
Dated Phylogeny of Astragalus Section Stereothrix (Fabaceae) and Allied Taxa in the Hypoglottis Clade


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Taxon Sampling
2.2. DNA Extraction, PCR Amplification, and DNA Sequencing
2.3. Sequence Alignments
2.4. Phylogenetic Inferences
2.5. Incongruent Length Difference Test
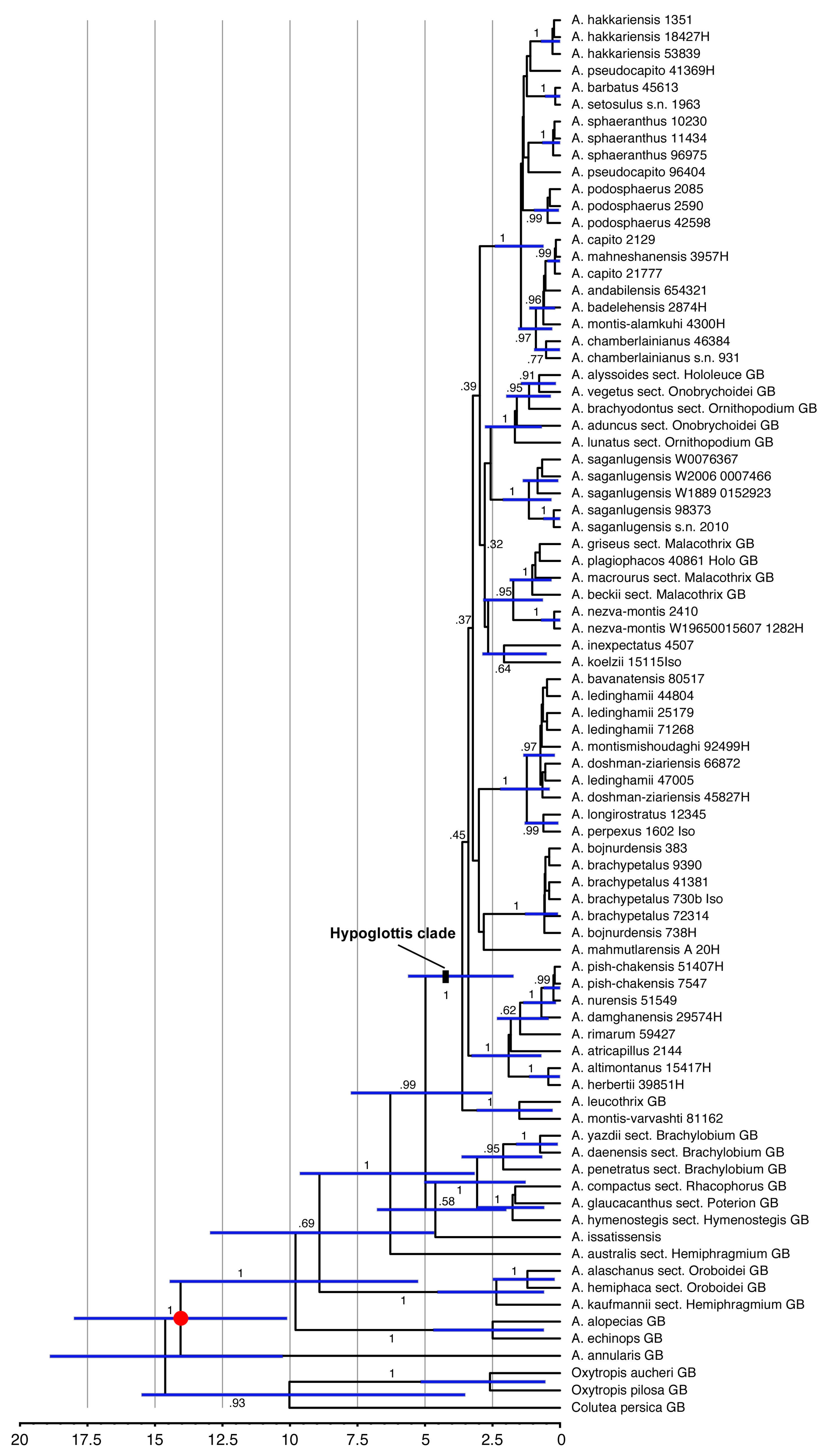
2.6. Divergence Time Estimation
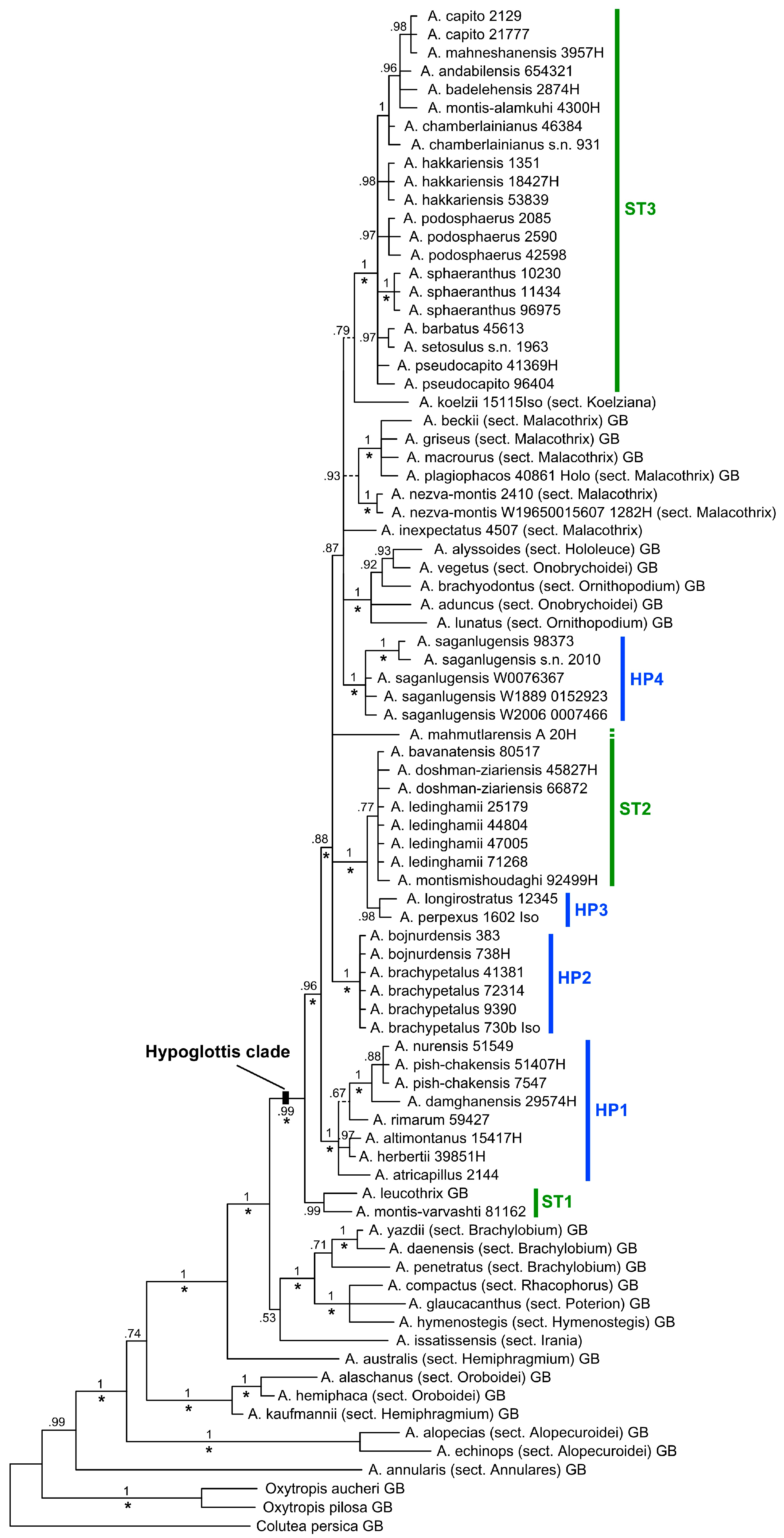
3. Results
3.1. Phylogenetic Analyses
3.2. Phylogenetic Reconstructions and Age Estimates
4. Discussion
4.1. Non-Monophyly of Bunge’s Traditional Subgenus Hypoglottis vs. Monophyly of the Hypoglottis Clade
4.2. Divergence Times and Fast and Young Diversification
4.3. Non-Monophyly of Sections Stereothrix, Hypoglottidei, and Their Allied Taxa
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Podlech, D.; Zarre, S. A Taxonomic Revision of the Genus Astragalus L. (Leguminosae) in the Old World, 1st ed.; Naturhistorisches Museum Wien: Vienna, Austria, 2013; Volume 1–3, p. 2439. [Google Scholar]
