
Goitered Gazelle Gazella subgutturosa Responded to Human Disturbance by Increasing Vigilance Rather than Changing the Group Size

 , , and
, , and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
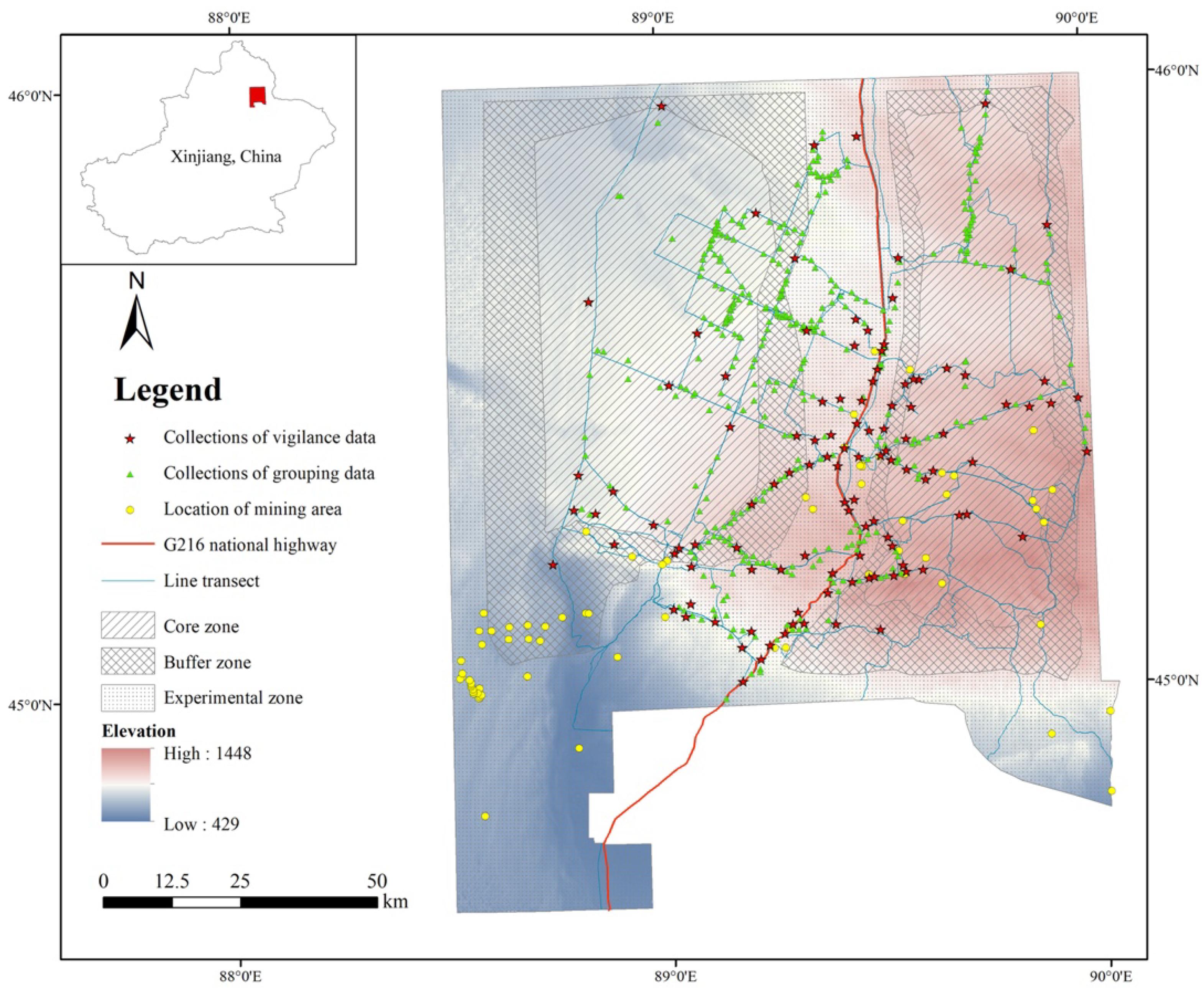
2.1. Study Area
2.2. Data Collection
2.3. Data Analysis
3. Results
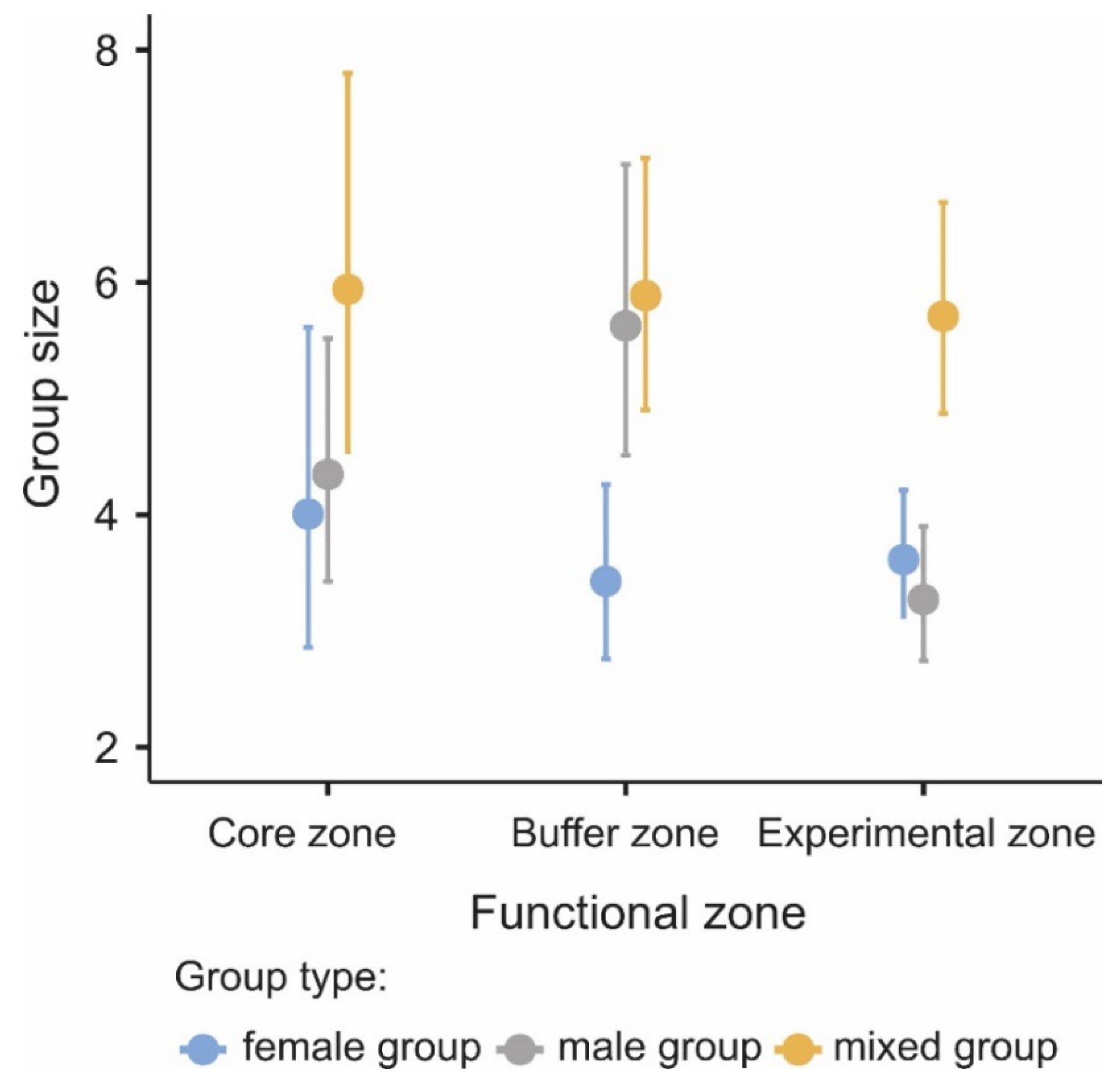
3.1. Difference in Group Size among Functional Zones
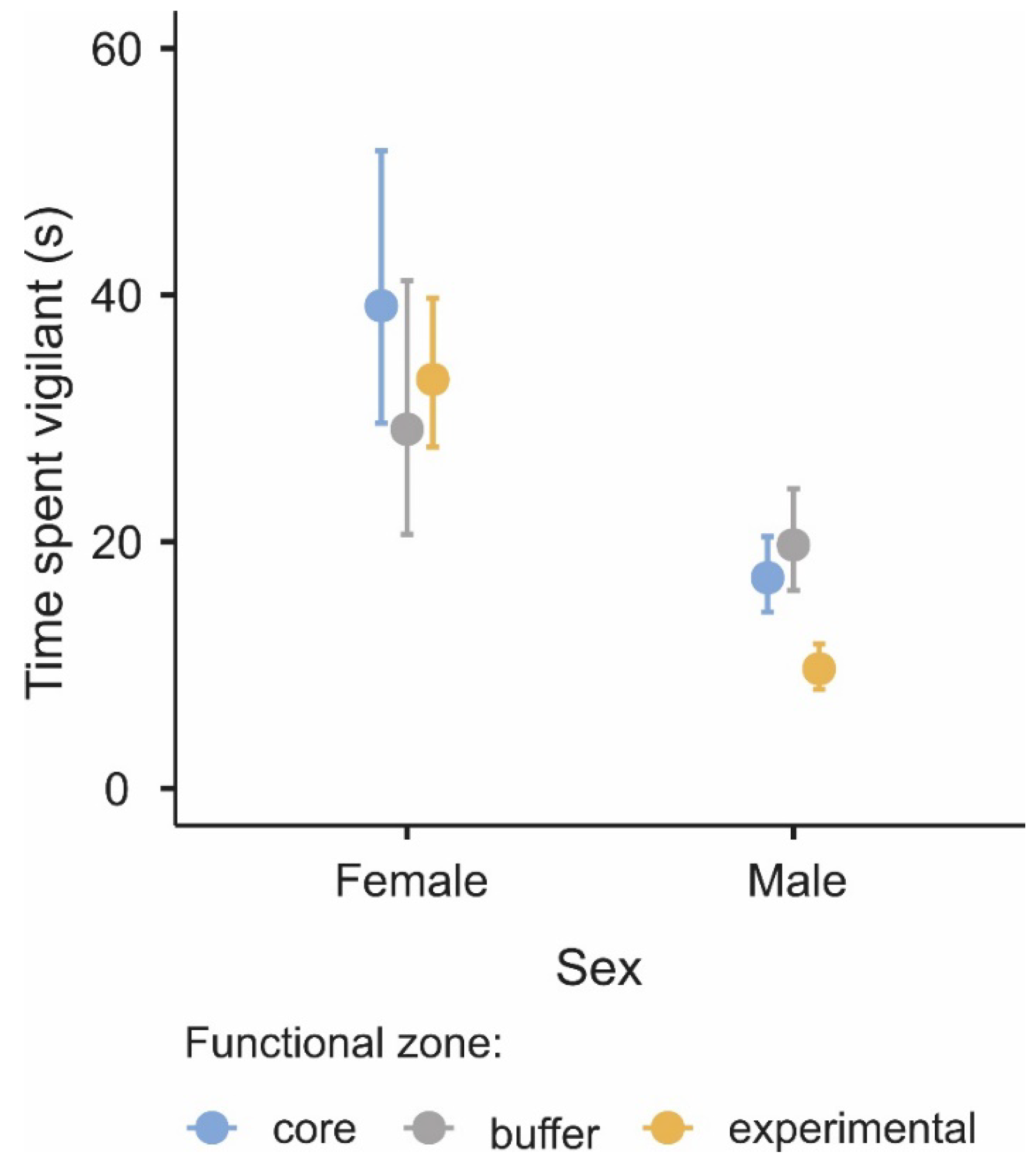
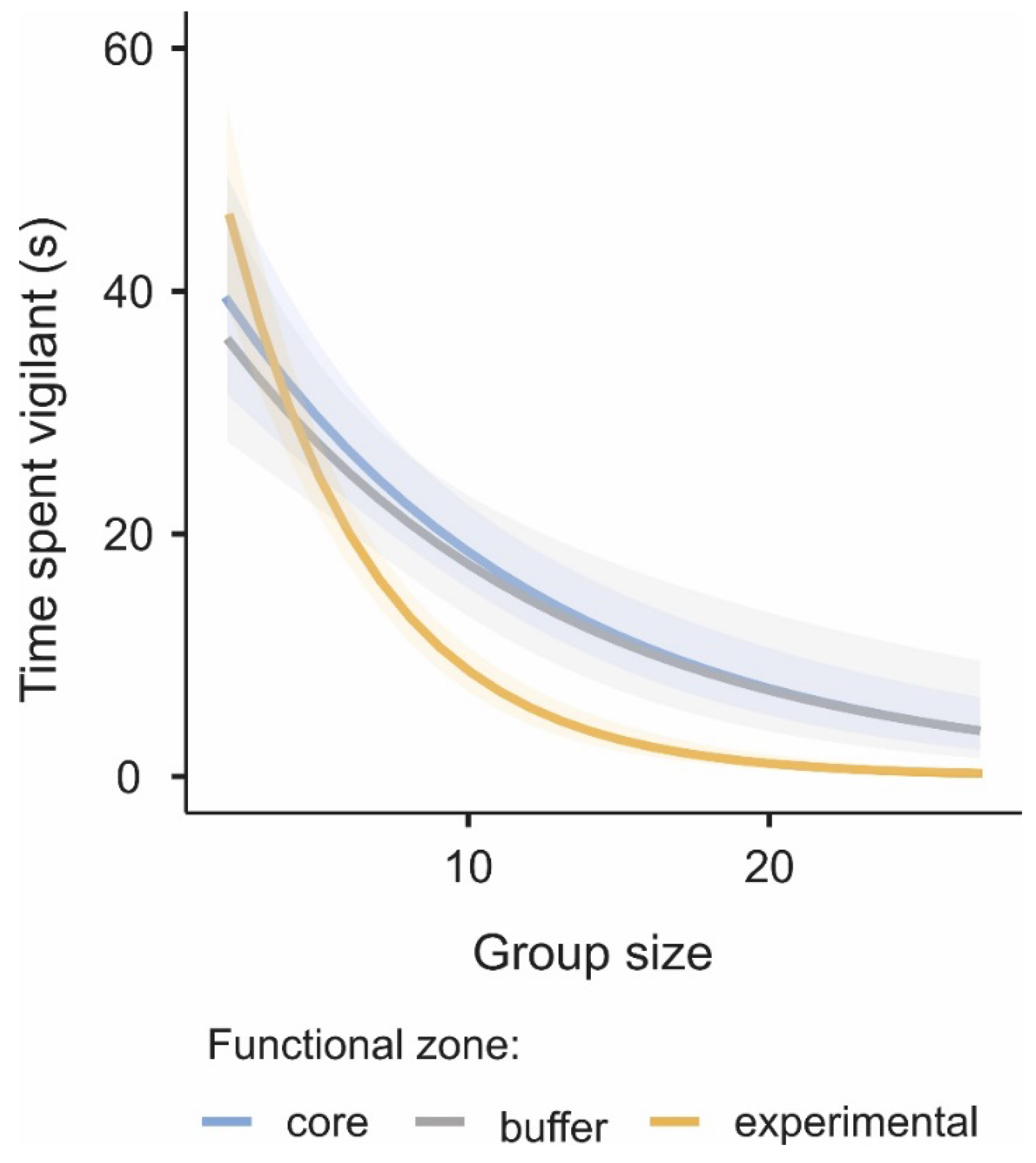
3.2. Difference in the Level of Vigilance among Functional Zones
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Frid, A.; Dill, L. Human-caused disturbance stimuli as a form of predation risk. Conserv. Ecol. 2002, 6, 11. [Google Scholar] [CrossRef]
- Jayakody, S.; Sibbald, A.M.; Gordon, I.J.; Lambin, X. Red deer Cervus elephus vigilance behaviour differs with habitat and type of human disturbance. Wildl. Biol. 2008, 14, 81–91. [Google Scholar] [CrossRef]
- Jiang, T.; Wang, X.; Ding, Y.; Liu, Z.; Wang, Z. Behavioral responses of blue sheep (Pseudois nayaur) to nonlethal human recreational disturbance. Chin. Sci. Bull. 2013, 58, 2237–2247. [Google Scholar] [CrossRef]
- Fischer, F.; Linsenmair, K. Changing social organization in an ungulate population subject to poaching and predation-the kob antelope (Kobus kob kob) in the Comoé National Park, Côte d’Ivoire. Afr. J. Ecol. 2007, 45, 285–292. [Google Scholar] [CrossRef]
- Stankowich, T. Ungulate flight responses to human disturbance: A review and meta-analysis. Biol. Conserv. 2008, 141, 2159–2173. [Google Scholar] [CrossRef]
