
Frontal Sinus Morphological and Dimensional Variation as Seen on Computed Tomography Scans


Abstract
:Simple Summary
Abstract
1. Introduction
1.1. Morphology, Development, and Function of the Frontal Sinus
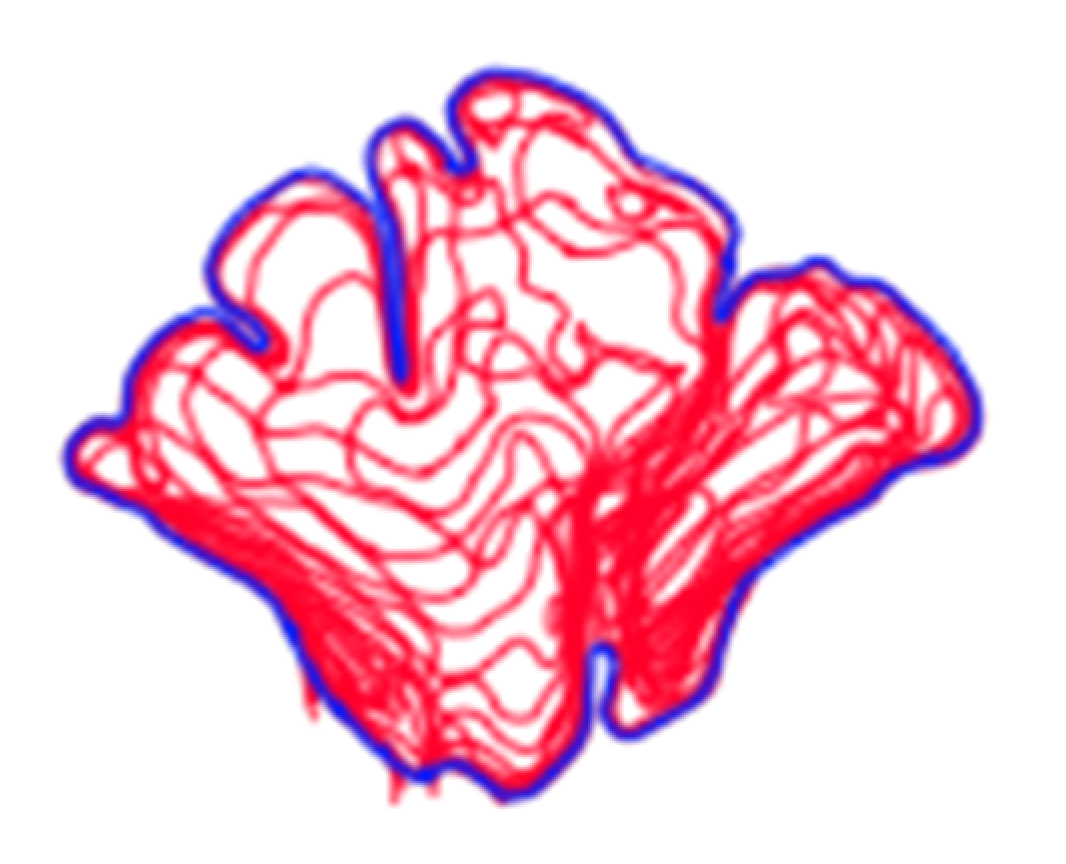
