



The Role of Two Linear β-Glucans Activated by c-di-GMP in Rhizobium etli CFN42
 , , , and
, , , and
Abstract
Simple Summary
Abstract
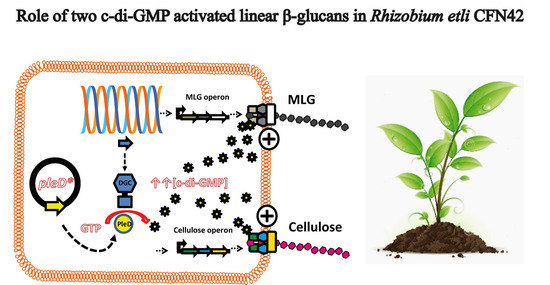
1. Introduction
2. Materials and Methods
2.1. Bacteria and Culture Conditions
2.2. Recombinant DNA Techniques
2.3. Construction of bgsA (RHE_PE00363) Mutant and bgsA celAB Double Mutant
2.4. Preparation of mRNA and Quantitative RT-PCR Assay
2.5. Calcofluor Binding Assays
2.6. Biofilm Assays
2.7. β-Glucan Isolation and Characterization
2.8. Plant Root Binding Assays
2.9. Symbiotic Assays
2.10. Nodules Histology and Microscopy
2.11. Phylogenetic Analyses
2.12. Statistical Analyses
3. Results
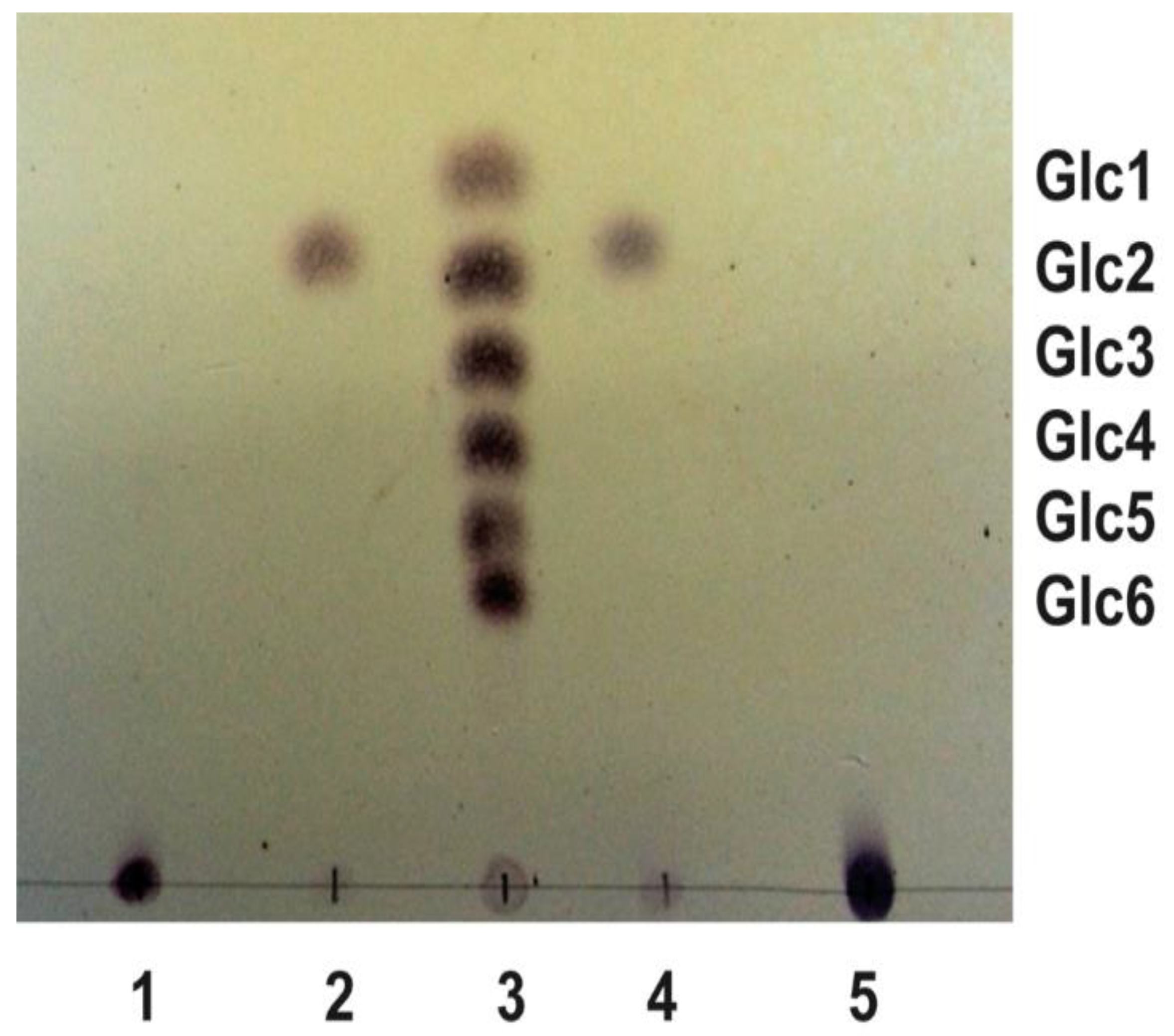
3.1. Rhizobium etli CFN42 Produces Two Linear β-Glucans Activated by c-di-GMP
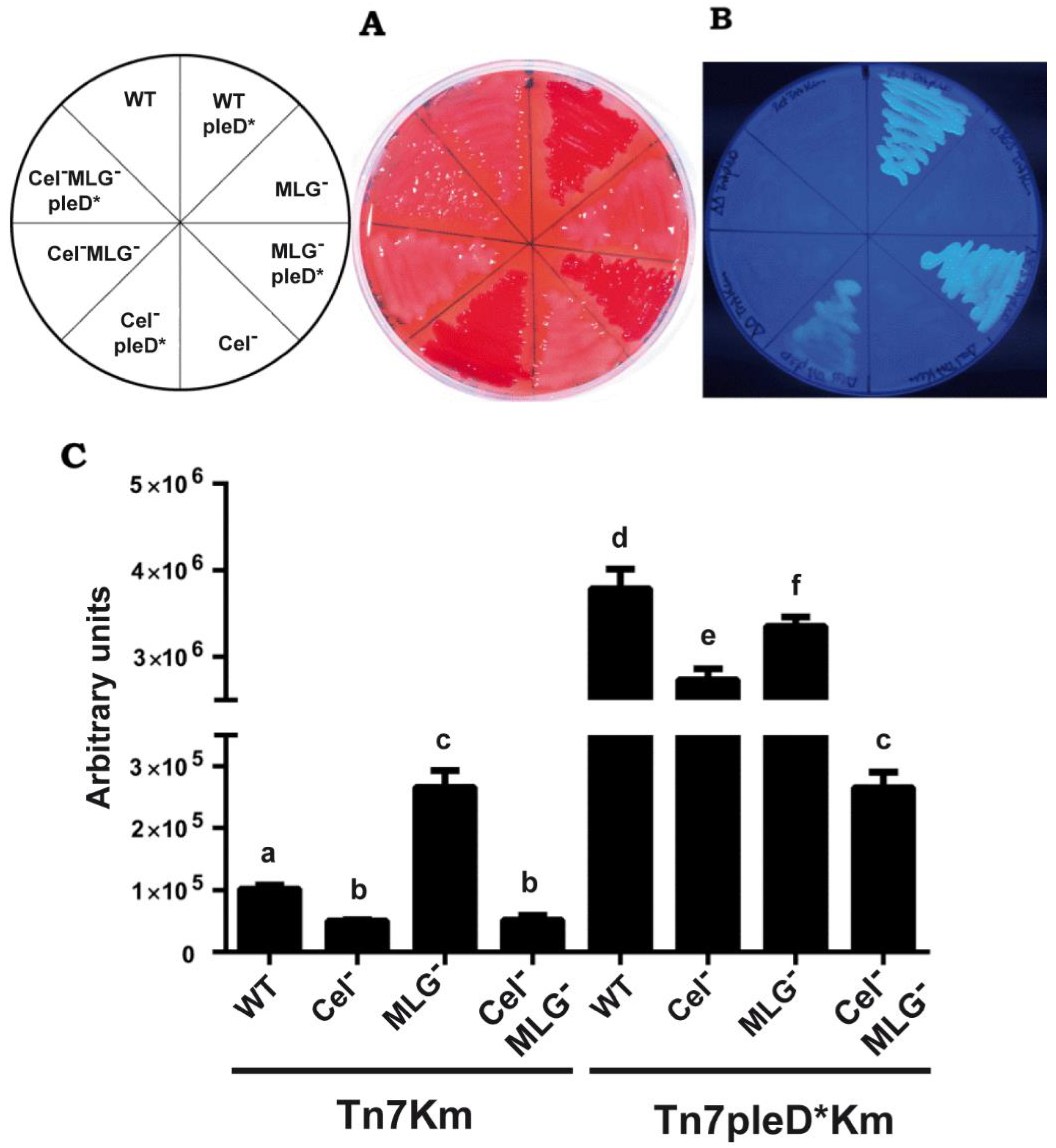
3.2. Cyclic-di-GMP Activation of MLG Production by Rhizobium etli CFN42
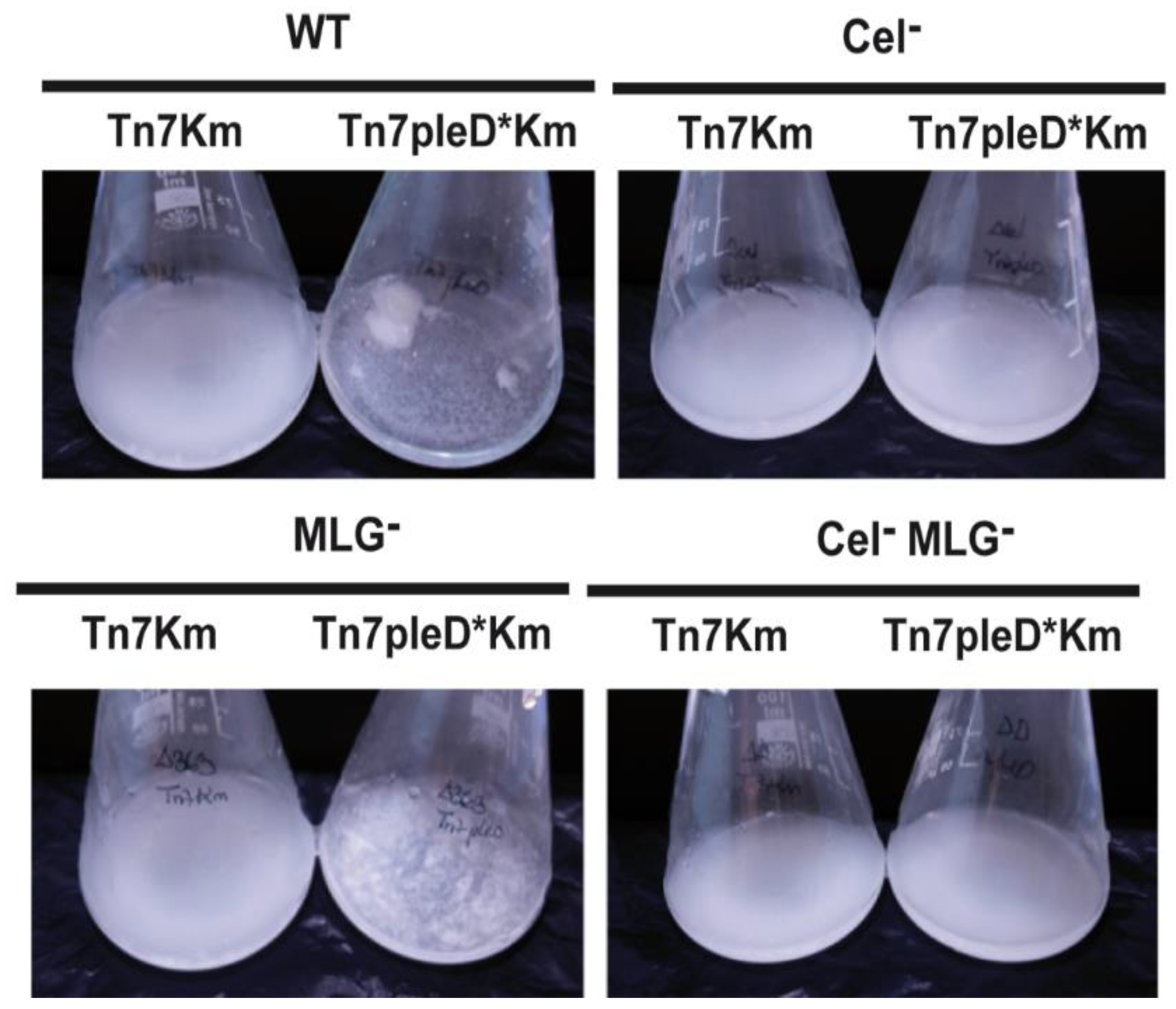
3.3. Cyclic-di-GMP Activation of Cellulose and MLG Production in Rhizobium etli CFN42

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Relevant Characteristic | Reference or Source |
|---|---|---|
| Rhizobia strains | ||
| R. etli CFN42 | Ret; Wild-type | [34] |
| S. meliloti pJBpleD* | Sme 8530 strain with pJBpleD* plasmid | [7] |
| Ret Tn7Km | Ret with a mini-Tn7Km transposon | [23] |
| Ret Tn7pleD*Km | Ret with a mini-Tn7pleD*Km transposon | [23] |
| Ret Tn7pleD*Tc | Ret with a mini-Tn7pleD*Tc transposon | [23] |
| LR102 | Ret Cel− Tn7Km; CFN42 ΔcelAB with a mini-Tn7Km transposon | This work |
| LR101 | Ret Cel− Tn7pleD*Km; CFN42 ΔcelAB with a mini-Tn7pleD*Km transposon | This work |
| LR104 | Ret MLG− Tn7Km; CFN42 ΔbgsA (RHE_PE00363) with a mini-Tn7Km transposon | This work |
| LR103 | Ret MLG− Tn7 pleD*Km; CFN42 ΔbgsA (RHE_PE00363) with a mini-Tn7pleD*Km transposon | This work |
| LR106 | Ret Cel− MLG− Tn7Km; CFN42 ΔcelAB and ΔbgsA with a mini-Tn7Km transposon | This work |
| LR105 | Ret Cel− MLG− Tn7pleD*Km; CFN42 ΔcelAB and ΔbgsA with a mini-Tn7pleD*Km transposon | This work |