- McLean, I.G. Conservation and the ontogeny of behaviour. In Behavioural Approaches to Conservation in the Wild; Clemmons, J.R., Buchholz, R., Eds.; Cambridge University Press: Cambridge, UK, 1997; p. 132156. [Google Scholar]
- Reed, J.M. Animal behavior as a tool in conservation biology. In Conservation Medicine: Ecological Health in Practise; Aquirre, A.A., Ostfeld, R.S., Tabor, G.M., House, C., Pearl, M.C., Eds.; Oxford University Press: Oxford, UK, 2002; pp. 145–163. [Google Scholar]
- Benhaiem, S.; Delon, M.; Lourtet, B.; Cargnelutti, B.; Aulagnier, S.; Hewison, A.J.M.; Morellet, N.; Verheyden, H. Hunting increases vigilance levels in roe deer and modifies feeding site selection. Anim. Behav. 2008, 76, 611–618. [Google Scholar] [CrossRef]
- Fernández-Juricic, E.; Schroeder, N. Do variations in scanning behavior affect tolerance to human disturbance? Appl. Anim. Behav. Sci. 2003, 84, 219–234. [Google Scholar] [CrossRef]
- Papouchis, C.M.; Singer, F.J.; Sloan, W.B. Responses of Desert Bighorn Sheep to Increased Human Recreation. J. Wildl. Manag. 2001, 65, 573–582. [Google Scholar] [CrossRef]
- Leweri, C.M.; Bartzke, G.S.; Msuha, M.J.; Treydte, A.C. Spatial and seasonal group size variation of wild mammalian herbivores in multiple use landscapes of the Ngorongoro Conservation Area, Tanzania. PLoS ONE 2022, 17, e0267082. [Google Scholar] [CrossRef]
- Wang, M.Y.; Ruckstuhl, K.E.; Xu, W.X.; Blank, D.; Yang, W.K. Human Activity Dampens the Benefits of Group Size on Vigilance in Khulan (Equus hemionus) in Western China. PLoS ONE 2016, 11, e0146725. [Google Scholar] [CrossRef]
