
Foliar Roughness and Water Content Impact on Escherichia coli Attachment in Baby Leafy Greens
 , , , , and
, , , , and 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Baby Leaves Cultivation and Characterization
2.2. Bacterial Strain
2.3. Surface Inoculation of the Baby Leaves
2.4. Analysis of Leaf Micro-Morphological Traits
2.4.1. Stomata
2.4.2. Roughness
2.5. UV Treatment
2.6. Statistical Analysis
3. Results
3.1. E. coli Attachment in the Baby Leaves
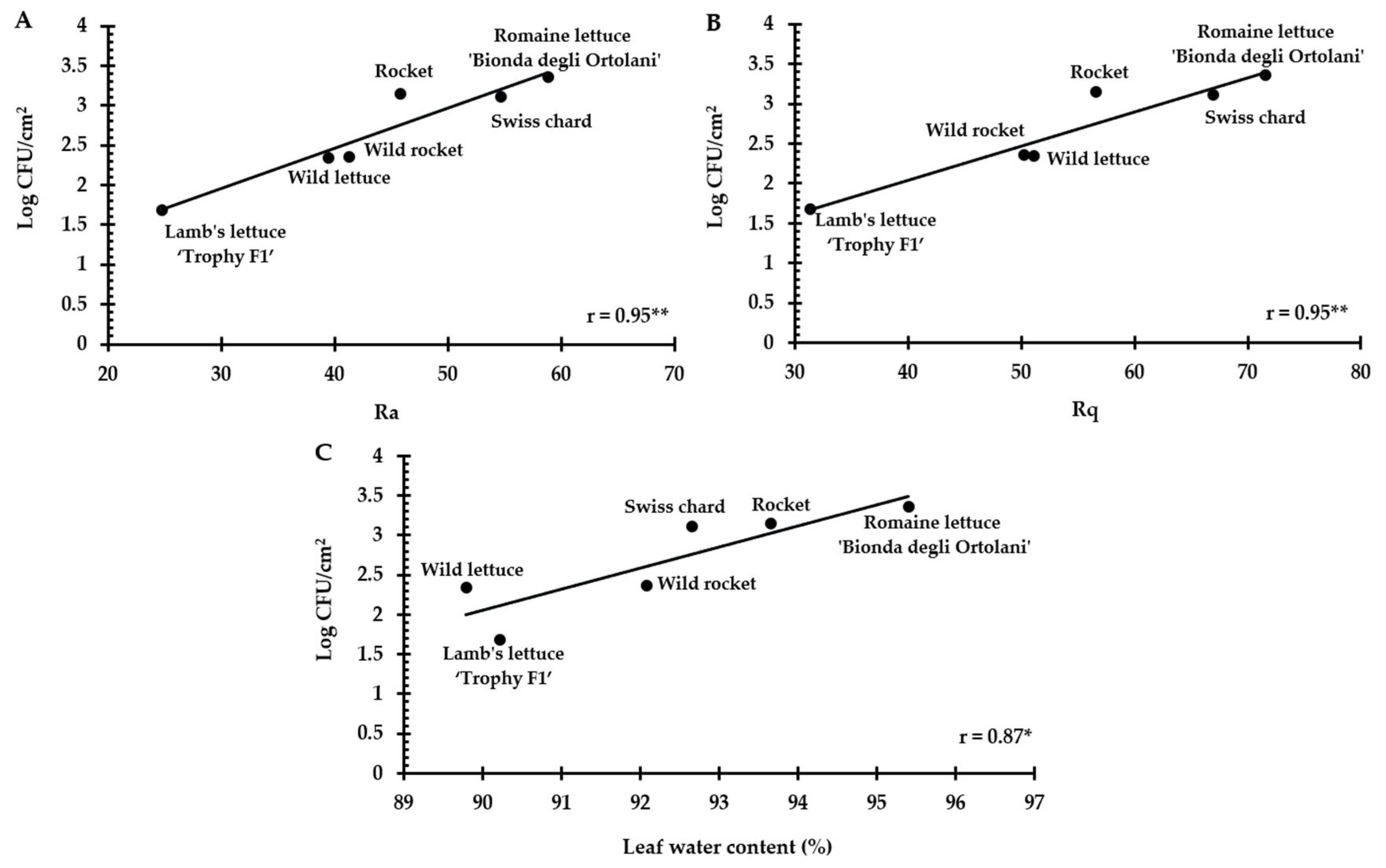
3.2. Leaf Micro-Morphological Traits and Water Content
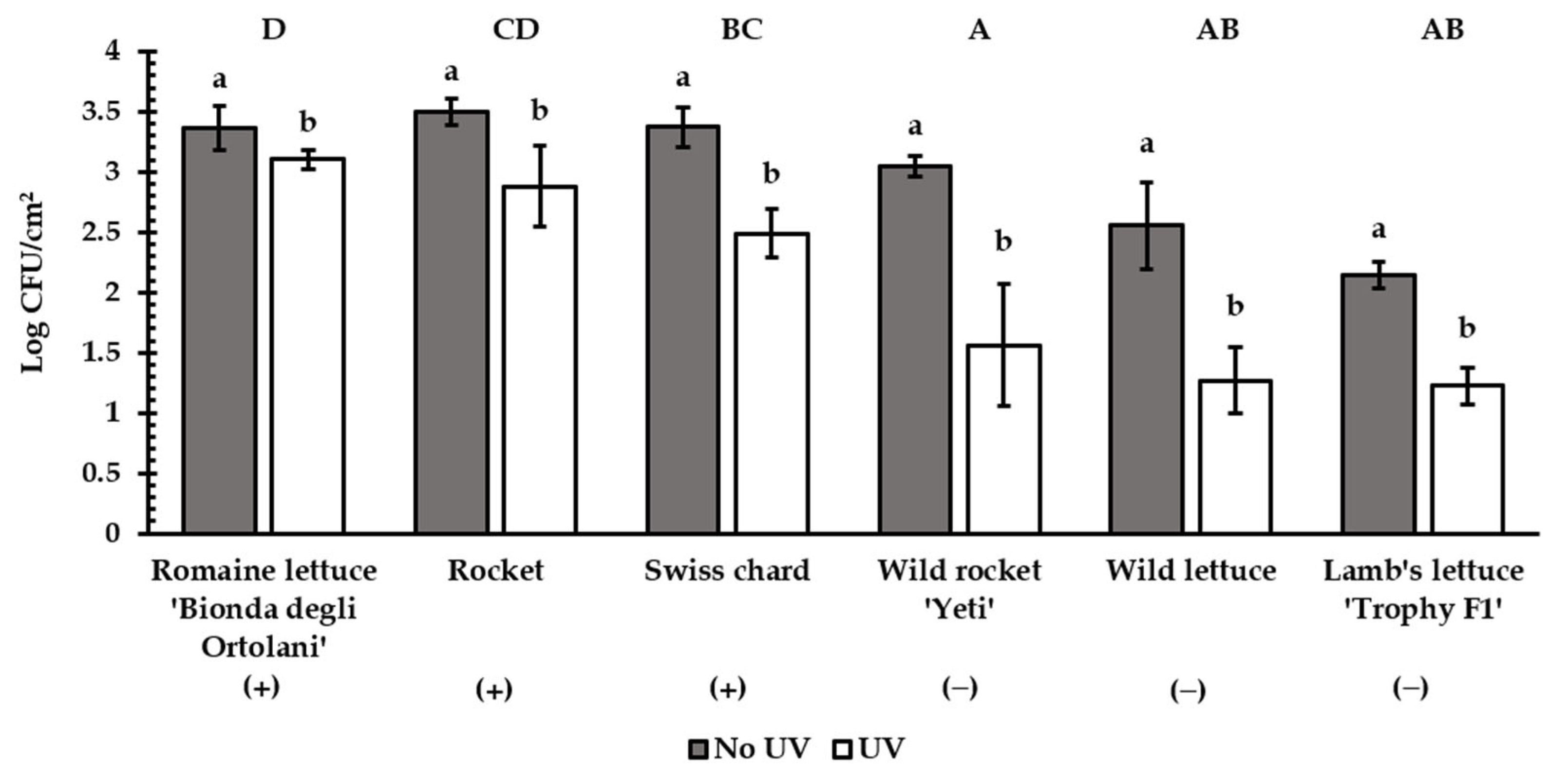
3.3. UV Experiment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Di Gioia, F.; Renna, M.; Santamaria, P. Sprouts, microgreens and “baby leaf” vegetables. In Minimally Processed Refrigerated Fruits and Vegetables; Springer: Boston, MA, USA, 2017; pp. 403–432. [Google Scholar] [CrossRef]
- Sucheta Singla, G.; Chaturvedi, K.; Sandhu, P.P. Status and recent trends in fresh-cut fruits and vegetables. In Fresh-Cut Fruits and Vegetables; Siddiqui, M.W., Ed.; Academic Press: Cambridge, MA, USA, 2020; pp. 17–49. [Google Scholar] [CrossRef]
- Grand View Research. Packaged Salad Market Size, Share & Trends Analysis Report by Product (Vegetarian, Non-Vegetarian), by Processing (Organic, Conventional), by Type, by Distribution Channel, by Region, and Segment Forecasts, 2021–2028. San Francisco (CA). 2021. Available online: https://www.grandviewresearch.com/industry-analysis/packaged-salad-market (accessed on 30 December 2022).
- Lenzi, A.; Orlandini, A.; Bulgari, R.; Ferrante, A.; Bruschi, P. Antioxidant and mineral composition of three wild leafy species: A comparison between microgreens and baby greens. Foods 2019, 8, 487. [Google Scholar] [CrossRef] [PubMed]
- Bartz, J.A.; Marvasi, M.; Teplitski, M. Salmonella and tomatoes. In The Produce Contamination Problem: Causes and Solution; Matthews, K.R., Sapers, G.M., Gerba, C.P., Eds.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 269–289. [Google Scholar] [CrossRef]