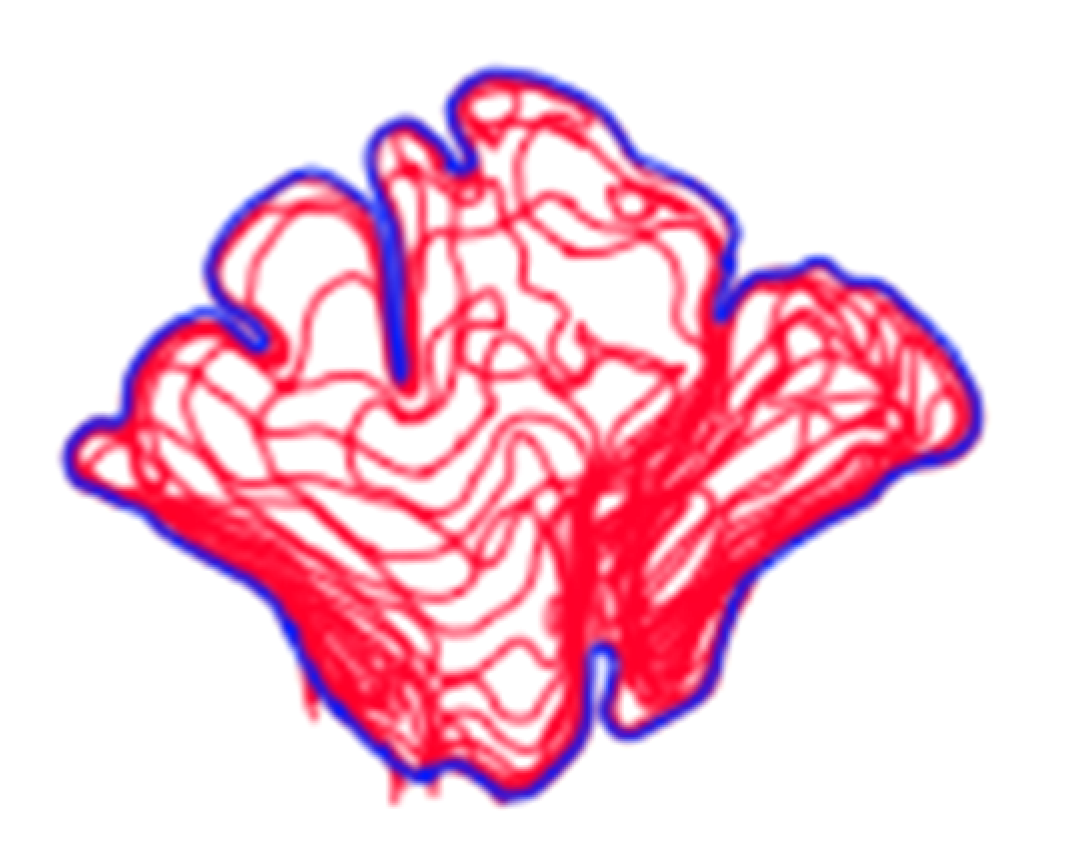
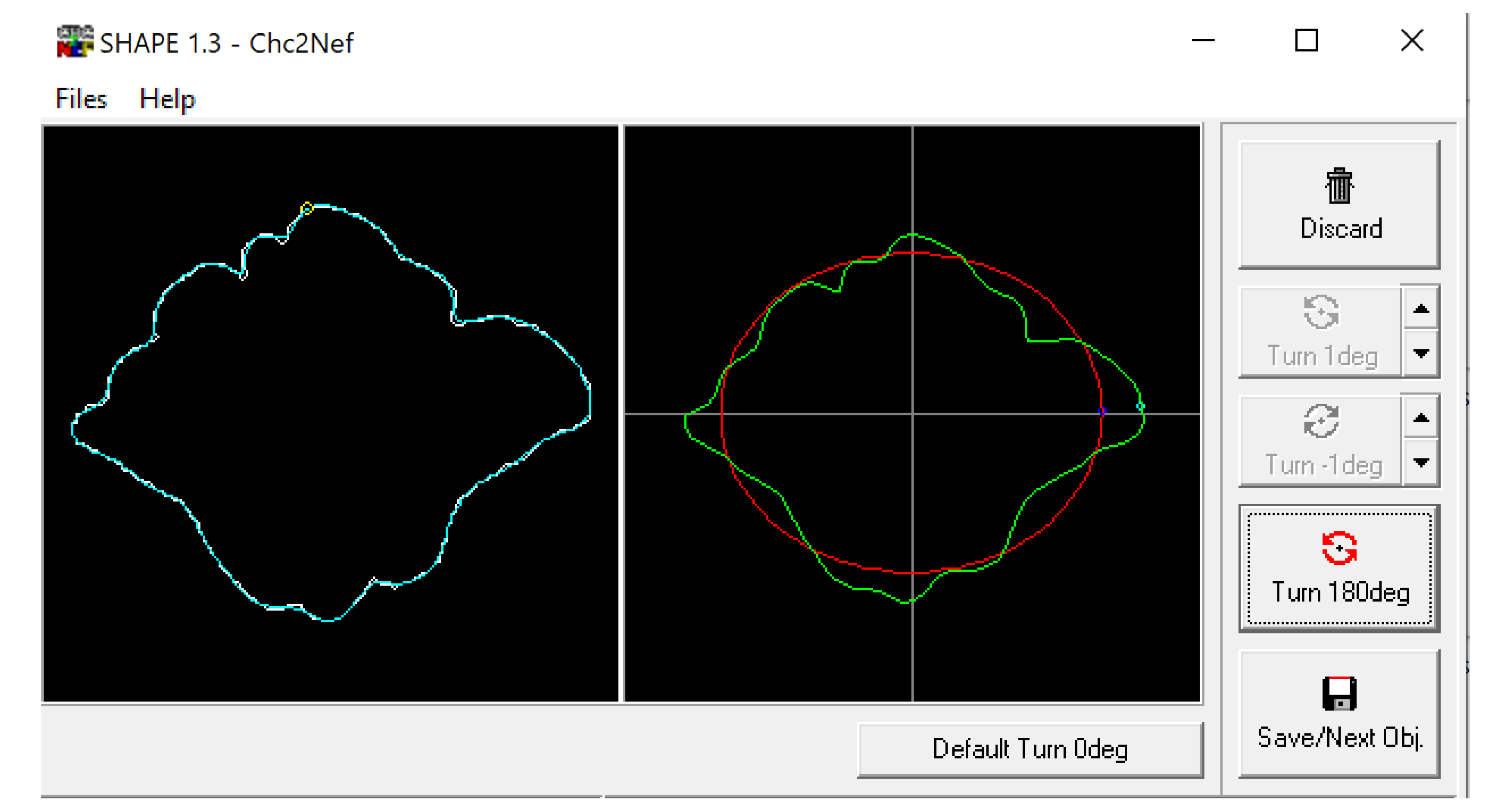
1.2. Visual Comparison