- Li, C.; Jiang, Z.; Tang, S.; Zeng, Y. Evidence of effects of human disturbance on alert response in Pere David’s deer (Elaphurus davidianus). Zoo Biol. 2007, 26, 461–470. [Google Scholar] [CrossRef]
- Zheng, W.; Beauchamp, G.; Jiang, X.L.; Li, Z.Q.; Yang, Q.L. Determinants of vigilance in a reintroduced population of Pere David’s deer. Curr. Zool. 2013, 59, 265–270. [Google Scholar] [CrossRef]
- Brown, N.A.; Ruckstuhl, K.E.; Donelon, S.; Corbett, C. Changes in vigilance, grazing behaviour and spatial distribution of bighorn sheep due to cattle presence in Sheep River Provincial Park, Alberta. Agric. Ecosyst. Environ. 2010, 135, 226–231. [Google Scholar] [CrossRef]
- Caro, T.M. Demography and behaviour of African mammals subject to exploitation. Biol. Conserv. 1999, 91, 91–97. [Google Scholar] [CrossRef]
- Kie, J.G.; Bowyer, R.T. Sexual Segregation in White-Tailed Deer: Density-Dependent Changes in Use of Space, Habitat Selection, and Dietary Niche. J. Mammal. 1999, 80, 1004–1020. [Google Scholar] [CrossRef]
- Périquet, S.; Todd-Jones, L.; Valeix, M.; Stapelkamp, B.; Elliot, N.; Wijers, M.; Pays, O.; Fortin, D.; Madzikanda, H.; Fritz, H.; et al. Influence of immediate predation risk by lions on the vigilance of prey of different body size. Behav. Ecol. 2012, 23, 970–976. [Google Scholar] [CrossRef]
- Pulliam, H.R. On the advantages of flocking. J. Theor. Biol. 1973, 38, 419–422. [Google Scholar] [CrossRef]
- Vine, I. Risk of visual detection and pursuit by a predator and the selective advantage of flocking behaviour. J. Theor. Biol. 1971, 30, 405–422. [Google Scholar] [CrossRef]
- Lima, S.L.; Bednekoff, P.A. Temporal Variation in Danger Drives Antipredator Behavior: The Predation Risk Allocation Hypothesis. Am. Nat. 1999, 153, 649–659. [Google Scholar] [CrossRef]