- Mir, S.A.; Shah, M.A.; Mir, M.M.; Dar, B.; Greiner, R.; Roohinejad, S. Microbiological contamination of ready-to-eat vegetable salads in developing countries and potential solutions in the supply chain to control microbial pathogens. Food Control 2018, 85, 235–244. [Google Scholar] [CrossRef]
- Ölmez, H. Foodborne pathogenic bacteria in fresh-cut vegetables and fruits. In Food Hygiene and Toxicology in Ready-to-Eat Foods; Kotzekidou, P., Ed.; Academic Press Books, Elsevier: Amsterdam, The Netherlands, 2016; pp. 151–166. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. E. coli Outbreak Linked to Packaged Salads. 2022. Available online: https://www.cdc.gov/ecoli/2021/o157h7-12-21/index.html (accessed on 15 September 2022).
- Dunne, W.M. Bacterial adhesion: Seen any good biofilms lately? Clin. Microbiol. Rev. 2002, 15, 155–166. [Google Scholar] [CrossRef] [PubMed]
- Grivokostopoulos, N.C.; Makariti, I.P.; Hilaj, N.; Apostolidou, Z.; Skandamis, P.N. Internalization of Salmonella in leafy greens and the impact on acid tolerance. Appl. Environ. Microbiol. 2022, 88, e02249-21. [Google Scholar] [CrossRef] [PubMed]
- Macarisin, D.; Patel, J.; Bauchan, G.; Giron, J.A.; Ravishankar, S. Effect of spinach cultivar and bacterial adherence factors on survival of Escherichia coli O157:H7 on spinach leaves. J. Food Prot. 2013, 76, 1829–1837. [Google Scholar] [CrossRef]
- Sela, S.; Manulis-Sasson, S. What else can we do to mitigate contamination of fresh produce by foodborne pathogens? Microb. Biotechnol. 2015, 8, 29–31. [Google Scholar] [CrossRef]
- Luna-Guevara, J.J.; Arenas-Hernandez, M.M.; Martínez de la Peña, C.; Silva, J.L.; Luna-Guevara, M.L. The role of pathogenic E. coli in fresh vegetables: Behavior, contamination factors, and preventive measures. Int. J. Microb. 2019, 2019, 2894328. [Google Scholar] [CrossRef]