| Escherichia coli strains | ||
| DH5α | supE44, ∆lacU169, Φ80, lacZ∆M1, recA1, endA1, gyrA96, thi1, relA1, 5hsdR171 | [35] |
| S17.1 | Tmpr, Smr, Spr; thi, pro, recA, hsdR, hsdM, Rp4Tc::Mu, Km::Tn7 | [36] |
| β2163 | (F−) RP4-2-Tc::Mu _dapA::(erm-pir) [Kmr Emr] | [22] |
| OmniMAX | F′ [proAB+ lacIq lacZΔM15 Tn10(TetR) Δ(ccdAB)] mcrA Δ(mrr-hsdRMS-mcrBC) φ80(lacZ)ΔM15 Δ(lacZYA-argF) U169 endA1 recA1 supE44 thi-1 gyrA96 relA1 tonA panD | Invitrogen |
| Plasmids | ||
| pK18mobsacB | Kmr; mobilizable suicide plasmid | [24] |
| pK18ΔbgsA | Kmr; pK18mobsacB carrying the deleted version of the bgsA(RHE_PE00363) gene | This work |
| pCR-2.1-TOPO | Kmr Apr; cloning vector for PCR products | Invitrogen |
| TOPOΔbgsA | Kmr Apr; pCR-2.1-TOPO carrying the deleted version of bgsA(RHE_PE00363) gene | This work |
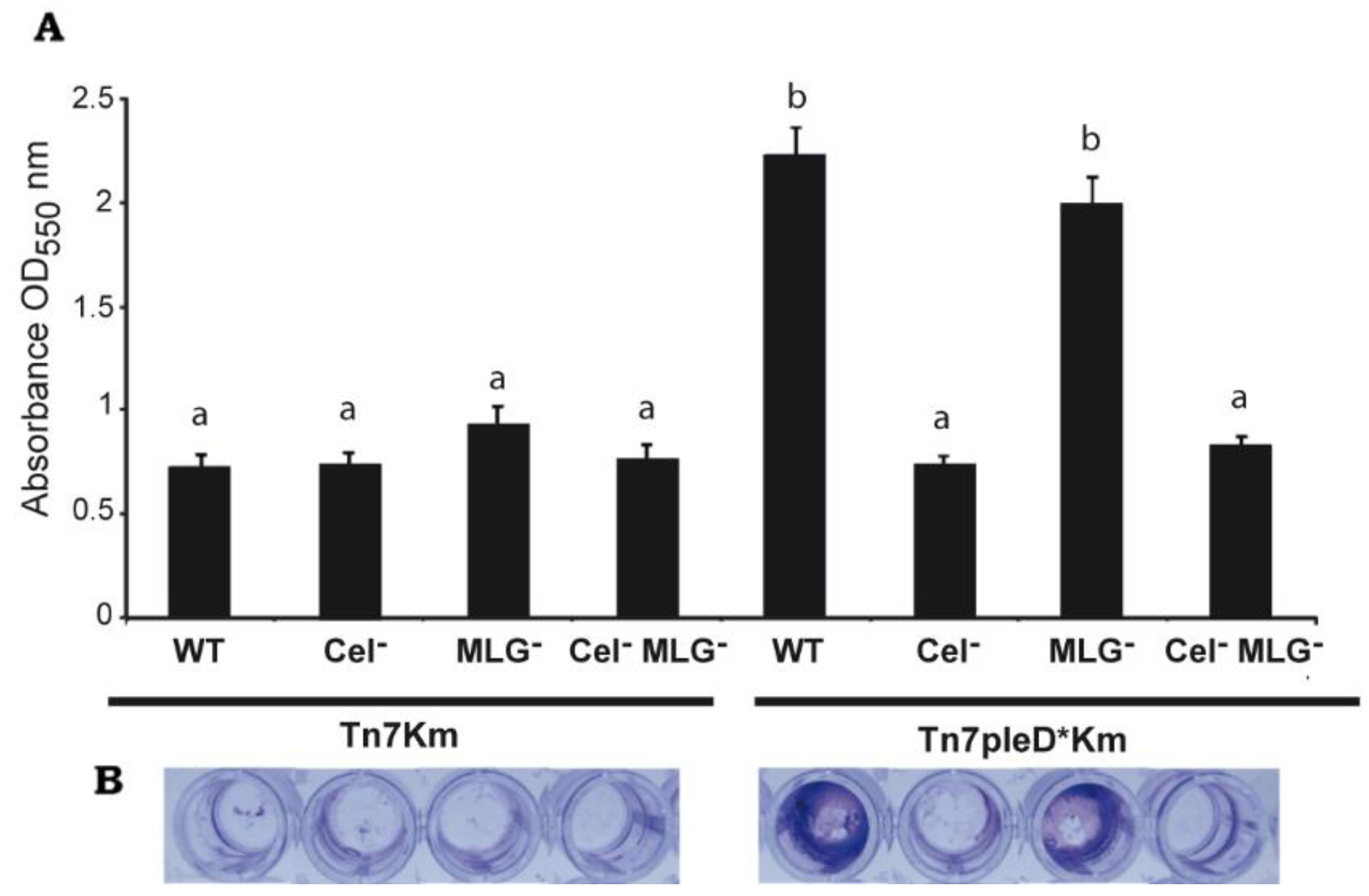
3.4. Role of Cellulose and MLG in Rhizobium etli CFN42 Aggregation and Biofilm Formation
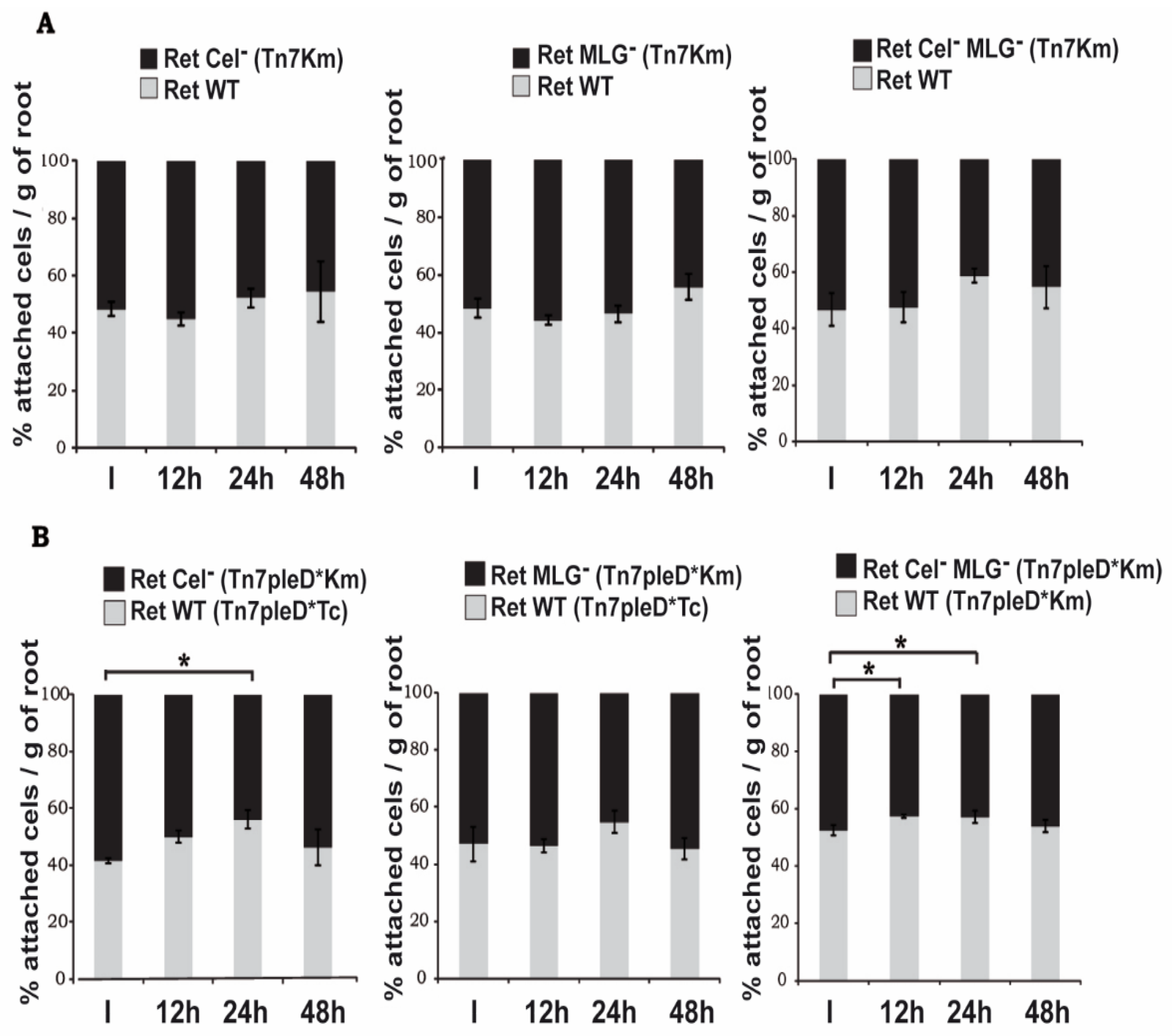
3.5. Role of Cellulose and MLG in Rhizobium etli CFN42 Adhesion to Plant Roots
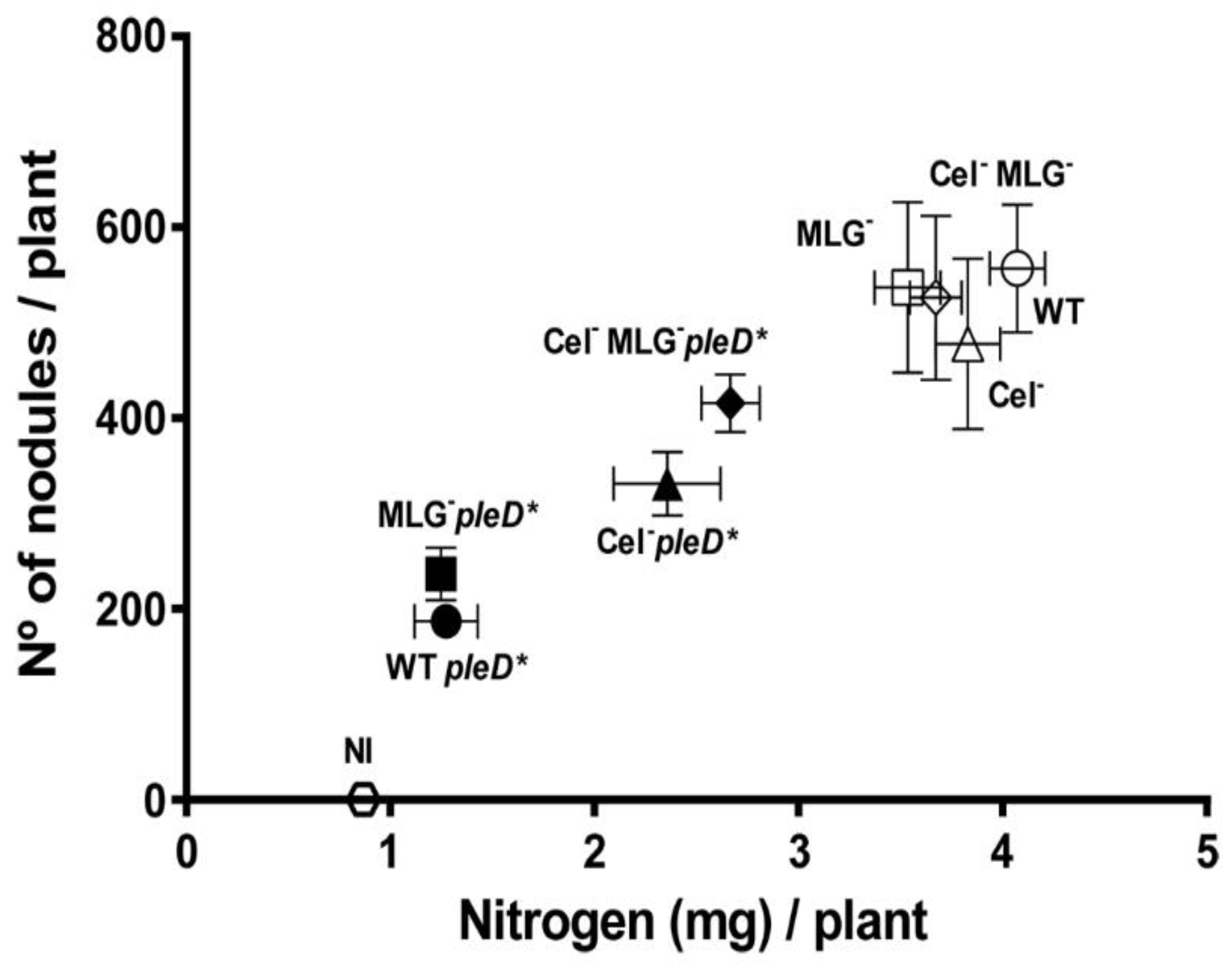
3.6. Role of Cellulose and MLG in Rhizobium etli CFN42 Symbiotic Interaction with the Legume Plant
3.7. Impact of High c-di-GMP on Nodule Structure
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bontemps-Gallo, S.; Bohin, J.P.; Lacroix, J.M. Osmoregulated Periplasmic Glucans. EcoSal Plus 2017, 7, 1–21. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, M.; Stone, B.A.; Stanisich, V.A. Curdlan and other bacterial (1-->3)-beta-D-glucans. Appl. Microbiol. Biotechnol. 2005, 68, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Stone, B.A. Chapter 2.1—Chemistry of β-Glucans. In Chemistry, Biochemistry, and Biology of 1-3 Beta Glucans and Related Polysaccharides; Bacic, A., Fincher, G.B., Stone, B.A., Eds.; Academic Press: San Diego, CA, USA, 2009; pp. 5–46. [Google Scholar]
- Harada, T.; Misaki, A.; Saito, H. Curdlan: A bacterial gel-forming β-1, 3-glucan. Arch. Biochem. Biophys. 1968, 124, 292–298. [Google Scholar] [CrossRef]
