibration. Comput. Electron. Agric. 2022, 196, 106869. [Google Scholar] [CrossRef]
- Stemkovski, M.; Pearse, W.D.; Griffin, S.R.; Pardee, G.L.; Gibbs, J.; Griswold, T.; Neff, J.L.; Oram, R.; Rightmyer, M.G.; Sheffield, C.S.; et al. Bee Phenology Is Predicted by Climatic Variation and Functional Traits. Ecol. Lett. 2020, 23, 1589–1598. [Google Scholar] [CrossRef] [PubMed]
- Joshi, N.K.; Otieno, M.; Rajotte, E.G.; Fleischer, S.J.; Biddinger, D.J. Proximity to Woodland and Landscape Structure Drives Pollinator Visitation in Apple Orchard Ecosystem. Front Ecol. Evol. 2016, 4, 38. [Google Scholar] [CrossRef]
- Allen-Perkins, A.; Magrach, A.; Dainese, M.; Garibaldi, L.A.; Kleijn, D.; Rader, R.; Reilly, J.R.; Winfree, R.; Lundin, O.; McGrady, C.M.; et al. CropPol: A Dynamic, Open and Global Database on Crop Pollination. Ecology 2022, 103, e3614. [Google Scholar] [CrossRef]
- Tangtorwongsakul, P.; Warrit, N.; Gale, G.A. Effects of Landscape Cover and Local Habitat Characteristics on Visiting Bees in Tropical Orchards. Agric. For. Entomol. 2018, 20, 28–40. [Google Scholar] [CrossRef]
- Steffan-Dewenter, I.; Leschke, K. Effects of Habitat Management on Vegetation and Above-Ground Nesting Bees and Wasps of Orchard Meadows in Central Europe. Biodivers. Conserv. 2003, 12, 1953–1968. [Google Scholar] [CrossRef]
- Goulson, D.; Nicholls, E.; Botías, C.; Rotheray, E.L. Bee Declines Driven by Combined Stress from Parasites, Pesticides, and Lack of Flowers. Science 2015, 347, 1255957. [Google Scholar] [CrossRef]
- Kammerer, M.A.; Biddinger, D.J.; Rajotte, E.G.; Mortensen, D.A. Local Plant Diversity across Multiple Habitats Supports a Diverse Wild Bee Community in Pennsylvania Apple Orchards. Environ. Entomol. 2016, 45, 32–38. [Google Scholar] [CrossRef]
- Evans, L.J.; Jesson, L.; Read, S.F.J.; Jochym, M.; Cutting, B.T.; Gayrard, T.; Jammes, M.A.S.; Roumier, R.; Howlett, B.G. Key Factors Influencing Forager Distribution across Macadamia Orchards Differ among Species of Managed Bees. Basic Appl. Ecol. 2021, 53, 74–85. [Google Scholar] [CrossRef]
- Sheffield, C.S.; Kevan, P.G.; Pindar, A.; Packer, L. Bee (Hymenoptera: Apoidea) Diversity within Apple Orchards and Old Fields in the Annapolis Valley, Nova Scotia, Canada. Can. Entomol. 2013, 145, 94–114. [Google Scholar] [CrossRef]
- Sheffield, C.S.; Kevan, P.G.; Westby, S.M.; Smith, R.F. Diversity of Cavity-Nesting Bees (Hymenoptera: Apoidea) within Apple Orchards and Wild Habitats in the Annapolis Valley, Nova Scotia, Canada. Can. Entomol. 2008, 140, 235–249. [Google Scholar] [CrossRef]
- Russo, L.; Danforth, B. Pollen Preferences among the Bee Species Visiting Apple (Malus Pumila) in New York. Apidologie 2017, 48, 806–820. [Google Scholar] [CrossRef]
- Tepedino, V.J.; Alston, D.G.; Bradley, B.A.; Toler, T.R.; Griswold, T.L. Orchard Pollination in Capitol Reef National Park, Utah, USA. Honey Bees or Native Bees? Biodivers. Conserv. 2007, 16, 3083–3094. [Google Scholar] [CrossRef]
- Thakur, M. Bees as Pollinators—Biodiversity and Conservation. Int. Res. J. Agric. Sci. Soil Sci. 2012, 2, 1–7. [Google Scholar]
- Kammerer, M.A.; Biddinger, D.J.; Joshi, N.K.; Rajotte, E.G.; Mortensen, D.A. Modeling Local Spatial Patterns of Wild Bee Diversity in Pennsylvania Apple Orchards. Landsc. Ecol. 2016, 31, 2459–2469. [Google Scholar] [CrossRef]
- Endler, J.A. Natural Selection in the Wild (MPB-21); Princeton University Press: Princeton, NJ, USA, 1986; Volume 21, ISBN 978-0-691-20951-7. [Google Scholar]
- Futuyma, D.J. Evolutionary Biology, 1st ed.; Sinauer Associates: Sunderland, MA, USA, 1986; ISBN 9780878931880. [Google Scholar]
- Wright, S. Evolution and the Genetics of Populations (Variability within and among Natural Populations), 1st ed.; University of Chicago Press: Chicago, IL, USA, 1984; Volume 4, ISBN 9780226910413. [Google Scholar]
- Avise, J.C. Molecular Markers, Natural History and Evolution, 3rd ed.; Springer: New York, NY, USA, 2012; Volume 3, ISBN 9781461523819. [Google Scholar]
- Kent, C.F.; Dey, A.; Patel, H.; Tsvetkov, N.; Tiwari, T.; MacPhail, V.J.; Gobeil, Y.; Harpur, B.A.; Gurtowski, J.; Schatz, M.C.; et al. Conservation Genomics of the Declining North American Bumblebee Bombus Terricola Reveals Inbreeding and Selection on Immune Genes. Front. Genet. 2018, 9, 316. [Google Scholar] [CrossRef]
- Cameron, S.A.; Hines, H.M.; Williams, P.H. A Comprehensive Phylogeny of the Bumble Bees (Bombus). Biol. J. Linn. Soc. 2007, 91, 161–188. [Google Scholar] [CrossRef]
- Cordes, N.; Huang, W.F.; Strange, J.P.; Cameron, S.A.; Griswold, T.L.; Lozier, J.D.; Solter, L.F. Interspecific Geographic Distribution and Variation of the Pathogens Nosema Bombi and Crithidia Species in United States Bumble Bee Populations. J. Invertebr. Pathol. 2012, 109, 209–216. [Google Scholar] [CrossRef]
- Koch, J.B.; Looney, C.; Sheppard, W.S.; Strange, J.P. Patterns of Population Genetic Structure and Diversity across Bumble Bee Communities in the Pacific Northwest. Conserv. Genet. 2017, 18, 507–520. [Google Scholar] [CrossRef]
- Ascher, J.S.; Pickering, J. Bee Species Guide and World Checklist (Hymenoptera: Apidae: Anthophila). Available online: http://www.discoverlife.org/mp/20q?guide=Apoidea_species (accessed on 21 September 2022).