- Doan, H.K.; Antequera-Gómez, M.L.; Parikh, A.N.; Leveau, J.H.J. Leaf surface topography contributes to the ability of Escherichia coli on leafy greens to resist removal by washing, escape disinfection with chlorine, and disperse through splash. Front. Microbiol. 2020, 11, 1485. [Google Scholar] [CrossRef]
- Doan, H.K.; Ngassam, V.N.; Gilmore, S.F.; Tecon, R.; Parikh, A.N.; Leveau, J.H.J. Topography-driven shape, spread, and retention of leaf surface water impacts microbial dispersion and activity in the phyllosphere. Phytobiomes J. 2020, 4, 268–280. [Google Scholar] [CrossRef]
- Melotto, M.; Underwood, W.; He, S.Y. Role of stomata in plant innate immunity and foliar bacterial diseases. Annu. Rev. Phytopathol. 2008, 46, 101–122. [Google Scholar] [CrossRef]
- Yaron, S.; Römling, U. Biofilm formation by enteric pathogens and its role in plant colonization and persistence. Microb. Biotechnol. 2014, 7, 496–516. [Google Scholar] [CrossRef]
- Kroupitski, Y.; Pinto, R.; Brandl, M.T.; Belausov, E.; Sela, S. Interactions of Salmonella enterica with lettuce leaves. J. Appl. Microbiol. 2009, 106, 1876–1885. [Google Scholar] [CrossRef]
- Saldana, Z.; Sánchez, E.; Xicohtencatl-Cortes, J.; Puente, J.L.; Girón, J.A. Surface structure involved in plant stomata and leaf colonization by Shiga-toxigenic Escherichia coli O157:H7. Front. Microbiol. 2011, 2, 119. [Google Scholar] [CrossRef]
- Wang, H.; Zhou, B.; Feng, H. Surface characteristics of fresh produce and their impact on attachment and removal of human pathogens on produce surfaces. In Decontamination of Fresh and Minimally Processed Produce, 1st ed.; Gómez-López, V.M., Ed.; John Wiley & Sons, Inc.: Ames, IA, USA, 2012; pp. 43–57. [Google Scholar]
- Melotto, M.; Panchal, S.; Roy, D. Plant innate immunity against human bacterial pathogens. Front. Microbiol. 2014, 5, 411. [Google Scholar] [CrossRef]