- Maassoumi, A.A.; Ashouri, P. The hotspots and conservation gaps of the mega genus Astragalus (Fabaceae) in the Old-World. Biodivers. Conserv. 2022, 31, 2119–2139. [Google Scholar] [CrossRef]
- Bunge, A. Generis Astragali species Gerontogeae. Pars prior, claves diagnosticae. Mem. Pres. Acad. Imp. Sci. St. Petersb. Septième Série 1868, 11, 1–140. [Google Scholar]
- Bunge, A. Generis Astragali species Gerontogeae. Pars altera, Specierum enumeration. Mem. Pres. Acad. Imp. Sci. St. Petersb. Septième Série 1869, 15, 1–245. [Google Scholar]
- Azani, N.; Bruneau, A.; Wojciechowski, M.F.; Zarre, S. Molecular phylogenetics of annual Astragalus (Fabaceae) and its systematic implications. Bot. J. Linn. Soc. 2017, 184, 347–365. [Google Scholar] [CrossRef]
- Azani, N.; Bruneau, A.; Wojciechowski, M.F.; Zarre, S. Miocene climate change as a driving force for multiple origins of annual species in Astragalus (Fabaceae, Papilionoideae). Mol. Phylogenet. Evol. 2019, 137, 210–221. [Google Scholar] [CrossRef]