- Jarman, P.J. Social-organization of antelope in telation to their ecology. Behaviour 1974, 48, 215–267. [Google Scholar] [CrossRef]
- Manor, R.; Saltz, D. Impact of Human Nuisance Disturbance on Vigilance and Group Size of a Social Ungulate. Ecol. Appl. 2003, 13, 1830–1834. [Google Scholar] [CrossRef]
- Blank, D.A.; Ruckstuhl, K.E.; Yang, W. Sexual segregation in goitered gazelles (Gazella subgutturosa). Can. J. Zool. 2012, 90, 955–960. [Google Scholar] [CrossRef]
- Wang, M.; Liu, W.; Alves da Silva, A.; Xu, W.; Yang, W.; Ruckstuhl, K.E.; Alves, J. Low size dimorphism does not lead to reduced sexual segregation: Exploring effects of habitat divergence and activity. Anim. Behav. 2021, 179, 225–233. [Google Scholar] [CrossRef]
- Qiao, J.F.; Yang, W.K.; Xu, W.X.; Xia, C.J.; Liu, W.; Blank, D. Social Structure of Goitred gazelles Gazella subgutturosa in Xinjiang, China. Pak. J. Zool. 2011, 43, 769–775. [Google Scholar]
- Wronski, T.; Cunningham, P.L. Seasonal changes in group size and composition of Arabian sand gazelle Gazella subgutturosa marica Thomas, 1897 during a period of drought in central western Saudi Arabia. Curr. Zool. 2011, 57, 36–42. [Google Scholar] [CrossRef]
- Blank, D.; Yang, W.; Xia, C.; Xu, W. Grouping pattern of the goitered gazelle, Gazella subgutturosa (Cetartiodactyla: Bovidae) in Kazakhstan. Mammalia 2012, 76, 149–155. [Google Scholar] [CrossRef]
- Zhang, X.C.; Shao, C.L.; Ge, Y.; Chen, C.; Xu, W.X.; Yang, W.K. Suitable summer habitat of the khulan in the Mt. Kalamaili Ungulate Nature Reserve and estimation of its population. Ying Yong Sheng Tai Xue Bao 2020, 31, 2993–3004. [Google Scholar] [CrossRef]