- Salmeán, A.A.; Duffieux, D.; Harholt, J.; Qin, F.; Michel, G.; Czjzek, M.; Willats, W.G.T.; Hervé, C. Insoluble (1 → 3), (1 → 4)-β-D-glucan is a component of cell walls in brown algae (Phaeophyceae) and is masked by alginates in tissues. Sci. Rep. 2017, 7, 2880. [Google Scholar] [CrossRef]
- Simmons, T.J.; Uhrin, D.; Gregson, T.; Murray, L.; Sadler, I.H.; Fry, S.C. An unexpectedly lichenase-stable hexasaccharide from cereal, horsetail and lichen mixed-linkage beta-glucans (MLGs): Implications for MLG subunit distribution. Phytochemistry 2013, 95, 322–332. [Google Scholar] [CrossRef]
- Pérez-Mendoza, D.; Rodríguez-Carvajal, M.A.; Romero-Jiménez, L.; Farias, G.A.; Lloret, J.; Gallegos, M.T.; Sanjuan, J. Novel mixed-linkage beta-glucan activated by c-di-GMP in Sinorhizobium meliloti. Proc. Natl. Acad. Sci. USA 2015, 112, E757. [Google Scholar] [CrossRef]
- Ross, P.; Weinhouse, H.; Aloni, Y.; Michaeli, D.; Weinbergerohana, P.; Mayer, R.; Braun, S.; Devroom, E.; Vandermarel, G.A.; Vanboom, J.H.; et al. Regulation of Cellulose Synthesis in Acetobacter xylinum by Cyclic Diguanylic Acid. Nature 1987, 325, 279–281. [Google Scholar] [CrossRef]
- Hengge, R.; Häussler, S.; Pruteanu, M.; Stülke, J.; Tschowri, N.; Turgay, K. Recent Advances and Current Trends in Nucleotide Second Messenger Signaling in Bacteria. J. Mol. Biol. 2019, 431, 908–927. [Google Scholar] [CrossRef]
- Petchiappan, A.; Naik, S.Y.; Chatterji, D. Tracking the homeostasis of second messenger cyclic-di-GMP in bacteria. Biophys. Rev. 2020, 12, 719–730. [Google Scholar] [CrossRef]
- Hengge, R. Novel tricks played by the second messenger c-di-GMP in bacterial biofilm formation. EMBO J. 2013, 32, 322–323. [Google Scholar] [CrossRef]
- Pérez-Mendoza, D.; Sanjuán, J. Exploiting the commons: Cyclic diguanylate regulation of bacterial exopolysaccharide production. Curr. Opin. Microbiol. 2016, 30, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Krol, E.; Schäper, S.; Becker, A. Cyclic di-GMP signaling controlling the free-living lifestyle of alpha-proteobacterial rhizobia. Biol. Chem. 2020, 401, 1335–1348. [Google Scholar] [CrossRef] [PubMed]