- Bagheri, A.; Maassoumi, A.A.; Rahiminejad, M.R.; Brassac, J.; Blattner, F.R. Molecular phylogeny and divergence times of Astragalus section Hymenostegis: An analysis of a rapidly diversifying species group in Fabaceae. Sci. Rep. 2017, 7, 14033. [Google Scholar] [CrossRef]
- Kazempour Osaloo, S.; Maassoumi, A.A.; Murakami, N. Molecular systematic of the genus Astragalus L. (Fabaceae): Phylogenetic analyses of nuclear ribosomal DNA internal transcribed spacers and chloroplast gene ndhF sequences. Plant Syst. Evol. 2003, 242, 1–32. [Google Scholar] [CrossRef]
- Kazempour Osaloo, S.; Maassoumi, A.A.; Murakami, N. Molecular systematics of the Old World Astragalus (Fabaceae) as inferred from nrDNA ITS sequence data. Brittonia 2005, 57, 367–381. [Google Scholar] [CrossRef]
- Maassoumi, A.A. The genus Astragalus in Iran; Research Institute of Forests and Rangelands: Tehran, Iran, 1989; Volume 2, p. 386. [Google Scholar]
- Maassoumi, A.A. Fabaceae: Astragalus III. Flora of Iran, No. 145.; Research Institute of Forests and Rangeland: Tehran, Iran, 2018; p. 766. [Google Scholar]
- Maassoumi, A.A. Astragalus of World. 2020. Available online: http://astragalusofworld.com (accessed on 20 July 2022).