- Exeler, N.; Kratochwil, A.; Hochkirch, A. Does Recent Habitat Fragmentation Affect the Population Genetics of a Heathland Specialist, Andrena Fuscipes (Hymenoptera: Andrenidae)? Conserv. Genet. 2010, 11, 1679–1687. [Google Scholar] [CrossRef]
- Ballare, K.M.; Jha, S. Genetic Structure across Urban and Agricultural Landscapes Reveals Evidence of Resource Specialization and Philopatry in the Eastern Carpenter Bee, Xylocopa virginica L. Evol. Appl. 2021, 14, 136–149. [Google Scholar] [CrossRef] [PubMed]
- Kelemen, E.P.; Rehan, S.M. Conservation Insights from Wild Bee Genetic Studies: Geographic Differences, Susceptibility to Inbreeding, and Signs of Local Adaptation. Evol. Appl. 2021, 14, 1485–1496. [Google Scholar] [CrossRef] [PubMed]
- López-Uribe, M.M.; Morreale, S.J.; Santiago, C.K.; Danforth, B.N. Nest Suitability, Fine-Scale Population Structure and Male-Mediated Dispersal of a Solitary Ground Nesting Bee in an Urban Landscape. PLoS ONE 2015, 10, e0125719. [Google Scholar] [CrossRef]
- Lozier, J.D.; Strange, J.P.; Koch, J.B. Landscape Heterogeneity Predicts Gene Flow in a Widespread Polymorphic Bumble Bee, Bombus Bifarius (Hymenoptera: Apidae). Conserv. Genet. 2013, 14, 1099–1110. [Google Scholar] [CrossRef]
- Ellis, J.S.; Knight, M.E.; Darvill, B.; Goulson, D. Extremely Low Effective Population Sizes, Genetic Structuring and Reduced Genetic Diversity in a Threatened Bumblebee Species, Bombus sylvarum (Hymenoptera: Apidae). Mol. Ecol. 2006, 15, 4375–4386. [Google Scholar] [CrossRef]
- Krombein, K.V.; Hurd, P.D., Jr.; Smith, D.R.; Burks, B.D. Catalog of Hymenoptera in America North of Mexico; Smithsonian Institution Press: Washington, DC, USA, 1979; Volume 1. [Google Scholar]
- Scott, V.L.; Ascher, J.S.; Griswold, T.; Nufio, C. The Bees of Colorado (Hymenoptera: Apoidea: Anthophila); University of Colorado Museum of Natural History: Boulder, CO, USA, 2011; Volume 23. [Google Scholar]
- Cane, J.H. Specialist Osmia Bees Forage Indiscriminately among Hybridizing Balsamorhiza Floral Hosts. Oecologia 2011, 167, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Torchio, P.F. Osmia Ribifloris, a Native Bee Species Developed as a Commercially Managed Pollinator of Highbush Blueberry (Hymenoptera: Megachilidae). J. Kans. Entomol. Soc. 1990, 63, 427–436. [Google Scholar]
- Sampson, B.J.; Stringer, S.J.; Cane, J.H.; Spiers, J.M. Screenhouse Evaluations of a Mason Bee Osmia ribifloris (Hymenoptera: Megachilidae) as a Pollinator for Blueberries in the Southeastern United States. Small Fruits Rev. 2004, 3, 381–392. [Google Scholar] [CrossRef]
- Biddinger, D.J.; Rajotte, E.G. Integrated Pest and Pollinator Management—Adding a New Dimension to an Accepted Paradigm. Curr. Opin. Insect Sci. 2015, 10, 204–209. [Google Scholar] [CrossRef]
- Bosch, J.; Blas, M. Effect of Over-Wintering and Incubation Temperatures on Adult Emergence in Osmia Cornuta Latr (Hymenoptera, Megachilidae). Apidologie 1994, 25, 265–277. [Google Scholar] [CrossRef]
- LeCroy, K.A.; Savoy-Burke, G.; Carr, D.E.; Delaney, D.A.; Roulston, T.H. Decline of Six Native Mason Bee Species Following the Arrival of an Exotic Congener. Sci. Rep. 2020, 10, 1–9. [Google Scholar] [CrossRef]
- Young, B.E.; Schweitzer, D.F.; Sears, N.A.; Ormes, M.F. Conservation and Management of North American Mason Bees; NatureServe: Arlington, VA, USA, 2015. [Google Scholar]
- Beadle, K.; Singh, K.S.; Troczka, B.J.; Randall, E.; Zaworra, M.; Zimmer, C.T.; Hayward, A.; Reid, R.; Kor, L.; Kohler, M.; et al. Genomic Insights into Neonicotinoid Sensitivity in the Solitary Bee Osmia bicornis. PLoS Genet. 2019, 15, e1007903. [Google Scholar] [CrossRef] [PubMed]
- Möllmann, J.S.; Colgan, T.J. Genomic Architecture and Sexually Dimorphic Expression Underlying Immunity in the Red Mason Bee, Osmia bicornis. Insect Mol. Biol. 2022, 31, 686–700. [Google Scholar] [CrossRef]
- Kim, H.-Y.; Lee, H.-Y.; Lee, S.-B.; Kim, S.-R.; Hong, M.-Y.; Kim, D.-Y.; Kim, I.-S. Mitochondrial DNA Sequence Variation of the Mason Bee, Osmia Cornifrons (Hymenoptera: Apidae). Int. J. Indust. Entomol. 2008, 16, 75–86. [Google Scholar]
- Fisher, K.; Caceres, C. A Filter Rearing System for Mass Reared Genetic Sexing Strains of Mediterranean Fruit Fly (Diptera: Tephritidae). In Proceedings of the Joint Proceedings of the International Conference on Area-Wide Control of Insect Pests and the 5th International Symposium on Fruit Flies of Economic Importance; Kenghong, T., Ed.; Penerbit Universiti Sains Malaysia: Penang, Malaysia, 2000; pp. 543–550. [Google Scholar]
- Klein, A.-M.; Vaissière, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of Pollinators in Changing Landscapes for World Crops. Proc. R. Soc. B Biol. Sci. 2007, 274, 303–313. [Google Scholar] [CrossRef]
- Hill-Burns, E.M.; Clark, A.G. X-Linked Variation in Immune Response in Drosophila melanogaster. Genetics 2009, 183, 1491. [Google Scholar] [CrossRef] [PubMed]
- Abraham, J.; Benhotons, G.S.; Krampah, I.; Tagba, J.; Amissah, C.; Abraham, J.D. Commercially Formulated Glyphosate Can Kill Non-Target Pollinator Bees under Laboratory Conditions. Entomol. Exp. Appl. 2018, 166, 695–702. [Google Scholar] [CrossRef]
- Bailey, J.; Scott-Dufree, C.; Harris, R.; Tolman, J.; Harris, B. Contact and Oral Toxicity to Honey Bees (Apis mellifera) of Agents Registered for Use for Sweet Corn Insect Control in Ontario, Canada. Apidologie 2005, 36, 623–633. [Google Scholar] [CrossRef]
- Azpiazu, C.; Bosch, J.; Viñuela, E.; Medrzycki, P.; Teper, D.; Sgolastra, F. Chronic Oral Exposure to Field- Realistic Pesticide Combinations via Pollen and Nectar: Effects on Feeding and Thermal Performance in a Solitary Bee. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef]
- Bromilow, R.H.; Chamberlain, K.; Evans, A.A. Physicochemical Aspects of Phloem Translocation of Herbicides. Weed Sci. 1990, 38, 305–314. [Google Scholar] [CrossRef]
- Joshi, N.K.; Biddinger, D.J.; Rajotte, E.G. A Survey of Apple Pollination Practices, Knowledge and Attitudes of Fruit Growers in Pennsylvania. In Proceedings of the 10th International Pollination Symposium, Puebla, Mexico, 27 June 2011. [Google Scholar]