- Mitra, R.; Cuesta-Alonso, E.; Wayadande, A.C.; Talley, J.; Gilliland, S.; Fletcher, J. Effect of route of introduction and host cultivar on the colonization, internalization, and movement of the human pathogen Escherichia coli O157:H7 in spinach. J. Food Prot. 2009, 72, 1521–1530. [Google Scholar] [CrossRef]
- Jacob, C.; Melotto, M. Human pathogen colonization of lettuce dependent upon plant genotype and defense response activation. Front. Plant Sci. 2020, 10, 1769. [Google Scholar] [CrossRef]
- Palma-Salgado, S.; Ku, K.-M.; Dong, M.; Nguyen, T.H.; Juvik, J.A.; Feng, H. Adhesion and removal of E. coli K12 as affected by leafy green produce epicuticular wax composition, surface roughness, produce and bacterial surface hydrophobicity, and sanitizers. Int. J. Food Microbiol. 2020, 334, 108834. [Google Scholar] [CrossRef]
- Yadav, R.K.P.; Karamanoli, K.; Vokou, D. Bacterial colonization of the phyllosphere of Mediterranean perennial species as influenced by leaf structural and chemical features. Microb. Ecol. 2005, 50, 185–196. [Google Scholar] [CrossRef]
- Chapman, T.A.; Wu, X.Y.; Barchia, I.; Bettelheim, K.A.; Driesen, S.; Trott, D.; Wilson, M.; Chin, J.J.C. Comparison of virulence gene profiles of Escherichia coli strains isolated from healthy and diarrheic swine. Appl. Environ. Microbiol. 2006, 72, 4782–4795. [Google Scholar] [CrossRef]
- Marvasi, M.; Noel, J.T.; George, A.S.; Farias, M.A.; Jenkins, K.T.; Hochmuth, G.; Xu, Y.; Giovanonni, J.J.; Teplitski, M. Ethylene signalling affects susceptibility of tomatoes to Salmonella. Microb. Biotechnol. 2014, 7, 545–555. [Google Scholar] [CrossRef]
- Pathan, A.; Bond, J.; Gaskin, R. Sample preparation for SEM of plant surfaces. Mater. Today 2010, 12, 32–43. [Google Scholar] [CrossRef]
- Rasband, W.S. ImageJ; U.S. National Institutes of Health: Bethesda, MD, USA, 1997–2016. Available online: https://imagej.nih.gov/ij/ (accessed on 16 September 2022).