- Podlech, D.; Zarre, S.; Maassoumi, A.A.; Ekici, M.; Sytin, A. Flora Iranica, Papilionaceae VI, Astragalus L. IV & Barnebyella Podlech; No. 178; Akademische Druck-u. Verlagsanstalt: Graz, Austria, 2010; p. 430. [Google Scholar]
- Podlech, D.; Maassoumi, A.A.; Zarre, S. Flora Iranica, Papilionaceae VII, Astragalus L. V.; No. 179; Akademische Druck-u. Verlagsanstalt: Graz, Austria, 2012; p. 312. [Google Scholar]
- Mahmoodi, M.; Maassoumi, A.A.; Noroozi, J. A new alpine species and a new record of Astragalus sect. Stereothrix (Fabaceae) from Iran, with comments on the phytogeography of the section. Willdenowia 2013, 43, 263–270. [Google Scholar]
- Ranjbar, M.; Mahmoudian, B.; Vitek, E. A synopsis of genus Astragalus L. sect. Stereothrix Bunge (Fabaceae). Biol. Diversity Conserv. 2013, 6, 114–133. [Google Scholar]
- Maassoumi, A.A.; Mahmoodi, M.; Bagheri, A. Astragalus sect. Irania (Fabaceae): A new monotypic section from Iran. Iran. J. Bot. 2020, 26, 100–110. [Google Scholar]
- Su, C.; Duan, L.; Liu, P.; Liu, J.; Chang, Z.; Wen, J. Chloroplast phylogenomics and character evolution of eastern Asian Astragalus (Leguminosae): Tackling the phylogenetic structure of the largest genus of flowering plants in Asia. Mol. Phylogenet. Evol. 2021, 156, 107025. [Google Scholar] [CrossRef] [PubMed]
- Thiers, B. Index Herbariorum: A Global Directory of Public Herbaria and Associated Staff. 2022. Available online: http://sweetgum.nybg.org/science/ih/ (accessed on 1 July 2022).
- Blattner, F.R. Direct amplification of the entire ITS region from poorly preserved plant material using recombinant PCR. Biotechniques 1999, 29, 1180–1186. [Google Scholar] [CrossRef] [PubMed]
- Riahi, M.; Zarre, S.; Maassoumi, A.A.; Wojciechowski, M.F. Towards a phylogeny for Astragalus section Caprini (Fabaceae) and its allies based on nuclear and plastid DNA sequences. Plant Syst. Evol. 2011, 293, 119–133. [Google Scholar] [CrossRef]
- Wojciechowski, M.F.; Lavin, M.; Sanderson, M.J. A phylogeny of legumes (Leguminosae) based on analysis of the plastid matK gene resolves many well-supported subclades within the family. Am. J. Bot. 2004, 91, 1846–1862. [Google Scholar] [CrossRef]
- Chromas. Available online: https://technelysium.com.au/wp/chromas/ (accessed on 10 July 2022).