- Holmern, T.; Mkama, S.; Muya, J.; Røskaft, E. Intraspecific prey choice of bushmeat hunters outside the Serengeti National Park, Tanzania: A preliminary analysis. Afr. Zool. 2006, 41, 81–87. [Google Scholar] [CrossRef]
- Hassan, S.N. Effects of Fire on Large Herbivores and Their Forage Resources in Serengeti, Tanzania; Norwegian University of Science and Technology: Trondheim, Norway, 2007. [Google Scholar]
- Chu, H.; Jiang, Z.; Ge, Y.; Jiang, F.; Tao, Y.; Wang, C. Population densities and number of khulan and goitred gazelle in Mt. Kalamaili Ungulate Nature Reserve. Biodivers. Sci. 2009, 17, 414–422. [Google Scholar]
- Peng, X. The current status and protection of Asiatic wild ass (Equus hemionus) at Kalamailishan Nature Reserve. Chin. J. Wildl. 2015, 36, 162–165. [Google Scholar]
- Clutton-Brock, T.H.; Guinness, F.E.; Albon, S. Red Deer: Behavior and Ecology of Two Sexes; University of Chicago: Chicago, IL, USA, 1982. [Google Scholar]
- Zuur, A.; Leno, E.; Walker, N. Mixed Effects Models and Extensions in Ecology with R.; Springer: Berlin/Heidelberg, Germany, 2009. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. (Version 4.0); R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Creel, S.; Schuette, P.; Christianson, D. Effects of predation risk on group size, vigilance, and foraging behavior in an African ungulate community. Behav. Ecol. 2014, 25, 773–784. [Google Scholar] [CrossRef]
- Setsaas, T.; Hunninck, L.; Jackson, C.R.; May, R.; Røskaft, E. The impacts of human disturbances on the behaviour and population structure of impala (Aepyceros melampus) in the Serengeti ecosystem, Tanzania. Glob. Ecol. Conserv. 2018, 16, e00467. [Google Scholar] [CrossRef]