and Superimposition
2. Materials and Methods
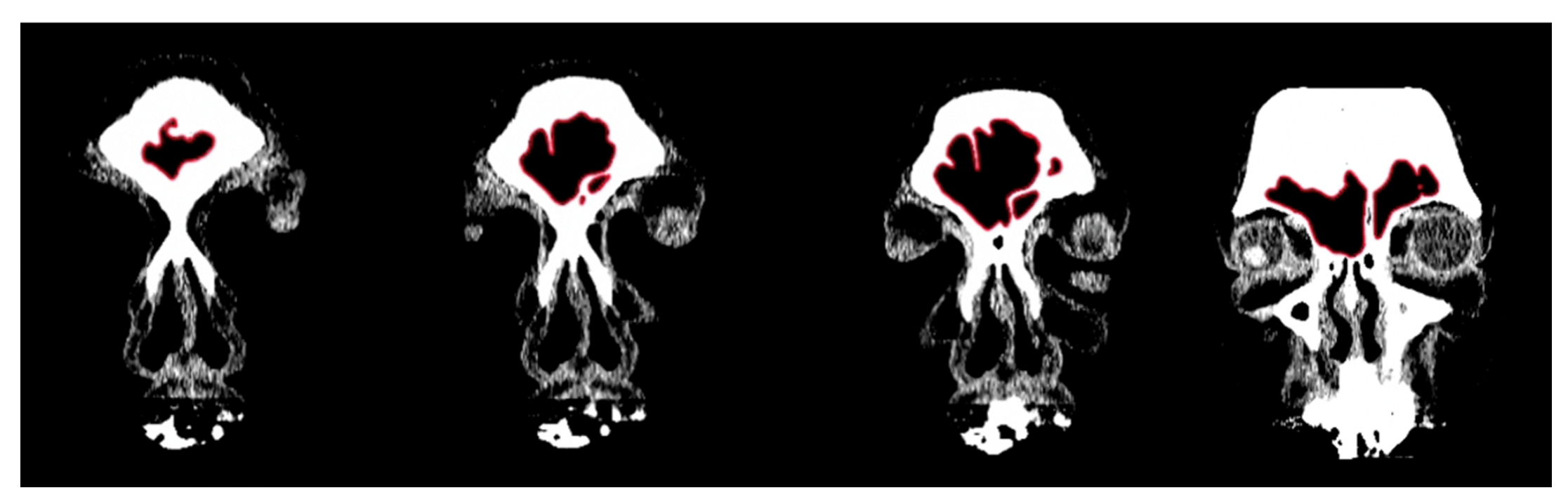
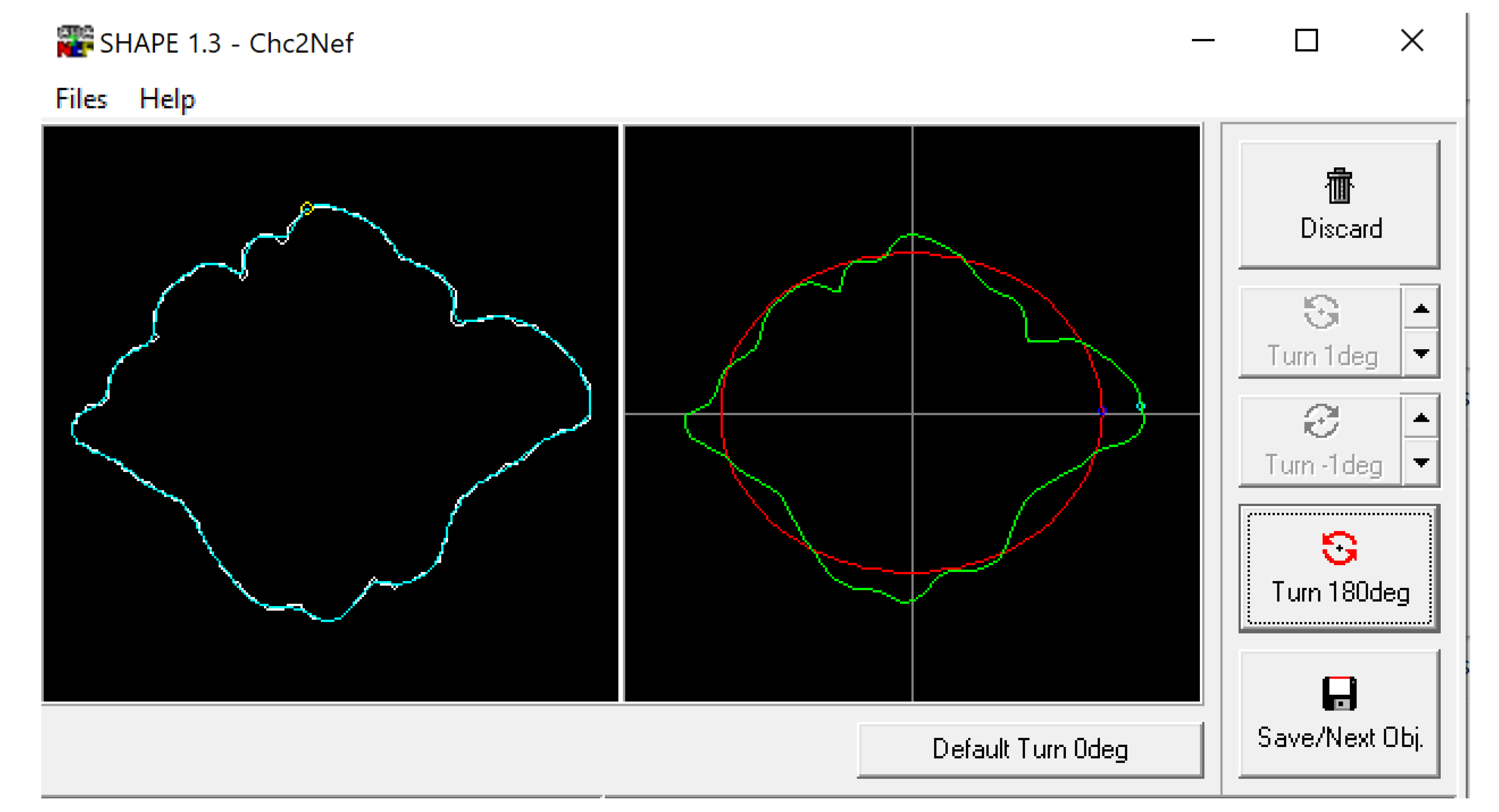