- Biddinger, D.J.; Rajotte, E.G.; Joshi, N.K. Integrating Pollinator Health into Tree Fruit IPM: A Case Study of Pennsylvania Apple Production. In The Pollination of Cultivated Plants: A Compendium for Practitioners; Roubik, D.W., Ed.; Food and Agriculture Organization of The United Nations: Rome, Italy, 2018; pp. 67–80. ISBN 9789251305065. [Google Scholar]
- Hull, L.A.; Joshi, N.K.; Zaman, F.U. Management of Internal Feeding Lepidopteran Pests in Apple. Arthropod Manag. Tests 2009, 34, A8. [Google Scholar] [CrossRef]
- Heller, S.; Joshi, N.K.; Chen, J.; Rajotte, E.G.; Mullin, C.; Biddinger, D.J. Pollinator Exposure to Systemic Insecticides and Fungicides Applied in the Previous Fall and Pre-Bloom Period in Apple Orchards. Environ. Pollut. 2020, 265, 114589. [Google Scholar] [CrossRef]
- Heller, S.; Fine, J.; Phan, N.T.; Rajotte, E.G.; Biddinger, D.J.; Joshi, N.K. Toxicity of Formulated Systemic Insecticides Used in Apple Orchard Pest Management Programs to the Honey Bee (Apis mellifera (L.)). Environments 2022, 9, 90. [Google Scholar] [CrossRef]
- Biddinger, D.J.; Robertson, J.L.; Mullin, C.A.; Frazier, J.L.; Ashcraft, S.A.; Rajotte, E.G.; Joshi, N.K.; Vaughan, M. Comparative Toxicities and Synergism of Apple Orchard Pesticides to Apis mellifera (L.) and Osmia Cornifrons (Radoszkowski). PLoS ONE 2013, 8, e72587. [Google Scholar] [CrossRef] [PubMed]
- Krupke, C.H.; Hunt, G.J.; Eitzer, B.D.; Andino, G.; Given, K. Multiple Routes of Pesticide Exposure for Honey Bees Living near Agricultural Fields. PLoS ONE 2012, 7, e29268. [Google Scholar] [CrossRef]
- Cane, J.H. Soils of Ground-Nesting Bees (Hymenoptera: Apoidea): Texture, Moisture, Cell Depth and Climate. J. Kans. Entomol. Soc. 1991, 64, 406–413. [Google Scholar]
- Schaafsma, A.; Limay-Rios, V.; Baute, T.; Smith, J.; Xue, Y. Neonicotinoid Insecticide Residues in Surface Water and Soil Associated with Commercial Maize (Corn) Fields in Southwestern Ontario. PLoS ONE 2015, 10, e0118139. [Google Scholar] [CrossRef] [PubMed]
- Samson-Robert, O.; Labrie, G.; Chagnon, M.; Fournier, V. Neonicotinoid-Contaminated Puddles of Water Represent a Risk of Intoxication for Honey Bees. PLoS ONE 2014, 9, e108443. [Google Scholar] [CrossRef] [PubMed]
- Hladik, M.L.; Vandever, M.; Smalling, K.L. Exposure of Native Bees Foraging in an Agricultural Landscape to Current-Use Pesticides. Sci. Total Environ. 2016, 542, 469–477. [Google Scholar] [CrossRef]
- IRAC International MoA Working Group. IRAC Mode of Action Classification Scheme; 10.3; CropLife International: Brussels, Belgium, 2022. [Google Scholar]
- Singh, A.; Sah, L.P.; Dhoj, G.C.Y.; Devkota, M.; Colavito, L.A.; Rajbhandari, B.P.; Norton, G.; Rajotte, E.G.; Muniappan, R. Evaluation of Pest Exclusion Net to Major Insect Pest of Tomato in Kavre and Lalitpur. Nepal. J. Agric. Sci. 2018, 16, 128–137. [Google Scholar]
- Belsky, J.; Biddinger, D.J.; Joshi, N.K. Whole-Body Acute Contact Toxicity of Formulated Insecticide Mixtures to Blue Orchard Bees (Osmia lignaria). Toxics 2021, 9, 61. [Google Scholar] [CrossRef] [PubMed]
- Hayward, A.; Beadle, K.; Singh, K.S.; Exeler, N.; Zaworra, M.; Almanza, M.-T.; Nikolakis, A.; Garside, C.; Glaubitz, J.; Bass, C.; et al. The Leafcutter Bee, Megachile Rotundata, Is More Sensitive to N-Cyanoamidine Neonicotinoid and Butenolide Insecticides than Other Managed Bees. Nat. Ecol. Evol. 2019, 3, 1521–1524. [Google Scholar] [CrossRef]
- Tosi, S.; Nieh, J.C. A Common Neonicotinoid Pesticide, Thiamethoxam, Alters Honey Bee Activity, Motor Functions, and Movement to Light. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef]
- Claus, G.; Pisman, M.; Spanoghe, P.; Smagghe, G.; Eeraerts, M. Larval Oral Exposure to Thiacloprid: Dose-Response Toxicity Testing in Solitary Bees, Osmia Spp. (Hymenoptera: Megachilidae). Ecotoxicol. Environ. Saf. 2021, 215, 112143. [Google Scholar] [CrossRef] [PubMed]
- Elston, C.; Thompson, H.M.; Walters, K.F.A. Sub-Lethal Effects of Thiamethoxam, a Neonicotinoid Pesticide, and Propiconazole, a DMI Fungicide, on Colony Initiation in Bumblebee (Bombus terrestris) Micro-Colonies. Apidologie 2013, 44, 563–574. [Google Scholar] [CrossRef]
- Phan, N.T.; Joshi, N.K.; Rajotte, E.G.; Zhu, F.; Peter, K.A.; López-Uribe, M.M.; Biddinger, D.J. Systemic Pesticides in a Solitary Bee Pollen Food Store Affect Larval Development and Increase Pupal Mortality. bioRxiv 2022. [Google Scholar] [CrossRef]
- Brandt, A.; Hohnheiser, B.; Sgolastra, F.; Bosch, J.; Meixner, M.D.; Büchler, R. Immunosuppression Response to the Neonicotinoid Insecticide Thiacloprid in Females and Males of the Red Mason Bee Osmia bicornis L. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef]
- Di Prisco, G.; Cavaliere, V.; Annoscia, D.; Varricchio, P.; Caprio, E.; Nazzi, F.; Gargiulo, G.; Pennacchio, F. Neonicotinoid Clothianidin Adversely Affects Insect Immunity and Promotes Replication of a Viral Pathogen in Honey Bees. Proc. Natl. Acad. Sci. USA 2013, 110, 18466–18471. [Google Scholar] [CrossRef] [PubMed]
- Fauser-Misslin, A.; Sadd, B.M.; Neumann, P.; Sandrock, C. Influence of Combined Pesticide and Parasite Exposure on Bumblebee Colony Traits in the Laboratory. J. Appl. Ecol. 2014, 51, 450–459. [Google Scholar] [CrossRef]
- Tsvetkov, N.; Samson-Robert, O.; Sood, K.; Patel, H.S.; Malena, D.A.; Gajiwala, P.H.; Maciukiewicz, P.; Fournier, V.; Zayed, A. Chronic Exposure to Neonicotinoids Reduces Honey Bee Health near Corn Crops. Science 2017, 356, 1395–1397. [Google Scholar] [CrossRef]
- Belsky, J.E.; Joshi, N.K. Effects of Fungicide and Herbicide Chemical Exposure on Apis and Non-Apis Bees in Agricultural Landscape. Front. Environ. Sci. 2020, 8, 81. [Google Scholar] [CrossRef]
- Bohnenblust, E.W.; Vaudo, A.D.; Egan, J.F.; Mortensen, D.A.; Tooker, J.F. Effects of the Herbicide Dicamba on Nontarget Plants and Pollinator Visitation. Environ. Toxicol. Chem. 2016, 35, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Phillips, J.R.; Whitcomb, W.H.; Dowell, G.C.; Boyer, W.P.; Bell, K.O.; Dean, G.L.; Matthews, E.J.; Graves, B.; Newsom, L.D.; Clower, D.F.; et al. Toxicity of Herbicides to Newly Emerged Honey Bees. Environ. Entomol. 1972, 1, 102–104. [Google Scholar] [CrossRef]