- Maurer, A.S.; Cove, M.V.; Siegal, O.M.; Lashley, M.A. Urbanization affects the behavior of a predator-free ungulate in protected lands. Landsc. Urban Plan. 2022, 222, 104391. [Google Scholar] [CrossRef]
- Blank, D.A. Vigilance, staring and escape running in antipredator behavior of goitered gazelle. Behav. Processes 2018, 157, 408–416. [Google Scholar] [CrossRef]
- Averbeck, C.; Apio, A.; Plath, M.; Wronski, T. Hunting differentially affects mixed-sex and bachelor-herds in a gregarious ungulate, the impala (Aepyceros melampus: Bovidae). Afr. J. Ecol. 2010, 48, 255–264. [Google Scholar] [CrossRef]
- Averbeck, C.; Plath, M.; Wronski, T.; Apio, A. Effect of human nuisance on the social organisation of large mammals: Group sizes and compositions of seven ungulate species in Lake Mburo National Park and the adjacent Ankole Ranching Scheme. Wildl. Biol. 2012, 18, 180–193. [Google Scholar] [CrossRef]
- Shi, J.; Dunbar, R.I.M.; Buckland, D.; Miller, D. Dynamics of grouping patterns and social segregation in feral goats (Capra hircus) on the Isle of Rum, NW Scotland. Mamm 2005, 69, 185–199. [Google Scholar] [CrossRef]
- Han, L.; Blanks, D.; Wang, M.Y.; Yang, W.K.; Alves Da Silva, A.; Alves, J. Grouping patterns and social organization in Siberian ibex (Capra sibirica): Feeding strategy matters. Folia Zool. 2019, 68, 35–42. [Google Scholar] [CrossRef]
- Lei, R.; Jiang, Z.; Liu, B. Group pattern and social segregation in Przewalski’s gazelle (Procapra przewalskii) around Qinghai Lake, China. J. Zool. 2001, 255, 175–180. [Google Scholar] [CrossRef]