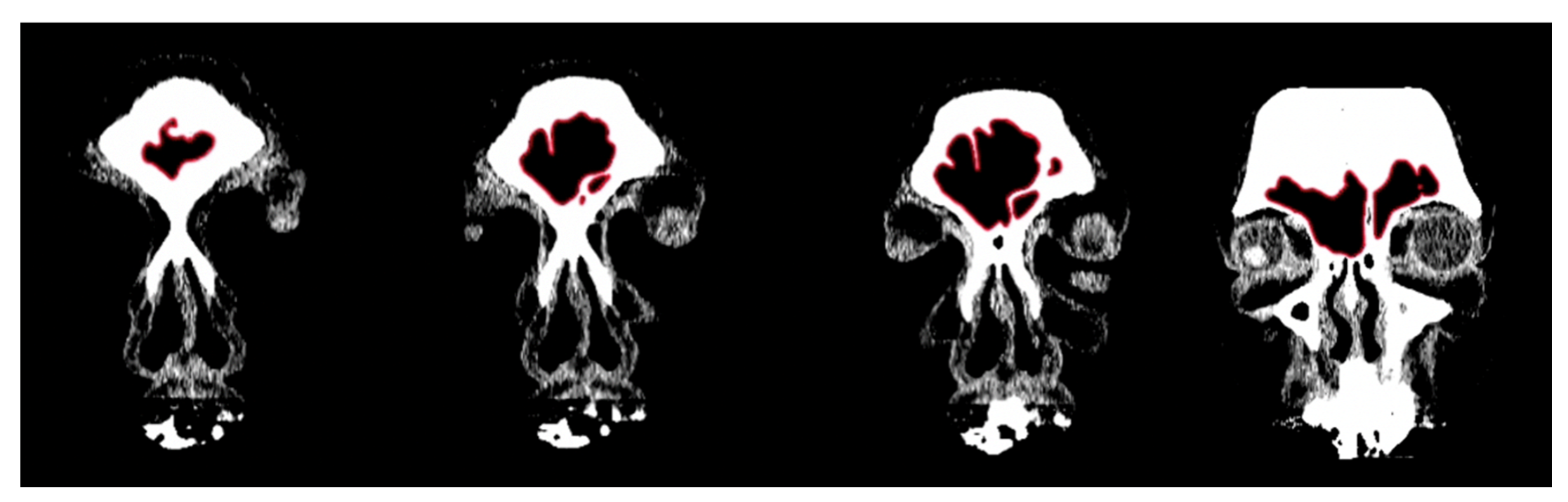
2.1. Morphological Analysis
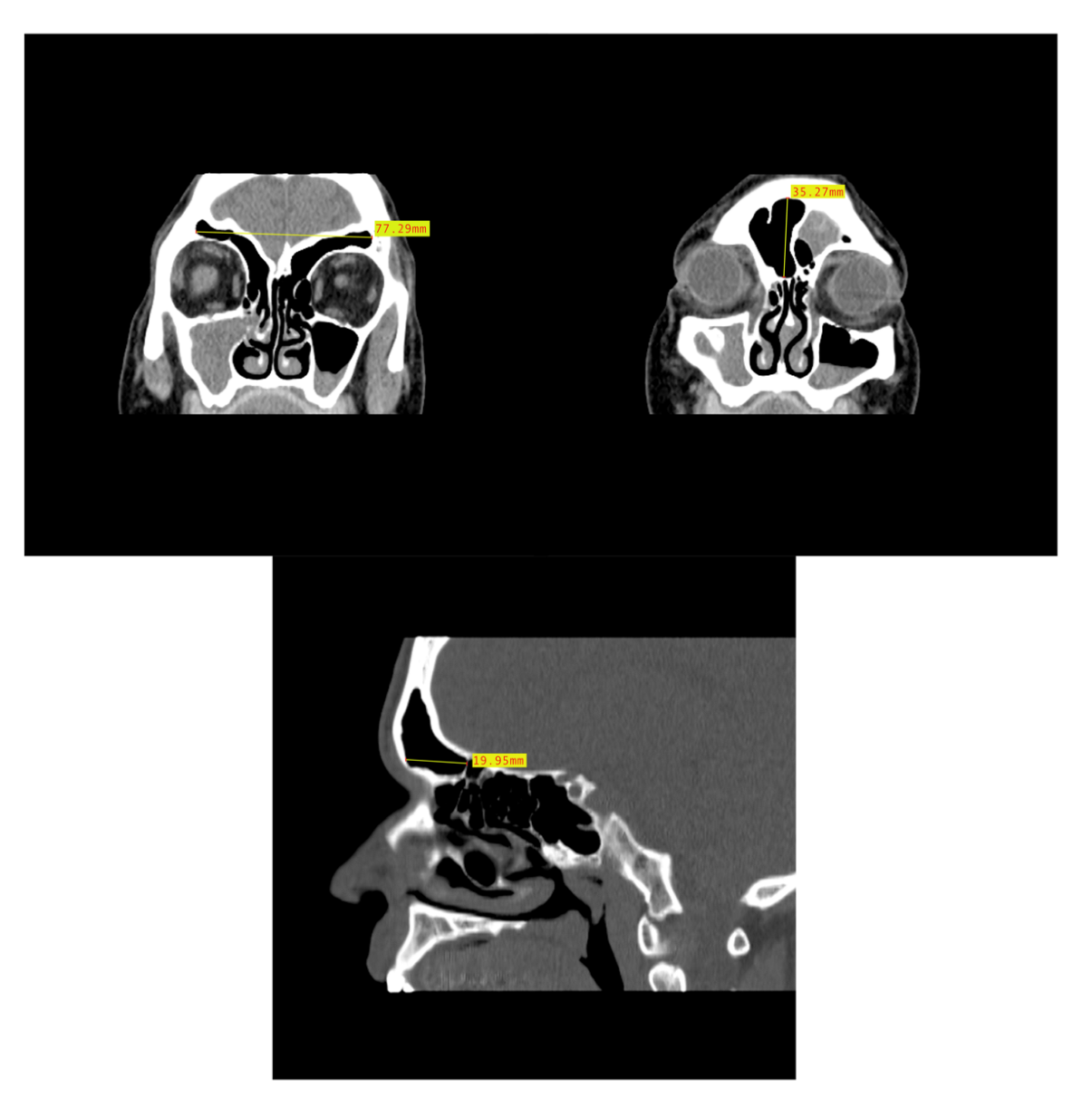