- Kakumanu, M.L.; Reeves, A.M.; Anderson, T.D.; Rodrigues, R.R.; Williams, M.A. Honey Bee Gut Microbiome Is Altered by In-Hive Pesticide Exposures. Front. Microbiol. 2016, 7, 1255. [Google Scholar] [CrossRef]
- Motta, E.V.S.; Raymann, K.; Moran, N.A. Glyphosate Perturbs the Gut Microbiota of Honey Bees. Proc. Natl. Acad. Sci. USA 2018, 115, 10305–10310. [Google Scholar] [CrossRef]
- Koch, H.; Schmid-Hempel, P. Socially Transmitted Gut Microbiota Protect Bumble Bees against an Intestinal Parasite. Proc. Natl. Acad. Sci. USA 2011, 108, 19288–19292. [Google Scholar] [CrossRef]
- Mockler, B.K.; Kwong, W.K.; Moran, N.A.; Koch, H. Microbiome Structure Influences Infection by the Parasite Crithidia bombi in Bumble Bees. Appl. Environ. Microbiol. 2018, 84, e02335. [Google Scholar] [CrossRef]
- Runting, R.K.; Mayfield, M.M. Abundance and Diversity of Pollinators on Green Roofs Are Affected by Environmental Factors. IOP Conf. Ser. Earth Environ. Sci. 2019, 358, 022053. [Google Scholar] [CrossRef]
- Zheng, H.; Powell, J.E.; Steele, M.I.; Dietrich, C.; Moran, N.A. Honeybee Gut Microbiota Promotes Host Weight Gain via Bacterial Metabolism and Hormonal Signaling. Proc. Natl. Acad. Sci. USA 2017, 114, 4775–4780. [Google Scholar] [CrossRef]
- Pettis, J.S.; Lichtenberg, E.M.; Andree, M.; Stitzinger, J.; Rose, R.; van Engelsdorp, D. Crop Pollination Exposes Honey Bees to Pesticides Which Alters Their Susceptibility to the Gut Pathogen Nosema ceranae. PLoS ONE 2013, 8, e70182. [Google Scholar] [CrossRef]
- Wu, J.Y.; Smart, M.D.; Anelli, C.M.; Sheppard, W.S. Honey Bees (Apis mellifera) Reared in Brood Combs Containing High Levels of Pesticide Residues Exhibit Increased Susceptibility to Nosema (Microsporidia) Infection. J. Invertebr. Pathol. 2012, 109, 326–329. [Google Scholar] [CrossRef] [PubMed]
- Cohen, H.; McFrederick, Q.S.; Philpott, S.M. Environment Shapes the Microbiome of the Blue Orchard Bee, Osmia lignaria. Microb. Ecol. 2020, 80, 897–907. [Google Scholar] [CrossRef] [PubMed]
- Engel, P.; Kwong, W.K.; McFrederick, Q.; Anderson, K.E.; Barribeau, S.M.; Chandler, J.A.; Cornman, R.S.; Dainat, J.; de Miranda, J.R.; Doublet, V.; et al. The Bee Microbiome: Impact on Bee Health and Model for Evolution and Ecology of Host-Microbe Interactions. mBio 2016, 7, e02164-15. [Google Scholar] [CrossRef] [PubMed]
- Alaniz, A.J.; Carvajal, M.A.; Vergara, P.M. Giants Are Coming? Predicting the Potential Spread and Impacts of the Giant Asian Hornet (Vespa mandarinia, Hymenoptera: Vespidae) in the USA. Pest Manag. Sci. 2021, 77, 104–112. [Google Scholar] [CrossRef]
- Wilson, T.M.; Takahashi, J.; Spichiger, S.E.; Kim, I.; van Westendorp, P. First Reports of Vespa mandarinia (Hymenoptera: Vespidae) in North America Represent Two Separate Maternal Lineages in Washington State, United States, and British Columbia, Canada. Ann. Entomol. Soc. Am. 2020, 113, 468–472. [Google Scholar] [CrossRef]
- Matsuura, M.; Yamane, S. Biology of the Vespine Wasps, 1st ed.; Springer: Berlin, Germany, 1990; ISBN 9783540519003. [Google Scholar]
- Jones, E.I.; Dornhaus, A. Predation Risk Makes Bees Reject Rewarding Flowers and Reduce Foraging Activity. Behav. Ecol. Sociobiol. 2011, 65, 1505–1511. [Google Scholar] [CrossRef]
- Eves, J. Biology of Monodontomerus obscurus Westwood, a Parasite of the Alfalfa Leafcutting Bee, Megachile rotundata (Fabricius) (Hymenoptera: Torymidae; Megachilidae). Melanderia 1970, 4, 1–18. [Google Scholar]
- Straka, J. Strepsiptera of Canada. Zookeys 2019, 819, 377–382. [Google Scholar] [CrossRef]
- Balzer, Z.S.; Davis, A.R. Anatomical Impact and Notes on Viviparous Development Related to Infestation by Neotenic Females of Stylops advarians (Strepsiptera: Stylopidae) on Adult Female Andrena Milwaukeensis (Hymenoptera: Andrenidae). Can. Entomol. 2021, 153, 500–510. [Google Scholar] [CrossRef]
- Wood, T.J.; Michez, D.; Paxton, R.J.; Drossart, M.; Neumann, P.; Gérard, M.; Vanderplanck, M.; Barraud, A.; Martinet, B.; Leclercq, N.; et al. Managed Honey Bees as a Radar for Wild Bee Decline? Apidologie 2020, 51, 1100–1116. [Google Scholar] [CrossRef]
- Habermannová, J.; Bogusch, P.; Straka, J. Flexible Host Choice and Common Host Switches in the Evolution of Generalist and Specialist Cuckoo Bees (Anthophila: Sphecodes). PLoS ONE 2013, 8, e64537. [Google Scholar] [CrossRef] [PubMed]
- Levin, M.D. Biological Notes on Osmia Lignaria and Osmia Californica (Hymenoptera: Apoidea, Megachilidae). J. Kans. Entomol. Soc. 1966, 39, 524–535. [Google Scholar]
- Minckley, R.L.; Danforth, B.N. Sources and Frequency of Brood Loss in Solitary Bees. Apidologie 2019, 50, 515–525. [Google Scholar] [CrossRef]
- Baz, A. Bark-Lice, Book-Lice or Psocids (Psocoptera). In Encyclopedia of Entomology; Capinera, J.L., Ed.; Springer Dordrecht: Berlin, Germany, 2008; pp. 381–399. ISBN 978-1-4020-6242-1. [Google Scholar]
- Zhantiev, R.D. Ecology and Classification of Dermestid Beetles (Coleoptera, Dermestidae) of the Palaearctic Fauna. Entomol. Rev. 2009, 89, 157–174. [Google Scholar] [CrossRef]
- Dolezal, A.G.; Hendrix, S.D.; Scavo, N.A.; Carrillo-Tripp, J.; Harris, M.A.; Wheelock, M.J.; O’Neal, M.E.; Toth, A.L. Honey Bee Viruses in Wild Bees: Viral Prevalence, Loads, and Experimental Inoculation. PLoS ONE 2016, 11, e0166190. [Google Scholar] [CrossRef]
- Radzevičiūtė, R.; Theodorou, P.; Husemann, M.; Japoshvili, G.; Kirkitadze, G.; Zhusupbaeva, A.; Paxton, R.J. Replication of Honey Bee-Associated RNA Viruses across Multiple Bee Species in Apple Orchards of Georgia, Germany and Kyrgyzstan. J. Invertebr. Pathol. 2017, 146, 14–23. [Google Scholar] [CrossRef]
- Genersch, E. American Foulbrood in Honeybees and Its Causative Agent, Paenibacillus Larvae. J. Invertebr. Pathol. 2010, 103, S10–S19. [Google Scholar] [CrossRef]
- Voulgari-Kokota, A.; Steffan-Dewenter, I.; Keller, A. Susceptibility of Red Mason Bee Larvae to Bacterial Threats Due to Microbiome Exchange with Imported Pollen Provisions. Insects 2020, 11, 373. [Google Scholar] [CrossRef]
- Vandenberg, J.D.; Stephen, W.P. Etiology and Symptomatology of Chalkbrood in the Alfalfa Leafcutting Bee, Megachile rotundata. J. Invertebr. Pathol. 1982, 39, 133–137. [Google Scholar] [CrossRef]
- Otti, O.; Schmid-Hempel, P. Nosema Bombi: A Pollinator Parasite with Detrimental Fitness Effects. J. Invertebr. Pathol. 2007, 96, 118–124. [Google Scholar] [CrossRef]