- Olivares, J.; Bedmar, E.J.; Sanjuan, J. Biological nitrogen fixation in the context of global change. Mol. Plant Microbe Interact. 2013, 26, 486–494. [Google Scholar] [CrossRef]
- Schäper, S.; Krol, E.; Skotnicka, D.; Kaever, V.; Hilker, R.; Sogaard-Andersen, L.; Becker, A. Cyclic Di-GMP Regulates Multiple Cellular Functions in the Symbiotic Alphaproteobacterium Sinorhizobium meliloti. J. Bacteriol. 2016, 198, 521–535. [Google Scholar] [CrossRef]
- Schäper, S.; Steinchen, W.; Krol, E.; Altegoer, F.; Skotnicka, D.; Søgaard-Andersen, L.; Bange, G.; Becker, A. AraC-like transcriptional activator CuxR binds c-di-GMP by a PilZ-like mechanism to regulate extracellular polysaccharide production. Proc. Natl. Acad. Sci. USA 2017, 114, E4822–E4831. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Mendoza, D.; Bertinetti, D.; Lorenz, R.; Gallegos, M.T.; Herberg, F.W.; Sanjuan, J. A novel c-di-GMP binding domain in glycosyltransferase BgsA is responsible for the synthesis of a mixed-linkage beta-glucan. Sci. Rep. 2017, 7, 8997. [Google Scholar] [CrossRef]
- Gao, S.; Romdhane, S.B.; Beullens, S.; Kaever, V.; Lambrichts, I.; Fauvart, M.; Michiels, J. Genomic analysis of cyclic-di-GMP-related genes in rhizobial type strains and functional analysis in Rhizobium etli. Appl. Microbiol. Biotechnol. 2014, 98, 4589–4602. [Google Scholar] [CrossRef]
- Pérez-Mendoza, D.; Aragón, I.M.; Prada-Ramírez, H.A.; Romero-Jiménez, L.; Ramos, C.; Gallegos, M.T.; Sanjuán, J. Responses to Elevated c-di-GMP Levels in Mutualistic and Pathogenic Plant-Interacting Bacteria. PLoS ONE 2014, 9, e91645. [Google Scholar] [CrossRef]
- Beringer, J.E. R Factor Transfer in Rhizobium-Leguminosarum. J. Gen. Microbiol. 1974, 84, 188–198. [Google Scholar] [CrossRef]
- Robertsen, B.K.; Aman, P.; Darvill, A.G.; Mcneil, M.; Albersheim, P. The Structure of Acidic Extracellular Polysaccharides Secreted by Rhizobium-Leguminosarum and Rhizobium-Trifolii. Plant Physiol. 1981, 67, 389–400. [Google Scholar] [CrossRef]