- Swofford, D.L. PAUP*: Phylogenetic Analysis Using Parsimony (*and Other Methods); Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Blattner, F.R. Phylogeny of Hordeum (Poaceae) as inferred by nuclear rDNA ITS sequences. Mol. Phylogenet. Evol. 2004, 33, 289–299. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Farris, J.S.; Källersjö, M.; Kluge, A.G.; Bult, C. Testing significance of incongruence. Cladistics 1994, 10, 315–319. [Google Scholar] [CrossRef]
- Cunningham, C.W. Can three incongruence tests predict when data should be combined? Mol. Biol. Evol. 1997, 14, 733–740. [Google Scholar] [CrossRef]
- Lavin, M.; Herendeen, P.S.; Wojciechowski, M.F. Evolutionary rates analysis of Leguminosae implicates a rapid diversification of lineages during the Tertiary. Syst. Biol. 2005, 54, 575–594. [Google Scholar] [CrossRef]
- Bouckaert, R.R.; Heled, J.; Kühnert, D.; Vaughan, T.; Wu, C.-H.; Xie, D.; Suchard, M.A.; Rambaut, A.; Drummond, A.J. BEAST 2: A software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 2014, 10, e1003537. [Google Scholar] [CrossRef] [PubMed]
- Bouckaert, R.R.; Vaughan, T.G.; Barido-Sottani, J.; Duchêne, S.; Fourment, M.; Gavryushkina, A.; Heled, A.; Jones, G.; Kühnert, D.; De Maio, N.; et al. BEAST 2.5: An advanced software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 2019, 15, e1006650. [Google Scholar] [CrossRef] [PubMed]
- Bouckaert, R.R.; Drummond, A.J. bModelTest: Bayesian phylogenetic site model averaging and model comparison. BMC Evol. Biol. 2017, 17, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Drummond, A. Improving the performance of Bayesian phylogenetic inference under relaxed clock models. BMC Evol. Biol. 2020, 20, 54. [Google Scholar] [CrossRef] [PubMed]
- Douglas, J.; Zhang, R.; Bouckaert, R.R. Adaptive dating and fast proposals: Revisiting the phylogenetic relaxed clock model. PLoS Comput. Biol. 2021, 17, e1008322. [Google Scholar] [CrossRef]
- Drummond, A.J.; Ho, S.Y.W.; Phillips, M.J.; Rambaut, A. Relaxed phylogenetics and dating with confidence. PLoS Biol. 2006, 4, e88. [Google Scholar] [CrossRef]
- Heled, J.; Drummond, A.J. Calibrated tree priors for relaxed phylogenetics and divergence time estimation. Syst. Biol. 2012, 61, 138–149. [Google Scholar] [CrossRef]
- Hillis, D.H.; Bull, J.J. An empirical test of bootstrapping as a method for assessing confidence in phylogenetic analysis. Syst. Biol. 1993, 42, 182–192. [Google Scholar] [CrossRef]
- Ranjbar, M.; Karamian, R. Astragalus sect. Astragalus (Fabaceae) in Iran, complementary notes with a key to the species. Nord. J. Bot. 2002, 22, 177–182. [Google Scholar] [CrossRef]
- Maassoumi, A.A.; Kazempour Osaloo, S.; Sotoodeh, A. A new infrageneric grouping of Astragalus (Fabaceae). Iran. J. Bot. 2016, 22, 88–100. [Google Scholar]
- Maassoumi, A.A. Old World Check-List of Astragalus, 1st ed.; Research Institute of Forests and Rangeland: Tehran, Iran, 1998; p. 617. [Google Scholar]
- Zarre, S.; Podlech, D.; Taeb, F. New species of the genus Astragalus L. (Fabaceae), mainly from the “Flora Iranica” area. Feddes Repert. 2005, 116, 54–79. [Google Scholar] [CrossRef]
- Bagheri, A.; Maassoumi, A.A.; Vitek, E. Rediscovery of Astragalus saganlugensis (Fabaceae, Galegeae) in Iran after 184 years. Phytotaxa 2018, 350, 297–299. [Google Scholar] [CrossRef]