- Graystock, P.; Goulson, D.; Hughes, W.O.H. The Relationship between Managed Bees and the Prevalence of Parasites in Bumblebees. PeerJ 2014, 2, e522. [Google Scholar] [CrossRef] [PubMed]
- Graystock, P.; Meeus, I.; Smagghe, G.; Goulson, D.; Hughes, W.O.H. The Effects of Single and Mixed Infections of Apicystis Bombi and Deformed Wing Virus in Bombus terrestris. Parasitology 2016, 143, 358–365. [Google Scholar] [CrossRef] [PubMed]
- Schmid-Hempel, R.; Eckhardt, M.; Goulson, D.; Heinzmann, D.; Lange, C.; Plischuk, S.; Escudero, L.R.; Salathé, R.; Scriven, J.J.; Schmid-Hempel, P. The Invasion of Southern South America by Imported Bumblebees and Associated Parasites. J. Anim. Ecol. 2014, 83, 823–837. [Google Scholar] [CrossRef] [PubMed]
- Wenzel, A.; Grass, I.; Belavadi, V.V.; Tscharntke, T. How Urbanization Is Driving Pollinator Diversity and Pollination—A Systematic Review. Biol. Conserv. 2020, 241, 108321. [Google Scholar] [CrossRef]
- Kearns, C.A.; Inouye, D.W.; Waser, N.M. Endangered Mutualisms: The Conservation of Plant-Pollinator Interactions. Annu. Rev. Ecol. Syst. 1998, 29, 83–112. [Google Scholar] [CrossRef]
- Potts, S.G.; Vulliamy, B.; Roberts, S.; O’Toole, C.; Dafni, A.; Ne’eman, G.; Willmer, P. Role of Nesting Resources in Organising Diverse Bee Communities in a Mediterranean Landscape. Ecol. Entomol. 2005, 30, 78–85. [Google Scholar] [CrossRef]
- Vulliamy, B.; Potts, S.G.; Vulliamy, P.G.W.; Potts, B.; Willmer, S.G.; Vulliamy, B.; Willmer, P.G. The Effects of Cattle Grazing on Plant-Pollinator Communities in a Fragmented Mediterranean Landscape. Oikos 2006, 114, 529–543. [Google Scholar] [CrossRef]
- Cole, L.J.; Brocklehurst, S.; Robertson, D.; Harrison, W.; McCracken, D.I. Exploring the Interactions between Resource Availability and the Utilisation of Semi-Natural Habitats by Insect Pollinators in an Intensive Agricultural Landscape. Agric. Ecosyst. Environ. 2017, 246, 157–167. [Google Scholar] [CrossRef]
- Martins, K.T.; Albert, C.H.; Lechowicz, M.J.; Gonzalez, A. Complementary Crops and Landscape Features Sustain Wild Bee Communities. Ecol. Appl. 2018, 28, 1093–1105. [Google Scholar] [CrossRef]
- Koh, I.; Lonsdorf, E.V.; Artz, D.R.; Pitts-Singer, T.L.; Ricketts, T.H. Ecology and Economics of Using Native Managed Bees for Almond Pollination. J. Econ. Entomol. 2018, 111, 16–25. [Google Scholar] [CrossRef]
- Rahimi, E.; Barghjelveh, S.; Dong, P. How Effective Are Artificial Nests in Attracting Bees? A Review. J. Ecol. Environ. 2021, 45, 1–11. [Google Scholar] [CrossRef]
- Gaston, K.J.; Smith, R.M.; Thompson, K.; Warren, P.H. Urban Domestic Gardens (II): Experimental Tests of Methods for Increasing Biodiversity. Biodivers. Conserv. 2005, 14, 395–413. [Google Scholar] [CrossRef]
- Holzschuh, A.; Dudenhöffer, J.H.; Tscharntke, T. Landscapes with Wild Bee Habitats Enhance Pollination, Fruit Set and Yield of Sweet Cherry. Biol. Conserv. 2012, 153, 101–107. [Google Scholar] [CrossRef]
- Nicholls, C.I.; Altieri, M.A. Plant Biodiversity Enhances Bees and Other Insect Pollinators in Agroecosystems. A Review. Agron. Sustain. Dev. 2013, 33, 257–274. [Google Scholar] [CrossRef]
- Wood, T.J.; Kaplan, I.; Szendrei, Z. Wild Bee Pollen Diets Reveal Patterns of Seasonal Foraging Resources for Honey Bees. Front. Ecol. Evol. 2018, 6, 210. [Google Scholar] [CrossRef]
- Graham, K.K.; Perkins, J.A.; Peake, A.; Killewald, M.; Zavalnitskaya, J.; Wilson, J.K.; Isaacs, R. Wildflower Plantings on Fruit Farms Provide Pollen Resources and Increase Nesting by Stem Nesting Bees. Agric. For. Entomol. 2020, 432, 222–231. [Google Scholar] [CrossRef]
- Asner, G.P.; Elmore, A.J.; Olander, L.P.; Martin, R.E.; Harris, T. Grazing Systems, Ecosystem Responses, and Global Change. Annu. Rev. Environ. Resour. 2004, 29, 261–299. [Google Scholar] [CrossRef]
- Foley, K.; Fazio, G.; Jensen, A.B.; Hughes, W.O.H. Nutritional Limitation and Resistance to Opportunistic Aspergillus Parasites in Honey Bee Larvae. J. Invertebr. Pathol. 2012, 111, 68–73. [Google Scholar] [CrossRef]
- Kennedy, C.M.; Lonsdorf, E.; Neel, M.C.; Williams, N.M.; Ricketts, T.H.; Winfree, R.; Bommarco, R.; Brittain, C.; Burley, A.L.; Cariveau, D.; et al. A Global Quantitative Synthesis of Local and Landscape Effects on Wild Bee Pollinators in Agroecosystems. Ecol. Lett. 2013, 16, 584–599. [Google Scholar] [CrossRef]
- Klaus, F.; Tscharntke, T.; Bischoff, G.; Grass, I. Floral Resource Diversification Promotes Solitary Bee Reproduction and May Offset Insecticide Effects—Evidence from a Semi-Field Experiment. Ecol. Lett. 2021, 24, 668–675. [Google Scholar] [CrossRef]
- Williams, N.M.; Ward, K.L.; Pope, N.; Isaacs, R.; Wilson, J.; May, E.A.; Ellis, J.; Daniels, J.; Pence, A.; Ullmann, K.; et al. Native Wildflower Plantings Support Wild Bee Abundance and Diversity in Agricultural Landscapes across the United States. Ecol. Appl. 2015, 25, 2119–2131. [Google Scholar] [CrossRef] [PubMed]
- Alaux, C.; Ducloz, F.; Crauser, D.; le Conte, Y. Diet Effects on Honeybee Immunocompetence. Biol. Lett. 2010, 6, 562–565. [Google Scholar] [CrossRef]
- Brunner, F.S.; Schmid-Hempel, P.; Barribeau, S.M. Protein-Poor Diet Reduces Host-Specific Immune Gene Expression in Bombus terrestris. Proc. R. Soc. B Biol. Sci. 2014, 281, 20140128. [Google Scholar] [CrossRef]
- Gresty, C.E.A.; Clare, E.; Devey, D.S.; Cowan, R.S.; Csiba, L.; Malakasi, P.; Lewis, O.T.; Willis, K.J. Flower Preferences and Pollen Transport Networks for Cavity-Nesting Solitary Bees: Implications for the Design of Agri-Environment Schemes. Ecol. Evol. 2018, 8, 7574–7587. [Google Scholar] [CrossRef] [PubMed]
- Bartomeus, I.; Ascher, J.S.; Gibbs, J.; Danforth, B.N.; Wagner, D.L.; Hedtke, S.M.; Winfree, R. Historical Changes in Northeastern US Bee Pollinators Related to Shared Ecological Traits. Proc. Natl. Acad. Sci. USA 2013, 110, 4656–4660. [Google Scholar] [CrossRef] [PubMed]
- Frankie, G.W.; Rizzardi, M.; Vinson, S.B.; Griswold, T.L. Decline in Bee Diversity and Abundance from 1972–2004 on a Flowering Leguminous Tree, Andira Inermis in Costa Rica at the Interface of Disturbed Dry Forest and the Urban Environment. J. Kans. Entomol. Soc. 2009, 82, 1–20. [Google Scholar] [CrossRef]