- Cacciari, I.; Ciofini, D.; Mascalchi, M.; Mencaglia, A.; Siano, S. Novel approach to the microscopic inspection during laser cleaning treatments of artworks. Anal. Bioanal. Chem. 2012, 402, 1585–1591. [Google Scholar] [CrossRef] [PubMed]
- Siano, S.; Mencaglia, A.A.; Cacciari, I. Microscopy Optoelectric Device with Focus Scanning. U.S. Patent 9,612,427 B, 4 April 2017. [Google Scholar]
- Cacciari, I.; Mencaglia, A.A.; Siano, S. Micromorphology of gold jewels: A novel algorithm for 3D reconstruction and its quality assessment. In Optics for Arts, Architecture, and Archaeology IV, Proceedings of the SPIE Optical Metrology, Munich, Germany, 30 May 2013; SPIE: Washington, DC, USA, 2013; Volume 8790B, pp. 70–80. [Google Scholar] [CrossRef]
- ISO 25178-2:2021; Standard—Geometrical Product Specifications (GPS)—Surface Texture: Areal—Part 2: Terms, Definitions and Surface Texture Parameters. ISO: Geneva, Switzerland, 2021.
- Lenzi, A.; Marvasi, M.; Baldi, A. Agronomic practices to limit pre-and post-harvest contamination and proliferation of human pathogenic Enterobacteriaceae in vegetable produce. Food Control 2020, 119, 107486. [Google Scholar] [CrossRef]
- Slayton, R.; Turabelidze, G.; Bennett, S.; Schwensohn, C.; Yaffee, A.; Khan, F.; Butler, C.; Trees, E.; Ayers, T.; Davis, M.; et al. Outbreak of Shiga toxin-producing Escherichia coli (STEC) O157:H7 associated with romaine lettuce consumption, 2011. PLoS ONE 2013, 8, e55300. [Google Scholar] [CrossRef] [PubMed]
- Taylor, E.V.; Nguyen, T.A.; Machesky, K.D.; Koch, E.; Sotir, M.J.; Bohm, S.R.; Folster, P.; Bokanyi, R.; Kupper, A.; Bidol, S.A.; et al. Multistate outbreak of Escherichia coli O145 infections associated with romaine lettuce consumption, 2010. J. Food Prot. 2013, 76, 939–944. [Google Scholar] [CrossRef]
- Coulombe, G.; Catford, A.; Martinez-Perez, A.; Buenaventura, E. Outbreaks of Escherichia coli O157:H7 infections linked to romaine lettuce in Canada from 2008 to 2018: An Analysis of Food Safety Context. J. Food Prot. 2020, 83, 1444–1462. [Google Scholar] [CrossRef]
- Waltenburg, M.A.; Schwensohn, C.; Madad, A.; Seelman, S.L.; Peralta, V.; Koske, S.E.; Boyle, M.M.; Arends, K.; Patel, K.; Mattioli, M.; et al. Two multistate outbreaks of a reoccurring Shiga toxin-producing Escherichia coli strain associated with romaine lettuce: USA, 2018–2019. Epidemiol. Infect. 2022, 150, E16. [Google Scholar] [CrossRef]
- Lenzi, A.; Baldi, A.; Lombardelli, L.; Truschi, S.; Marvasi, M.; Bruschi, P. Contamination of microgreens by Salmonella enterica and Escherichia coli is influenced by selection breeding in chicory (Cichorium intybus L.). Food Qual. Saf. 2022, 6, fyac030. [Google Scholar] [CrossRef]