- Sirjaev, G.; Rechinger, K.H. Astragali novi iranici I. (Rechingeri iter iranicum secundum No. 28.). Österr. Akad. Wiss. Math.-Nat. KI. Anz. 1953, 90, 114–125. [Google Scholar]
- Raca, I.; Blattner, F.R.; Waminal, N.E.; Kerndorff, H.; Ranđelović, V.; Harpke, D. Disentangling a polyploid complex within Crocus series. Verni. Biology, 2023; in press. [Google Scholar]



{kind=link}
{kind=link}
| Species | Podlech et al., 2010 [13] | Podlech et al., 2012 [14] | Podlech and Zarre 2013 [1] | Maassoumi 2018 [11] | Maassoumi 2020 [12] | Current Study |
|---|---|---|---|---|---|---|
| A. altimontanus Podlech and Maassoumi | Stereothrix | Stereothrix | Stereothrix | Stereothrix | Stereothrix | Hypoglottidei |
| A. andabilensis Ranjbar and Mahmoudian | - | - | - | - | Stereothrix | Stereothrix |
| A. atricapillus Bornm. | Hypoglottidei | Hypoglottidei | Hypoglottidei | Hypoglottidei | Hypoglottidei | Hypoglottidei |
| A. badelehensis Maassoumi and Taheri | - | - | - | Stereothrix | Stereothrix | Stereothrix |
| A. barbatus Lam. | Stereothrix | - | Stereothrix | - | Stereothrix | Stereothrix |
| A. bavanatensis Zarre and Podlech | Stereothrix | Stereothrix | Stereothrix | Stereothrix | Stereothrix | Stereothrix |
| A. bojnurdensis Podlech | Brachylobium | Brachylobium | Brachylobium | Hypoglottidei | Hypoglottidei | Hypoglottidei |
| A. brachypetalus Trautv. | Hypoglottidei | Hypoglottidei | Stereothrix | Hypoglottidei | Hypoglottidei | Hypoglottidei |
| A. capito Boiss. and Hohen. | Stereothrix | Stereothrix | Stereothrix | Stereothrix | Stereothrix | Stereothrix |
| A. chamberlainianus H.Sümbül | - | - | Stereothrix | - | - | Stereothrix |
| A. daenensis Boiss. | Brachylobium | Brachylobium | Brachylobium | Brachylobium | Brachylobium | Brachylobium |
| A. damghanensis Podlech | Stereothrix | Stereothrix | Stereothrix | Hypoglottidei | Stereothrix | Hypoglottidei |
| A. doshman-ziariensis Maassoumi and Podlech | Stereothrix | Stereothrix | Stereothrix | Stereothrix | Koelziana | Stereothrix |
| A. hakkariensis Podlech | Stereothrix | - | - | Stereothrix | Stereothrix | Stereothrix |
| A. herbertii Maassoumi | Hypoglottidei | Hypoglottidei | Hypoglottidei | Hypoglottidei | Hypoglottidei | Hypoglottidei |
| A. inexpectatus Maassoumi and Podlech | Malacothrix | Malacothrix | Malacothrix | Malacothrix/ Plagiophaca | Plagiophaca | Malacothrix |
| A. koelzii Barneby | Stereothrix | Stereothrix | Stereothrix | Koelziana | Koelziana | Koelziana |
| A. ledinghamii Barneby | Stereothrix | Stereothrix | Stereothrix | Stereothrix | Koelziana | Stereothrix |
| A. leucothrix Freyn and Bornm. | - | - | Stereothrix | - | Stereothrix | Stereothrix |
| A. longirostratus Pau (=A. perpexus Maassoumi) | Hhypoglottidei | Hemiphaca | Oroboidei | Hypoglottidei | Hypoglottidei | Hypoglottidei |
| A. mahmutlarensis Podlech | - | - | Stereothrix | - | Stereothrix | Stereothrix |
| A. mahneshanensis Maassoumi and Moussavi | - | Stereothrix | Stereothrix | Stereothrix | Stereothrix | Stereothrix |
| A. montis-alamkuhi Maassoumi | - | - | - | Stereothrix | Stereothrix | Stereothrix |
| A. montismishoudaghi Sheikh Akbari Mehr, Ghorbani and Maassoumi | - | - | - | Stereothrix | Stereothrix | Stereothrix |
| A. montis-varvashti Podlech | Stereothrix | Stereothrix | Stereothrix | Stereothrix | Stereothrix | Stereothrix |