- Ricketts, T.H.; Daily, G.C.; Ehrlich, P.R.; Michener, C.D. Economic Value of Tropical Forest to Coffee Production. Proc. Natl. Acad. Sci. USA 2004, 101, 12579–12582. [Google Scholar] [CrossRef] [PubMed]
- Kline, O.; Joshi, N.K. Mitigating the Effects of Habitat Loss on Solitary Bees in Agricultural Ecosystems. Agriculture 2020, 10, 115. [Google Scholar] [CrossRef]
- Westrich, P. Habitat Requirements of Central European Bees and the Problems of Partial Habitats. In Linnean Society Symposium Series; Academic Press: Cambridge, MA, USA, 1996; Volume 18, pp. 1–16. ISBN 0-12-479740-7. [Google Scholar]
- Cane, J.H. Nesting Resins Obtained from Larrea Pollen Host by an Oligolectic Bee, Trachusa larreae (Cockerell) (Hymenoptera: Megachilidae). J. Kans. Entomol. Soc. 1996, 69, 99–102. [Google Scholar]
- Cane, J.H. A Native Ground-Nesting Bee (Nomia melanderi) Sustainably Managed to Pollinate Alfalfa across an Intensively Agricultural Landscape. Apidologie 2008, 39, 315–323. [Google Scholar] [CrossRef]
- Vinchesi, A.; Cobos, D.; Lavine, L.; Walsh, D. Manipulation of Soil Temperatures to Influence Brood Emergence in the Alkali Bee (Nomia Melanderi). Apidologie 2013, 44, 286–294. [Google Scholar] [CrossRef][Green Version]
- Julier, H.E.; Roulston, T.H. Wild Bee Abundance and Pollination Service in Cultivated Pumpkins: Farm Management, Nesting Behavior and Landscape Effects. J. Econ. Entomol. 2009, 102, 563–573. [Google Scholar] [CrossRef] [PubMed]
- Skidmore, A.R.; Short, C.A.; Dills, C.; Goodell, K.; Bessin, R.T. Preference of Peponapis pruinosa (Hymenoptera: Apoidea) for Tilled Soils Regardless of Soil Management System. Environ. Entomol. 2019, 48, 961–967. [Google Scholar] [CrossRef] [PubMed]
- Minckley, R.L.; Wcislo, W.T.; Yanega, D.; Buchmann, S.L. Behavior and Phenology of a Specialist Bee (Dieunomia) and Sunflower (Helianthus) Pollen Availability. Ecology 1994, 75, 1406–1419. [Google Scholar] [CrossRef]
- Cane, J.H.; Neff, J.L. Predicted Fates of Ground-Nesting Bees in Soil Heated by Wildfire: Thermal Tolerances of Life Stages and a Survey of Nesting Depths. Biol. Conserv. 2011, 144, 2631–2636. [Google Scholar] [CrossRef]
- Harmon-Threatt, A. Influence of Nesting Characteristics on Health of Wild Bee Communities. Annu. Rev. Entomol. 2020, 65, 39–56. [Google Scholar] [CrossRef]
- Sann, M.; Niehuis, O.; Peters, R.S.; Mayer, C.; Kozlov, A.; Podsiadlowski, L.; Bank, S.; Meusemann, K.; Misof, B.; Bleidorn, C.; et al. Phylogenomic Analysis of Apoidea Sheds New Light on the Sister Group of Bees. BMC Evol. Biol. 2018, 18, 71. [Google Scholar] [CrossRef]
- Torchio, P.F. The Biology of Perdita Nuda and Descriptions of Its Immature Forms and Those of Its Sphecodes Parasite (Hymenoptera: Apoidea). J. Kans. Entomol. Soc. 1975, 48, 257–279. [Google Scholar]
- Danforth, B.N. Emergence Dynamics and Bet Hedging in a Desert Bee, Perdita portalis. Proc. R. Soc. B Biol. Sci. 1999, 266, 1985–1994. [Google Scholar] [CrossRef]
- Vaughan, M.; Hopwood, J.; Lee-Mäder, E.; Shepherd, M.; Kremen, C.; Stine, A.; Black, S.H. Farming for Bees—Guidelines for Providing Native Bee Habitat on Farms, 4th ed.; West, K., Morris, S., Shepherd, M., Eds.; The Xerces Society for Invertebrate Conservation: Portland, OR, USA, 2015. [Google Scholar]
- Brockmann, H.J. Nest-Site Selection in the Great Golden Digger Wasp, Sphex Ichneumoneus L. (Sphecidae). Ecol. Entomol. 1979, 4, 211–224. [Google Scholar] [CrossRef]
- Potts, S.G.; Willmer, P. Abiotic and Biotic Factors Influencing Nest-Site Selection by Halictus rubicundus, a Ground-Nesting Halictine Bee. Ecol. Entomol. 1997, 22, 319–328. [Google Scholar] [CrossRef]
- Brünnert, U.; Kelber, A.; Zeil, J. Ground-Nesting Bees Determine the Location of Their Nest Relative to a Landmark by Other than Angular Size Cues. J. Comp. Physiol. A 1994, 175, 363–369. [Google Scholar] [CrossRef]
- Packer, L.; Sampson, B.; Lockerbie, C.; Jessome, V. Nest Architecture and Brood Mortality in Four Species of Sweat Bee (Hymenoptera; Halictidae) from Cape Breton Island. Can. J. Zool. 2011, 67, 2864–2870. [Google Scholar] [CrossRef]
- Antoine, C.M.; Forrest, J.R.K. Nesting Habitat of Ground-Nesting Bees: A Review. Ecol. Entomol. 2021, 46, 143–159. [Google Scholar] [CrossRef]
- Bihaly, Á.D.; Kovács-Hostyánszki, A.; Szalai, M.; Sárospataki, M. Nesting Activity of Cavity-Nesting Bees and Wasps Is Lower in Small-Scale Apple Orchards Compared to Nearby Semi-Natural Habitats. Agric. For. Entomol. 2021, 23, 49–58. [Google Scholar] [CrossRef]
- Requier, F.; Leonhardt, S.D. Beyond Flowers: Including Non-Floral Resources in Bee Conservation Schemes. J. Insect Conserv. 2020, 24, 5–16. [Google Scholar] [CrossRef]
- Kovács-Hostyánszki, A.; Espíndola, A.; Vanbergen, A.J.; Settele, J.; Kremen, C.; Dicks, L.V. Ecological Intensification to Mitigate Impacts of Conventional Intensive Land Use on Pollinators and Pollination. Ecol. Lett. 2017, 20, 673–689. [Google Scholar] [CrossRef] [PubMed]
- Gill, R.J.; Ramos-Rodriguez, O.; Raine, N.E. Combined Pesticide Exposure Severely Affects Individual- and Colony-Level Traits in Bees. Nature 2012, 491, 105–108. [Google Scholar] [CrossRef]
- Watson, J.C.; Wolf, A.T.; Ascher, J.S. Forested Landscapes Promote Richness and Abundance of Native Bees (Hymenoptera: Apoidea: Anthophila) in Wisconsin Apple Orchards. Environ. Entomol. 2011, 40, 621–632. [Google Scholar] [CrossRef] [PubMed]
- Fabian, Y.; Sandau, N.; Bruggisser, O.T.; Aebi, A.; Kehrli, P.; Rohr, R.P.; Naisbit, R.E.; Bersier, L.F. The Importance of Landscape and Spatial Structure for Hymenopteran-Based Food Webs in an Agro-Ecosystem. J. Anim. Ecol. 2013, 82, 1203–1214. [Google Scholar] [CrossRef]
- Földesi, R.; Kovács-Hostyánszki, A.; Korösi, Á.; Somay, L.; Elek, Z.; Markó, V.; Sárospataki, M.; Bakos, R.; Varga, Á.; Nyisztor, K.; et al. Relationships between Wild Bees, Hoverflies and Pollination Success in Apple Orchards with Different Landscape Contexts. Agric. For. Entomol. 2016, 18, 68–75. [Google Scholar] [CrossRef]
- Mallinger, R.E.; Gibbs, J.; Gratton, C. Diverse Landscapes Have a Higher Abundance and Species Richness of Spring Wild Bees by Providing Complementary Floral Resources over Bees’ Foraging Periods. Landsc. Ecol. 2016, 31, 1523–1535. [Google Scholar] [CrossRef]