- Demarre, G.; Guerout, A.M.; Matsumoto-Mashimo, C.; Rowe-Magnus, D.A.; Marliere, P.; Mazel, D. A new family of mobilizable suicide plasmids based on broad host range R388 plasmid (IncW) and RP4 plasmid (IncPalpha) conjugative machineries and their cognate Escherichia coli host strains. Res. Microbiol. 2005, 156, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Romero-Jiménez, L.; Rodríguez-Carbonell, D.; Gallegos, M.T.; Sanjuán, J.; Pérez-Mendoza, D. Mini-Tn7 vectors for stable expression of diguanylate cyclase PleD* in Gram-negative bacteria. BMC Microbiol. 2015, 15, 190. [Google Scholar] [CrossRef] [PubMed]
- Schäfer, A.; Tauch, A.; Jäger, W.; Kalinowski, J.; Thierbach, G.; Pühler, A. Small mobilizable multi-purpose cloning vectors derived from the Escherichia coli plasmids pK18 and pK19: Selection of defined deletions in the chromosome of Corynebacterium glutamicum. Gene 1994, 145, 69–73. [Google Scholar] [CrossRef]
- Blom, W.; Luteyn, J.C.; Kelholt-Dijkman, H.H.; Huijmans, J.G.; Loonen, M.C. Thin-layer chromatography of oligosaccharides in urine as a rapid indication for the diagnosis of lysosomal acid maltase deficiency (Pompe’s disease). Clin. Chim. Acta Int. J. Clin. Chem. 1983, 134, 221–227. [Google Scholar] [CrossRef]
- Rigaud, J.; Puppo, A. Indole-3 Acetic Acid Catabolism by Soybean Bacteroids. J. Gen. Microbiol. 1975, 88, 223–228. [Google Scholar] [CrossRef]
- Redondo, F.J.; de la Peña, T.C.; Morcillo, C.N.; Lucas, M.M.; Pueyo, J.J. Overexpression of flavodoxin in bacteroids induces changes in antioxidant metabolism leading to delayed senescence and starch accumulation in alfalfa root nodules. Plant Physiol. 2009, 149, 1166–1178. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Schmid, J.; Heider, D.; Wendel, N.J.; Sperl, N.; Sieber, V. Bacterial Glycosyltransferases: Challenges and opportunities of a highly diverse enzyme class toward tailoring natural products. Front. Microbiol. 2016, 7, 182. [Google Scholar] [CrossRef]