- Wang, Z.; Li, Z.; Beauchamp, G.; Jiang, Z. Flock size and human disturbance affect vigilance of endangered red-crowned cranes (Grus japonensis). Biol. Conserv. 2011, 144, 101–105. [Google Scholar] [CrossRef]
- Wang, M.-Y.; Chen, Q.; Kuerbanjiang, H.; Xu, F.; Blank, D.; Yang, W.-K. Group size and disturbance effects on group vigilance in the Great Bustard Otis tarda in western China. Bird Study 2015, 62, 438–442. [Google Scholar] [CrossRef]
- Beauchamp, G. On how risk and group size interact to influence vigilance. Biol. Rev. Camb. Philos. Soc. 2019, 94, 1918–1934. [Google Scholar] [CrossRef]
- Saltz, D.; Berger-Tal, O.; Motro, U.; Shkedy, Y.; Raanan, N. Conservation implications of habituation in Nubian ibex in response to ecotourism. Anim. Conserv. 2019, 22, 220–227. [Google Scholar] [CrossRef]
- Xu, F.; Ma, M.; Yang, W.K.; Blank, D.; Ding, P.; Zhang, T. Group size effect on vigilance and daytime activity budgets of the Equus kiang (Equidae, Perissodactyla) in Arjinshan National Nature Reserve, Xinjiang, China. Folia Zool. 2013, 62, 76–81. [Google Scholar] [CrossRef]
- Fury, C.A.; Ruckstuhl, K.E.; Harrison, P.L. Spatial and social sexual segregation patterns in indo-pacific bottlenose dolphins (Tursiops aduncus). PLoS ONE 2013, 8, e52987. [Google Scholar] [CrossRef]
- Crosmary, W.-G.; Makumbe, P.; Côté, S.D.; Fritz, H. Vulnerability to predation and water constraints limit behavioural adjustments of ungulates in response to hunting risk. Anim. Behav. 2012, 83, 1367–1376. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | χ2 | df | p |