- Ulyshen, M.D.; Soon, V.; Hanula, J.L. On the Vertical Distribution of Bees in a Temperate Deciduous Forest. Insect Conserv. Divers. 2010, 3, 222–228. [Google Scholar] [CrossRef]
- Mader, E.; Spivak, M.; Evans, E. Managing Alternative Pollinators: A Handbook for Beekeepers, Growers, and Conservationists; SARE and NRAES: Ithaca, NY, USA; College Park, MD, USA, 2010. [Google Scholar]
- Bortolotti, L.; Bogo, G.; de Manincor, N.; Fisogni, A.; Galloni, M. Integrated Conservation of Bee Pollinators of a Rare Plant in a Protected Area near Bologna, Italy. Conserv. Evid. 2016, 13, 51–56. [Google Scholar]
- Ritz, A.; Sahli, H.; Biddinger, D.; Schupp, J.; Rajotte, E.; Joshi, N.K. Quantifying the Efficacy of Native Bees for Orchard Pollination in Pennsylvania to Offset the Increased Cost and Decreased Reliability of Honeybees. Penn Fruit News 2012, 92, 6–66. [Google Scholar]
- Westerkamp, C. Honey Bees Are Poor Pollinators—Why? Plant Syst. Evol. 1991, 177, 71–75. [Google Scholar] [CrossRef]
- Martins, K.T.; Gonzalez, A.; Lechowicz, M.J. Pollination Services Are Mediated by Bee Functional Diversity and Landscape Context. Agric. Ecosyst. Environ. 2015, 200, 12–20. [Google Scholar] [CrossRef]
- Ramalho, M.; Imperatriz-Fonseca, V.L.; Giannini, T.C. Within-Colony Size Variation of Foragers and Pollen Load Capacity in the Stingless Bee Melipona Quadrifasciata Anthidioides Lepeletier (Apidae, Hymenoptera). Apidologie 1998, 29, 221–228. [Google Scholar] [CrossRef]
- Streinzer, M.; Huber, W.; Spaethe, J. Body Size Limits Dim-Light Foraging Activity in Stingless Bees (Apidae: Meliponini). J. Comp. Physiol. A 2016, 202, 643–655. [Google Scholar] [CrossRef]
- Robinson, W.S.; Fell, R.D. Effect of Honey Bee Foraging Behaviors on’Delicious’ Apple Set. HortScience 1981, 16, 326–328. [Google Scholar] [CrossRef]
- Free, J.B. Insect Pollination of Crops, 2nd ed.; Academic Press: London, UK, 1993; ISBN 9780122666513. [Google Scholar]
- Schneider, D.; Stern, R.A.; Eisikowitch, D.; Goldway, M. The Relationship between Floral Structure and Honeybee Pollination Eficiency in ‘Jonathan’ and ‘Topred’ Apple Cultivars. J. Hortic. Sci. Biotechnol. 2002, 77, 48–51. [Google Scholar] [CrossRef]
- Bohart, G.E.; Todd, F.E. Pollination of Seed Crops by Insects. In Yearbook of Agriculture; US Department of Agriculture: Washington, DC, USA, 1961; ISBN 630297-62. [Google Scholar]
- Greenleaf, S.A.S. Local-Scale and Foraging-Scale Habitats Affect Bee Community Abundance, Species Richness, and Pollination Services in Northern California; Princeton University: Ann Arbor, MI, USA, 2005. [Google Scholar]
- Greenleaf, S.S.; Kremen, C. Wild Bee Species Increase Tomato Production and Respond Differently to Surrounding Land Use in Northern California. Biol. Conserv. 2006, 133, 81–87. [Google Scholar] [CrossRef]
- Thomson, J.D.; Goodell, K. Pollen Removal and Deposition by Honeybee and Bumblebee Visitors to Apple and Almond Flowers. J. Appl. Ecol. 2001, 38, 1032–1044. [Google Scholar] [CrossRef]
- Isaacs, R.; Tuell, J.; Fiedler, A.; Gardiner, M.; Landis, D. Maximizing Arthropod-Mediated Ecosystem Services in Agricultural Landscapes: The Role of Native Plants. Front. Ecol. Environ. 2009, 7, 196–203. [Google Scholar] [CrossRef]
- Pywell, R.F.; Heard, M.S.; Woodcock, B.A.; Hinsley, S.; Ridding, L.; Nowakowski, M.; Bullock, J.M. Wildlife-Friendly Farming Increases Crop Yield: Evidence for Ecological Intensification. Proc. R. Soc. B Biol. Sci. 2015, 282, 20151740. [Google Scholar] [CrossRef]
- Donkersley, P.; Rhodes, G.; Pickup, R.W.; Jones, K.C.; Power, E.F.; Wright, G.A.; Wilson, K. Nutritional Composition of Honey Bee Food Stores Vary with Floral Composition. Oecologia 2017, 185, 749–761. [Google Scholar] [CrossRef]
- Tuell, J.K.; Fiedler, A.K.; Landis, D.; Isaacs, R. Visitation by Wild and Managed Bees (Hymenoptera: Apoidea) to Eastern U.S. Native Plants for Use in Conservation Programs. Environ. Entomol. 2008, 37, 707–718. [Google Scholar] [CrossRef]
- Holland, J.M.; Smith, B.M.; Storkey, J.; Lutman, P.J.W.; Aebischer, N.J. Managing Habitats on English Farmland for Insect Pollinator Conservation. Biol. Conserv. 2015, 182, 215–222. [Google Scholar] [CrossRef]
- Fitzgerald, J.D.; Solomon, M.G. Can Flowering Plants Enhance Numbers of Beneficial Arthropods in UK Apple and Pear Orchards? Biocontrol Sci. Technol. 2010, 14, 291–300. [Google Scholar] [CrossRef]
- Boyle, N.K.; Pitts-Singer, T.L. Assessing Blue Orchard Bee (Osmia lignaria) Propagation and Pollination Services in the Presence of Honey Bees (Apis mellifera) in Utah Tart Cherries. PeerJ 2019, 7, e7639. [Google Scholar] [CrossRef]
- Ratto, F.; Steward, P.; Sait, S.M.; Pryke, J.S.; Gaigher, R.; Samways, M.J.; Kunin, W. Proximity to Natural Habitat and Flower Plantings Increases Insect Populations and Pollination Services in South African Apple Orchards. J. Appl. Ecol. 2021, 58, 2540–2551. [Google Scholar] [CrossRef]
- Campbell, A.J.; Wilby, A.; Sutton, P.; Wäckers, F.L. Do Sown Flower Strips Boost Wild Pollinator Abundance and Pollination Services in a Spring-Flowering Crop? A Case Study from UK Cider Apple Orchards. Agric. Ecosyst. Environ. 2017, 239, 20–29. [Google Scholar] [CrossRef]
- Muñoz, A.E.; Amouroux, P.; Zaviezo, T. Native Flowering Shrubs Promote Beneficial Insects in Avocado Orchards. Agric. For. Entomol. 2021, 23, 463–472. [Google Scholar] [CrossRef]
- Peace, A.; Pattemore, D.; Broussard, M.; Fonseka, D.; Tomer, N.; Bosque-Pérez, N.A.; Crowder, D.; Shaw, A.K.; Jesson, L.; Howlett, B.G.; et al. Orchard Layout and Plant Traits Influence Fruit Yield More Strongly than Pollinator Behaviour and Density in a Dioecious Crop. PLoS ONE 2020, 15, e0231120. [Google Scholar] [CrossRef]
- Kratschmer, S.; Pachinger, B.; Gaigher, R.; Pryke, J.S.; van Schalkwyk, J.; Samways, M.J.; Melin, A.; Kehinde, T.; Zaller, J.G.; Winter, S. Enhancing Flowering Plant Functional Richness Improves Wild Bee Diversity in Vineyard Inter-Rows in Different Floral Kingdoms. Ecol. Evol. 2021, 11, 7927–7945. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.; Groom, S.V.C.; Rader, R.; Hogendoorn, K.; Cunningham, S.A. Land Cover Associations of Wild Bees Visiting Flowers in Apple Orchards across Three Geographic Regions of Southeast Australia. Agric. Ecosyst. Environ. 2022, 324, 107717. [Google Scholar] [CrossRef]