- Drula, E.; Garron, M.-L.; Dogan, S.; Lombard, V.; Henrissat, B.; Terrapon, N. The carbohydrate-active enzyme database: Functions and literature. Nucleic Acids Res. 2021, 50, D571–D577. [Google Scholar] [CrossRef] [PubMed]
- Marketon, M.M.; González, J.E. Identification of two quorum-sensing systems in Sinorhizobium meliloti. J. Bacteriol. 2002, 184, 3466–3475. [Google Scholar] [CrossRef] [PubMed]
- Quinto, C.; Delavega, H.; Flores, M.; Leemans, J.; Cevallos, M.A.; Pardo, M.A.; Azpiroz, R.; Girard, M.D.; Calva, E.; Palacios, R. Nitrogenase Reductase—A Functional Multigene Family in Rhizobium phaseoli. Proc. Natl. Acad. Sci. USA 1985, 82, 1170–1174. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D. Studies on transformation of Escherichia coli with Plasmids. J. Mol. Biol. 1983, 166, 557–580. [Google Scholar] [CrossRef]
- Simon, R.; Priefer, U.; Pühler, A. A Broad Host Range Mobilization System for in vivo Genetic-Engineering-Transposon Mutagenesis in Gram-Negative Bacteria. Biotechnology 1983, 1, 784–791. [Google Scholar] [CrossRef]
- Xu, J.; Kim, J.; Koestler, B.J.; Choi, J.H.; Waters, C.M.; Fuqua, C. Genetic analysis of Agrobacterium tumefaciens unipolar polysaccharide production reveals complex integrated control of the motile-to-sessile switch. Mol. Microbiol. 2013, 89, 929–948. [Google Scholar] [CrossRef]
- Pesavento, C.; Becker, G.; Sommerfeldt, N.; Possling, A.; Tschowri, N.; Mehlis, A.; Hengge, R. Inverse regulatory coordination of motility and curli-mediated adhesion in Escherichia coli. Genes Dev. 2008, 22, 2434–2446. [Google Scholar] [CrossRef] [PubMed]
- Lindenberg, S.; Klauck, G.; Pesavento, C.; Klauck, E.; Hengge, R. The EAL domain protein YciR acts as a trigger enzyme in a c-di-GMP signalling cascade in E. coli biofilm control. EMBO J. 2013, 32, 2001–2014. [Google Scholar] [CrossRef]
- Römling, U.; Bian, Z.; Hammar, M.; Sierralta, W.D.; Normark, S. Curli fibers are highly conserved between Salmonella typhimurium and Escherichia coli with respect to operon structure and regulation. J. Bacteriol. 1998, 180, 722–731. [Google Scholar] [CrossRef]