- Hunter, P.J.; Shaw, R.K.; Berger, C.N.; Frankel, G.; Pink, D.; Hand, P. Older leaves of lettuce (Lactuca spp.) support higher levels of Salmonella enterica ser. Senftenberg attachment and show greater variation between plant accessions than do younger leaves. FEMS Microbiol. Lett. 2015, 362, fnv077. [Google Scholar] [CrossRef]
- Gomes, C.; Da Silva, P.; Moreira, R.G.; Castell-Perez, E.; Ellis, E.A.; Pendleton, M. Understanding E. coli internalization in lettuce leaves for optimization of irradiation treatment. Int. J. Food Microbiol. 2009, 135, 238–247. [Google Scholar] [CrossRef]
- Golberg, D.; Kroupitski, Y.; Belausov, E.; Pinto, R.; Sela, S. Salmonella Typhimurium internalization is variable in leafy vegetables and fresh herbs. Int. J. Food Microbiol. 2011, 145, 250–257. [Google Scholar] [CrossRef]
- Brandl, M.T.; Cox, C.E.; Teplitski, M. Salmonella interactions with plants and their associated microbiota. Phytopathology 2013, 103, 316–325. [Google Scholar] [CrossRef]
- Ku, K.-M.; Chiu, Y.-C.; Shen, C.; Jenks, M. Leaf cuticular waxes of lettuce are associated with reduced attachment of the foodborne pathogen Salmonella spp. at harvest and after postharvest storage. LWT 2020, 117, 108657. [Google Scholar] [CrossRef]
- Yi, J.; Leveau, J.H.; Nitin, N. Role of multiscale leaf surface topography in antimicrobial efficacy of chlorine-based sanitizers. J. Food Eng. 2022, 332, 111118. [Google Scholar] [CrossRef]
- Wang, H.; Feng, H.; Liang, W.; Luo, Y.; Malyarchuk, V. Effect of surface roughness on retention and removal of Escherichia coli O157:H7 on surfaces of selected fruits. J. Food Sci. 2009, 74, E8–E15. [Google Scholar] [CrossRef]
- Fransisca, L.; Feng, H. Effect of surface roughness on inactivation of Escherichia coli O157:H7 87-23 by new organic acid–surfactant combinations on alfalfa, broccoli, and radish seeds. J. Food Prot. 2012, 75, 261–269. [Google Scholar] [CrossRef]
- Singh, H.; Bhardwaj, S.K.; Khatri, M.; Kim, K.-H.; Bhardwaj, N. UVC radiation for food safety: An emerging technology for the microbial disinfection of food products: A Review. Chem. Eng. J. 2021, 417, 128084. [Google Scholar] [CrossRef]
- Chun, H.H.; Kim, J.Y.; Song, K.B. Inactivation of foodborne pathogens in ready-to-eat salad using UV-C irradiation. Food Sci. Biotechnol. 2010, 19, 547–551. [Google Scholar] [CrossRef]
- Kim, Y.H.; Jeong, S.G.; Back, K.H.; Park, K.H.; Chung, M.S.; Kang, D.H. Effect of various conditions on inactivation of Escherichia coli O157:H7, Salmonella typhimurium, and Listeria monocytogenes in fresh-cut lettuce using ultraviolet radiation. Int. J. Food Microbiol. 2013, 166, 349–355. [Google Scholar] [CrossRef]
- Woodling, S.E.; Moraru, C.I. Influence of surface topography on the effectiveness of pulsed light treatment for the inactivation of Listeria innocua on stainless-steel surfaces. J. Food Sci. 2005, 70, m345–m351. [Google Scholar] [CrossRef]