- Eeraerts, M.; Smagghe, G.; Meeus, I. Pollinator Diversity, Floral Resources and Semi-Natural Habitat, Instead of Honey Bees and Intensive Agriculture, Enhance Pollination Service to Sweet Cherry. Agric. Ecosyst. Environ. 2019, 284, 106586. [Google Scholar] [CrossRef]
- Wu, P.; Axmacher, J.C.; Li, X.; Song, X.; Yu, Z.; Xu, H.; Tscharntke, T.; Westphal, C.; Liu, Y. Contrasting Effects of Natural Shrubland and Plantation Forests on Bee Assemblages at Neighboring Apple Orchards in Beijing, China. Biol. Conserv. 2019, 237, 456–462. [Google Scholar] [CrossRef]
- Stroot, L.; Brinkert, A.; Hölzel, N.; Rüsing, A.; Bucharova, A. Establishment of Wildflower Strips in a Wide Range of Environments: A Lesson from a Landscape-Scale Project. Restor. Ecol. 2022, 30, e13542. [Google Scholar] [CrossRef]
- Schmidt, A.; Kirmer, A.; Kiehl, K.; Tischew, S. Seed Mixture Strongly Affects Species-Richness and Quality of Perennial Flower Strips on Fertile Soil. Basic Appl. Ecol. 2020, 42, 62–72. [Google Scholar] [CrossRef]
- Fuccillo Battle, K.; de Rivera, C.E.; Cruzan, M.B. The Role of Functional Diversity and Facilitation in Small-Scale Pollinator Habitat. Ecol. Appl. 2021, 31, e02355. [Google Scholar] [CrossRef] [PubMed]
- Wen, A.; Elgersma, K.J.; Sherrard, M.E.; Jackson, L.L.; Meissen, J.; Myers, M.C. Wild Bee Visitors and Their Association with Sown and Unsown Floral Resources in Reconstructed Pollinator Habitats within an Agriculture Landscape. Insect Conserv. Divers. 2022, 15, 102–113. [Google Scholar] [CrossRef]
- Harder, L.D. Morphology as a Predictor of Flower Choice by Bumble Bees. Ecology 1985, 66, 198–210. [Google Scholar] [CrossRef]
- Rodríguez-Gironés, M.A.; Santamaría, L. Models of Optimal Foraging and Resource Partitioning: Deep Corollas for Long Tongues. Behav. Ecol. 2006, 17, 905–910. [Google Scholar] [CrossRef]
- Kohler, F.; Verhulst, J.; van Klink, R.; Kleijn, D. At What Spatial Scale Do High-Quality Habitats Enhance the Diversity of Forbs and Pollinators in Intensively Farmed Landscapes? J. Appl. Ecol. 2008, 45, 753–762. [Google Scholar] [CrossRef]
- Morandin, L.A.; Kremen, C. Hedgerow Restoration Promotes Pollinator Populations and Exports Native Bees to Adjacent Fields. Ecol. Appl. 2013, 23, 829–839. [Google Scholar] [CrossRef] [PubMed]
- M’Gonigle, L.K.; Ponisio, L.C.; Cutler, K.; Kremen, C. Habitat Restoration Promotes Pollinator Persistence and Colonization in Intensively Managed Agriculture. Ecol. Appl. 2015, 25, 1557–1565. [Google Scholar] [CrossRef]
- Ponisio, L.C.; M’Gonigle, L.K.; Kremen, C. On-Farm Habitat Restoration Counters Biotic Homogenization in Intensively Managed Agriculture. Glob. Chang. Biol. 2016, 22, 704–715. [Google Scholar] [CrossRef]
- Biddinger, D.J.; Surcică, A.; Joshi, N.K. A Native Predator Utilising the Invasive Brown Marmorated Stink Bug, Halyomorpha halys (Hemiptera: Pentatomidae) as a Food Source. Biocontrol Sci. Technol. 2017, 27, 903–907. [Google Scholar] [CrossRef]
- Sidhu, C.S.; Joshi, N.K. Establishing Wildflower Pollinator Habitats in Agricultural Farmland to Provide Multiple Ecosystem Services. Front. Plant Sci. 2016, 7, 363. [Google Scholar] [CrossRef]
- Mancini, F.; Woodcock, B.A.; Isaac, N.J.B. Agrochemicals in the Wild: Identifying Links between Pesticide Use and Declines of Nontarget Organisms. Curr. Opin. Environ. Sci. Health 2019, 11, 53–58. [Google Scholar] [CrossRef]
- Ansell, G.R.; Frewin, A.J.; Gradish, A.E.; Scott-Dupree, C.D. Contact Toxicity of Three Insecticides for Use in Tier I Pesticide Risk Assessments with Megachile rotundata (Hymenoptera: Megachilidae). PeerJ 2021, 9, e10744. [Google Scholar] [CrossRef] [PubMed]
- Thompson, H.M.; Pamminger, T. Are Honeybees Suitable Surrogates for Use in Pesticide Risk Assessment for Non-Apis Bees? Pest Manag. Sci. 2019, 75, 2549–2557. [Google Scholar] [CrossRef]
- Joshi, N.K.; Leslie, T.; Rajotte, E.G.; Biddinger, D.J. Environmental Impacts of Reduced-Risk and Conventional Pesticide Programs Differ in Commercial Apple Orchards, but Similarly Influence Pollinator Community. Chemosphere 2020, 240, 124926. [Google Scholar] [CrossRef] [PubMed]
- Kremen, C.; Williams, N.M.; Thorp, R.W. Crop Pollination from Native Bees at Risk from Agricultural Intensification. Proc. Natl. Acad. Sci. USA 2002, 99, 16812–16816. [Google Scholar] [CrossRef]
- Kremen, C.; M’Gonigle, L.K. Small-Scale Restoration in Intensive Agricultural Landscapes Supports More Specialized and Less Mobile Pollinator Species. J. Appl. Ecol. 2015, 52, 602–610. [Google Scholar] [CrossRef]
- Morandin, L.A.; Long, R.F.; Kremen, C. Pest Control and Pollination Cost–Benefit Analysis of Hedgerow Restoration in a Simplified Agricultural Landscape. J. Econ. Entomol. 2016, 109, 1020–1027. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kline, O.; Phan, N.T.; Porras, M.F.; Chavana, J.; Little, C.Z.; Stemet, L.; Acharya, R.S.; Biddinger, D.J.; Reddy, G.V.P.; Rajotte, E.G.; et al. Biology, Genetic Diversity, and Conservation of Wild Bees in Tree Fruit Orchards. Biology 2023, 12, 31. https://doi.org/10.3390/biology12010031
Kline O, Phan NT, Porras MF, Chavana J, Little CZ, Stemet L, Acharya RS, Biddinger DJ, Reddy GVP, Rajotte EG, et al. Biology, Genetic Diversity, and Conservation of Wild Bees in Tree Fruit Orchards. Biology. 2023; 12(1):31. https://doi.org/10.3390/biology12010031
Chicago/Turabian StyleKline, Olivia, Ngoc T. Phan, Mitzy F. Porras, Joshua Chavana, Coleman Z. Little, Lilia Stemet, Roshani S. Acharya, David J. Biddinger, Gadi V. P. Reddy, Edwin G. Rajotte, and et al. 2023. "Biology, Genetic Diversity, and Conservation of Wild Bees in Tree Fruit Orchards" Biology 12, no. 1: 31. https://doi.org/10.3390/biology12010031
APA StyleKline, O., Phan, N. T., Porras, M. F., Chavana, J., Little, C. Z., Stemet, L., Acharya, R. S., Biddinger, D. J., Reddy, G. V. P., Rajotte, E. G., & Joshi, N. K. (2023). Biology, Genetic Diversity, and Conservation of Wild Bees in Tree Fruit Orchards. Biology, 12(1), 31. https://doi.org/10.3390/biology12010031