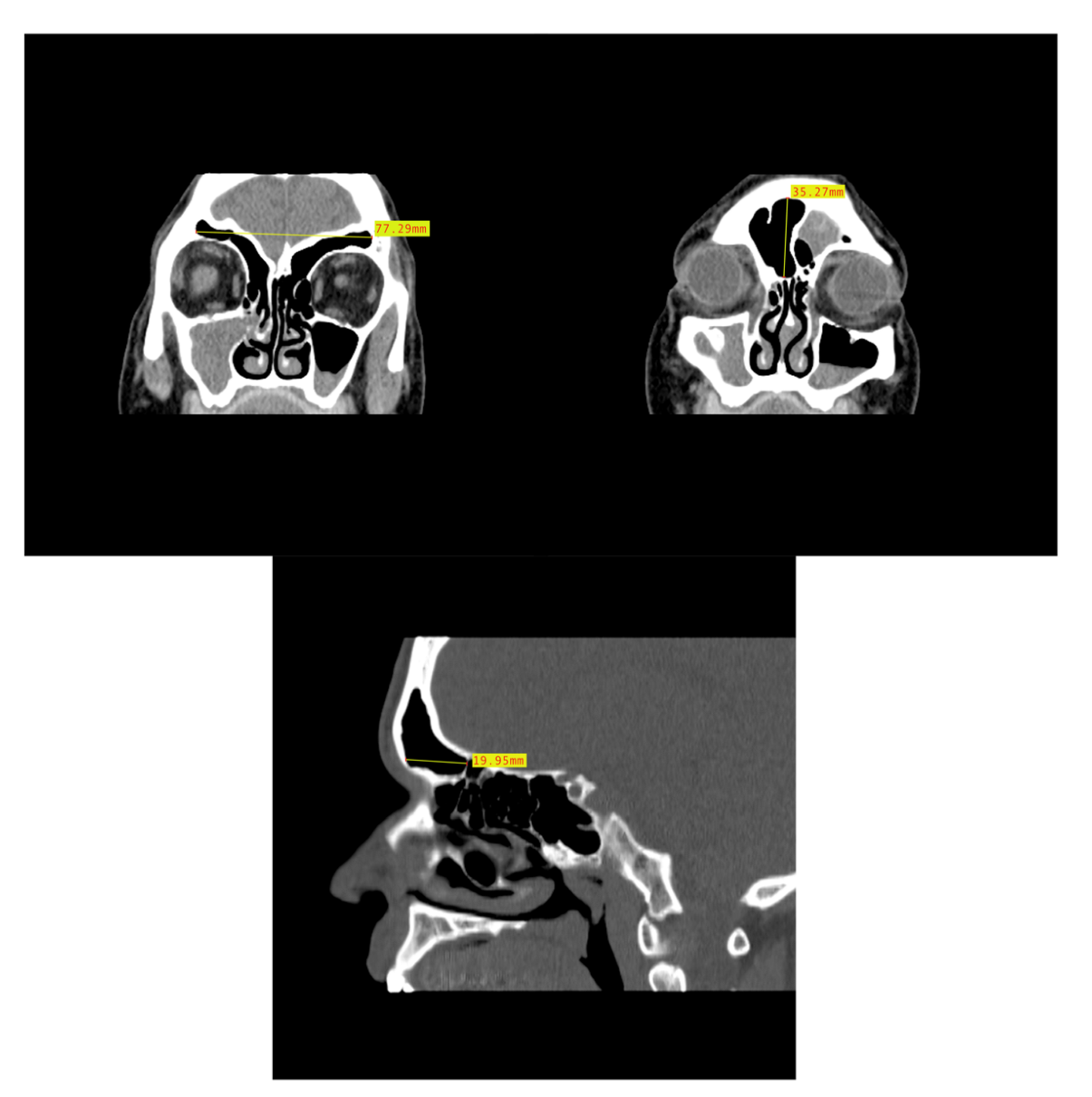
2.2. Dimensional Analysis
2.3. Statistical Analysis
3. Results