- Marvasi, M.; Hochmuth, G.J.; Giurcanu, M.C.; George, A.S.; Noel, J.T.; Bartz, J.; Teplitski, M. Factors that affect proliferation of Salmonella in tomatoes post-harvest: The roles of seasonal effects, irrigation regime, crop and pathogen genotype. PLoS ONE 2013, 8, e80871. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Specie | Variety/Type/Cultivar | Accession Name | |
|---|---|---|---|---|
| 1 | Asteraceae | Cichorium intybus L. | var. sativus Witloof type | Witloof chicory |
| 2 | var. foliosum cv. ‘Magdeburgo’ | Chicory ‘Magdeburgo’ | ||
| 3 | - | Wild chicory Ingegnoli | ||
| 4 | - | Wild chicory B&T | ||
| 5 | - | Wild chicory local | ||
| 6 | var. foliosum cv. ‘Biondissima di Trieste’ | Chicory ‘Biondissima di Trieste’ | ||
| 7 | var. foliosum cv. ‘Spadona da taglio’ | Chicory ‘Spadona da taglio’ | ||
| 8 | Cichorium endivia L. | var. crispum | Endive | |
| 9 | Lactuca sativa L. | var. crispa Lollo verde type | Lollo verde lettuce | |
| 10 | var. crispa Lollo rossa type | Lollo rossa lettuce | ||
| 11 | var. crispa blonde type | Blonde lettuce | ||
| 12 | var. crispa cv. ‘Pamela’ | Lettuce ‘Pamela’ | ||
| 13 | var. longifolia cv. ‘Bionda degli ortolani’ | Romaine lettuce ‘Bionda degli Ortolani’ | ||
| 14 | var. longifolia cv. ‘Maraichere’ | Romaine lettuce ‘Maraichere’ | ||
| 15 | Lactuca serriola L. | - | Wild lettuce | |
| 16 | Taraxacum campylodes G.E.Haglund | - | Dandelion local | |
| 17 | - | Dandelion Ingegnoli | ||
| 18 | Brassicaceae | Eruca sativa Miller | - | Rocket |
| 19 | Diplotaxis tenuifolia L. | - | Wild rocket Ingegnoli | |
| 20 | cv. ‘Yeti’ | Wild rocket ‘Yeti’ | ||
| 21 | Brassica rapa L. | subsp. chinensis | Pak choi | |
| 22 | subsp. nipposinica | Mizuna | ||
| 23 | Brassica juncea L. | - | Wasabina leaf mustard | |
| 24 | - | Red Giant leaf mustard | ||
| 25 | - | Red leaf mustard | ||
| 26 | Chenopodiaceae | Beta vulgaris L. | subsp. cycla | Swiss chard |
| 27 | subsp. cycla cv.‘Bull’s Blood Artica’ | Red chard ‘Bull’s Blood Artica’ | ||
| 28 | Spinacia oleracea L. | cv. ‘Cugoe RZ F1′ | Spinach ‘Cugoe RZ F1′ | |
| 29 | Valerianeceae | Valerianella locusta L. | cv. ‘Trophy F1′ | Lamb’s lettuce ‘Trophy F1′ |
| 30 | Polygonaceae | Rumex acetosa L. | - | Sorrel |
| Accession | Attachment log CFU/cm2 | Accession | Attachment log CFU/cm2 |
|---|---|---|---|
| Romaine lettuce ‘Bionda degli Ortolani’ | 3.36 ± 0.36 a | Dandelion (local) | 2.94 ± 0.32 abcd |
| Rocket | 3.15 ± 0.14 ab | Chicory ‘Biondissima di Trieste’ | 2.93 ± 0.31 abcd |
| Swiss chard | 3.11 ± 0.20 ab | Pak-choi | 2.93 ± 0.24 abcd |
| Endive | 3.11 ± 0.26 ab | Lollo verde lettuce | 2.90 ± 0.30 abcd |
| Spinach ‘Cugoe RZ F1′ | 3.10 ± 0.23 ab | Blonde lettuce | 2.87 ± 0.21 abcde |
| Mizuna | 3.10 ± 0.20 ab | Red leaf mustard | 2.83 ± 0.27 abcde |
| Wild chicory (B&T) | 3.03 ± 0.43 abc | Wild chicory (Ingegnoli) | 2.81 ± 0.36 abcde |