- Römling, U.; Sierralta, W.D.; Eriksson, K.; Normark, S. Multicellular and aggregative behaviour of Salmonella typhimurium strains is controlled by mutations in the agfD promoter. Mol. Microbiol. 1998, 28, 249–264. [Google Scholar] [CrossRef]
- Zogaj, X.; Nimtz, M.; Rohde, M.; Bokranz, W.; Römling, U. The multicellular morphotypes of Salmonella typhimurium and Escherichia coli produce cellulose as the second component of the extracellular matrix. Mol. Microbiol. 2001, 39, 1452–1463. [Google Scholar] [CrossRef] [PubMed]
- Morgan, J.L.; McNamara, J.T.; Zimmer, J. Mechanism of activation of bacterial cellulose synthase by cyclic di-GMP. Nat. Struct. Mol. Biol. 2014, 21, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Ahmad, I.; Blanka, A.; Schottkowski, M.; Cimdins, A.; Galperin, M.Y.; Römling, U.; Gomelsky, M. GIL, a new c-di-GMP-binding protein domain involved in regulation of cellulose synthesis in enterobacteria. Mol. Microbiol. 2014, 93, 439–452. [Google Scholar] [CrossRef] [PubMed]
- Serra, D.O.; Richter, A.M.; Klauck, G.; Mika, F.; Hengge, R. Microanatomy at cellular resolution and spatial order of physiological differentiation in a bacterial biofilm. mBio 2013, 4, e00103. [Google Scholar] [CrossRef]
- Richter, A.M.; Possling, A.; Malysheva, N.; Yousef, K.P.; Herbst, S.; von Kleist, M.; Hengge, R. Local c-di-GMP Signaling in the Control of Synthesis of the E. coli Biofilm Exopolysaccharide pEtN-Cellulose. J. Mol. Biol. 2020, 432, 4576–4595. [Google Scholar] [CrossRef]
- Ruffing, A.M.; Chen, R.R. Transcriptome profiling of a curdlan-producing Agrobacterium reveals conserved regulatory mechanisms of exopolysaccharide biosynthesis. Microb. Cell Fact. 2012, 11, 17. [Google Scholar] [CrossRef]
- Orr, M.W.; Galperin, M.Y.; Lee, V.T. Sustained sensing as an emerging principle in second messenger signaling systems. Curr. Opin. Microbiol. 2016, 34, 119–126. [Google Scholar] [CrossRef]