|---|---|---|---|
| Functional zone | 5.85 | 2.00 | 0.054 |
| Group type | 29.96 | 2.00 | <0.001 |
| Functional zone × Group type | 11.92 | 4.00 | <0.018 |
| Parameters | χ2 | df | p |
|---|---|---|---|
| Functional zone | 12.7 | 2.00 | 0.002 |
| Group size | 157.2 | 1.00 | <0.001 |
| Sex | 75.5 | 1.00 | <0.001 |
| Functional zone × sex | 13.8 | 2.00 | 0.001 |
| Group size × functional zone | 29.5 | 2.00 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, W.; Wang, M.; Blank, D.; Alves da Silva, A.; Yang, W.; Ruckstuhl, K.E.; Alves, J. Goitered Gazelle Gazella subgutturosa Responded to Human Disturbance by Increasing Vigilance Rather than Changing the Group Size. Biology 2022, 11, 1236. https://doi.org/10.3390/biology11081236
Xu W, Wang M, Blank D, Alves da Silva A, Yang W, Ruckstuhl KE, Alves J. Goitered Gazelle Gazella subgutturosa Responded to Human Disturbance by Increasing Vigilance Rather than Changing the Group Size. Biology. 2022; 11(8):1236. https://doi.org/10.3390/biology11081236
Chicago/Turabian StyleXu, Wenxuan, Muyang Wang, David Blank, António Alves da Silva, Weikang Yang, Kathreen E. Ruckstuhl, and Joana Alves. 2022. "Goitered Gazelle Gazella subgutturosa Responded to Human Disturbance by Increasing Vigilance Rather than Changing the Group Size" Biology 11, no. 8: 1236. https://doi.org/10.3390/biology11081236
APA StyleXu, W., Wang, M., Blank, D., Alves da Silva, A., Yang, W., Ruckstuhl, K. E., & Alves, J. (2022). Goitered Gazelle Gazella subgutturosa Responded to Human Disturbance by Increasing Vigilance Rather than Changing the Group Size. Biology, 11(8), 1236. https://doi.org/10.3390/biology11081236