| Dandelion (Ingegnoli) | 3.02 ± 0.46 abc | Red chard ‘Bull’s Blood Artica’ | 2.79 ± 0.27 abcde |
| Red Giant leaf mustard | 3.01 ± 0.25 abc | Wild chicory (local) | 2.68 ± 0.55 bcde |
| Wasabina leaf mustard | 3.00 ± 0.48 abc | Romaine lettuce ‘Maraichere’ | 2.65 ± 0.34 bcde |
| Wild rocket (Ingegnoli) | 2.98 ± 0.32 abcd | Lettuce ‘Pamela’ | 2.45 ± 0.20 cde |
| Chicory ‘Magdeburgo’ | 2.97 ± 0.46 abcd | Sorrel | 2.40 ± 0.69 de |
| Lollo rossa lettuce | 2.96 ± 0.50 abcd | Wild rocket ‘Yeti’ | 2.36 ± 0.52 e |
| Chicory ‘Spadona da taglio’ | 2.96 ± 0.21 abcd | Wild lettuce | 2.34 ± 0.53 e |
| Witloof Chicory | 2.95 ± 0.46 abcd | Lamb’s lettuce ‘Trophy F1′ | 1.68 ± 0.39 f |
| Stomatal Density (n/mm2) | Stomata Length (µm) | Stomata Width (µm) | Stomatal Rim Area (µm2) | Ra | Rq | Water Content (%) |
|---|---|---|---|---|---|---|
| Romaine lettuce ‘Bionda degli Ortolani’ (+) | ||||||
| 71.96 ± 6.11 ab | 25.93 ± 1.96 b | 20.56 ± 3.28 c | 23.03 ± 5.52 bc | 58.81 ± 11.52 a | 71.56 ± 13.71 a | 95.36 ± 0.84 a |
| Rocket (+) | ||||||
| 92.20 ± 28.40 a | 23.46 ± 1.39 c | 15.66 ± 1.92 d | 11.70 ± 3.55 c | 45.81 ± 7.74 bc | 56.58 ± 9.79 bc | 93.66 ± 1.59 b |
| Swiss chard (+) | ||||||
| 61.65 ± 10.80 b | 30.10 ± 1.98 a | 21.85±3.38 b | 45.58±6.78 a | 54.70 ± 15.84 ab | 66.92 ± 18.35 ab | 92.66 ± 1.62 b |
| Wild rocket ‘Yeti’ (−) | ||||||
| 80.29 ± 25.67 ab | 21.85 ± 1.64 d | 14.38±2.23 e | 24.35±7.63 b | 41.33 ± 4.01 c | 50.23 ± 5.01 c | 92.08 ± 1.03 bc |
| Wild lettuce (−) | ||||||
| - | - | - | - | 39.43 ± 12.42 c | 51.14 ± 15.49 c | 89.79 ± 1.13 c |
| Lamb’s lettuce ‘Trophy F1′ (−) | ||||||
| 57.14 ± 5.47 b | 30.19 ± 1.33 a | 23.83 ± 2.05 a | 19.84 ± 1.52 bc | 24.74 ± 10.92 d | 31.38 ± 14.24 d | 90.22 ± 2.30 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Truschi, S.; Baldi, A.; Bruschi, P.; Cacciari, I.; Marvasi, M.; Lenzi, A. Foliar Roughness and Water Content Impact on Escherichia coli Attachment in Baby Leafy Greens. Biology 2023, 12, 102. https://doi.org/10.3390/biology12010102
Truschi S, Baldi A, Bruschi P, Cacciari I, Marvasi M, Lenzi A. Foliar Roughness and Water Content Impact on Escherichia coli Attachment in Baby Leafy Greens. Biology. 2023; 12(1):102. https://doi.org/10.3390/biology12010102
Chicago/Turabian StyleTruschi, Stefania, Ada Baldi, Piero Bruschi, Ilaria Cacciari, Massimiliano Marvasi, and Anna Lenzi. 2023. "Foliar Roughness and Water Content Impact on Escherichia coli Attachment in Baby Leafy Greens" Biology 12, no. 1: 102. https://doi.org/10.3390/biology12010102
APA StyleTruschi, S., Baldi, A., Bruschi, P., Cacciari, I., Marvasi, M., & Lenzi, A. (2023). Foliar Roughness and Water Content Impact on Escherichia coli Attachment in Baby Leafy Greens. Biology, 12(1), 102. https://doi.org/10.3390/biology12010102