- Ausmees, N.; Jonsson, H.; Hoglund, S.; Ljunggren, H.; Lindberg, M. Structural and putative regulatory genes involved in cellulose synthesis in Rhizobium leguminosarum bv. trifolii. Microbiology 1999, 145 Pt 5, 1253–1262. [Google Scholar] [CrossRef]
- Smit, G.; Kijne, J.W.; Lugtenberg, B.J. Involvement of both cellulose fibrils and a Ca2+-dependent adhesin in the attachment of Rhizobium leguminosarum to pea root hair tips. J. Bacteriol. 1987, 169, 4294–4301. [Google Scholar] [CrossRef]
- Xu, J.; Kim, J.; Danhorn, T.; Merritt, P.M.; Fuqua, C. Phosphorus limitation increases attachment in Agrobacterium tumefaciens and reveals a conditional functional redundancy in adhesin biosynthesis. Res. Microbiol. 2012, 163, 674–684. [Google Scholar] [CrossRef]
- Isenberg, R.Y.; Christensen, D.G.; Visick, K.L.; Mandel, M.J. High Levels of Cyclic Diguanylate Interfere with Beneficial Bacterial Colonization. mBio 2022, 13, e0167122. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Mendoza, D.; Romero-Jiménez, L.; Rodríguez-Carvajal, M.Á.; Lorite, M.J.; Muñoz, S.; Olmedilla, A.; Sanjuán, J. The Role of Two Linear β-Glucans Activated by c-di-GMP in Rhizobium etli CFN42. Biology 2022, 11, 1364. https://doi.org/10.3390/biology11091364
Pérez-Mendoza D, Romero-Jiménez L, Rodríguez-Carvajal MÁ, Lorite MJ, Muñoz S, Olmedilla A, Sanjuán J. The Role of Two Linear β-Glucans Activated by c-di-GMP in Rhizobium etli CFN42. Biology. 2022; 11(9):1364. https://doi.org/10.3390/biology11091364
Chicago/Turabian StylePérez-Mendoza, Daniel, Lorena Romero-Jiménez, Miguel Ángel Rodríguez-Carvajal, María J. Lorite, Socorro Muñoz, Adela Olmedilla, and Juan Sanjuán. 2022. "The Role of Two Linear β-Glucans Activated by c-di-GMP in Rhizobium etli CFN42" Biology 11, no. 9: 1364. https://doi.org/10.3390/biology11091364
APA StylePérez-Mendoza, D., Romero-Jiménez, L., Rodríguez-Carvajal, M. Á., Lorite, M. J., Muñoz, S., Olmedilla, A., & Sanjuán, J. (2022). The Role of Two Linear β-Glucans Activated by c-di-GMP in Rhizobium etli CFN42. Biology, 11(9), 1364. https://doi.org/10.3390/biology11091364