| A. nezva-montis Podlech and Zarre | Hypoglottidei | Hypoglottidei | Hypoglottidei | Plagiophaca | Plagiophaca | Malacothrix |
| A. nurensis Boiss. and Buhse (=A. pish-chakensis Maassoumi) | Hypoglottidei | Hypoglottidei | Hypoglottidei | Hypoglottidei | Hypoglottidei | Hypoglottidei |
| A. penetratus Maassoumi | Brachylobium | Brachylobium | Brachylobium | Brachylobium | Brachylobium | Brachylobium |
| A. plagiophacos Maassoumi and Podlech | Plagiophaca | Plagiophaca | Plagiophaca | Plagiophaca | Plagiophaca | Plagiophaca |
| A. podosphaerus Boiss. and Hausskn. | Stereothrix | Stereothrix | Stereothrix | Stereothrix | Stereothrix | Stereothrix |
| A. pseudocapito Podlech | Stereothrix | Stereothrix | Stereothrix | Stereothrix | Stereothrix | Stereothrix |
| A. rimarum Bornm. | Hypoglottidei | Hypoglottidei | Hypoglottidei | Hypoglottidei | Hypoglottidei | Hypoglottidei |
| A. saganlugensis Trautv. | Hypoglottidei | Hypoglottidei | Hypoglottidei | Hypoglottidei | Hypoglottidei | Hypoglottidei |
| A. setosulus Gontsch. | - | - | Stereothrix | - | Stereothrix | Stereothrix |
| A. sphaeranthus Boiss. | Stereothrix | Stereothrix | Stereothrix | Stereothrix | Stereothrix | Stereothrix |
| A. yazdii (Vassilcz.) Podlech and Maassoumi | Brachylobium | Brachylobium | Brachylobium | Brachylobium | Brachylobium | Brachylobium |
| Section | Established by | Year |
|---|---|---|
| Hypoglottidei | DC. | 1825 |
| Oroboidei | A.Gray | 1864 |
| Stereothri | Bunge | 1868 |
| Malacothrix | Bunge | 1868 |
| Hemiphaca | Bunge | 1868 |
| Brachylobium | Boiss. | 1872 |
| Koelziana | Širj. and Rech.f. | 1953 |
| Plagiophaca | Maassoumi and Podlech | 1989 |
| ETS | ITS | ETS + ITS | matK | ETS + ITS + matK | |
|---|---|---|---|---|---|
| Alignment lengths | 282 | 618 | 900 | 1153 | 2053 |
| Constant characteristics | 252 | 459 | 711 | 1044 | 1755 |
| Variable characteristics | 30 | 159 | 189 | 109 | 298 |
| Parsimony-informative characteristics | 7 | 99 | 106 | 61 | 167 |
| MP tree lengths | 208 | 279 | 318 | 150 | 711 |
| Consistency index (CI) | 0.78 | 0.75 | 0.75 | 0.76 | 0.76 |
| Retention index (RI) | 0.82 | 0.86 | 0.86 | 0.85 | 0.86 |
| Sequence evolution model (No. of categories) | F81(1) | SYM + Γ(6) | TrN + I + Γ(6) | TVM + I + Γ(6) | TVM + I + Γ(6) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bagheri, A.; Maassoumi, A.A.; Brassac, J.; Blattner, F.R. Dated Phylogeny of Astragalus Section Stereothrix (Fabaceae) and Allied Taxa in the Hypoglottis Clade. Biology 2023, 12, 138. https://doi.org/10.3390/biology12010138
Bagheri A, Maassoumi AA, Brassac J, Blattner FR. Dated Phylogeny of Astragalus Section Stereothrix (Fabaceae) and Allied Taxa in the Hypoglottis Clade. Biology. 2023; 12(1):138. https://doi.org/10.3390/biology12010138
Chicago/Turabian StyleBagheri, Ali, Ali Asghar Maassoumi, Jonathan Brassac, and Frank R. Blattner. 2023. "Dated Phylogeny of Astragalus Section Stereothrix (Fabaceae) and Allied Taxa in the Hypoglottis Clade" Biology 12, no. 1: 138. https://doi.org/10.3390/biology12010138
APA StyleBagheri, A., Maassoumi, A. A., Brassac, J., & Blattner, F. R. (2023). Dated Phylogeny of Astragalus Section Stereothrix (Fabaceae) and Allied Taxa in the Hypoglottis Clade. Biology, 12(1), 138. https://doi.org/10.3390/biology12010138