3.1. Shape Variation Analysis
3.2. Dimensional Variation Analysis
4. Discussion
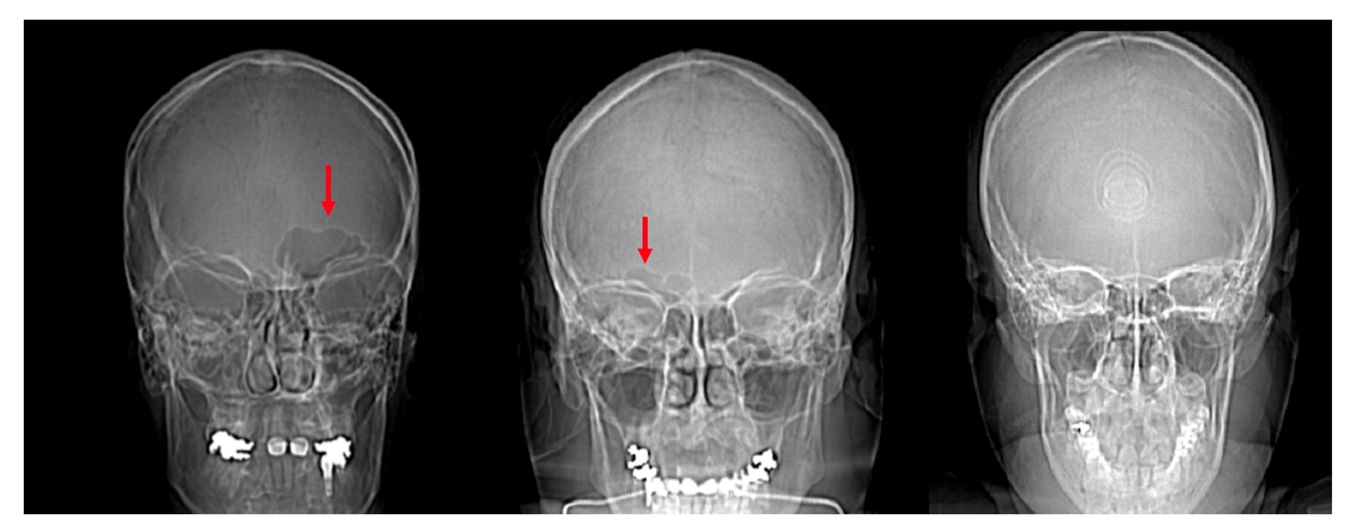
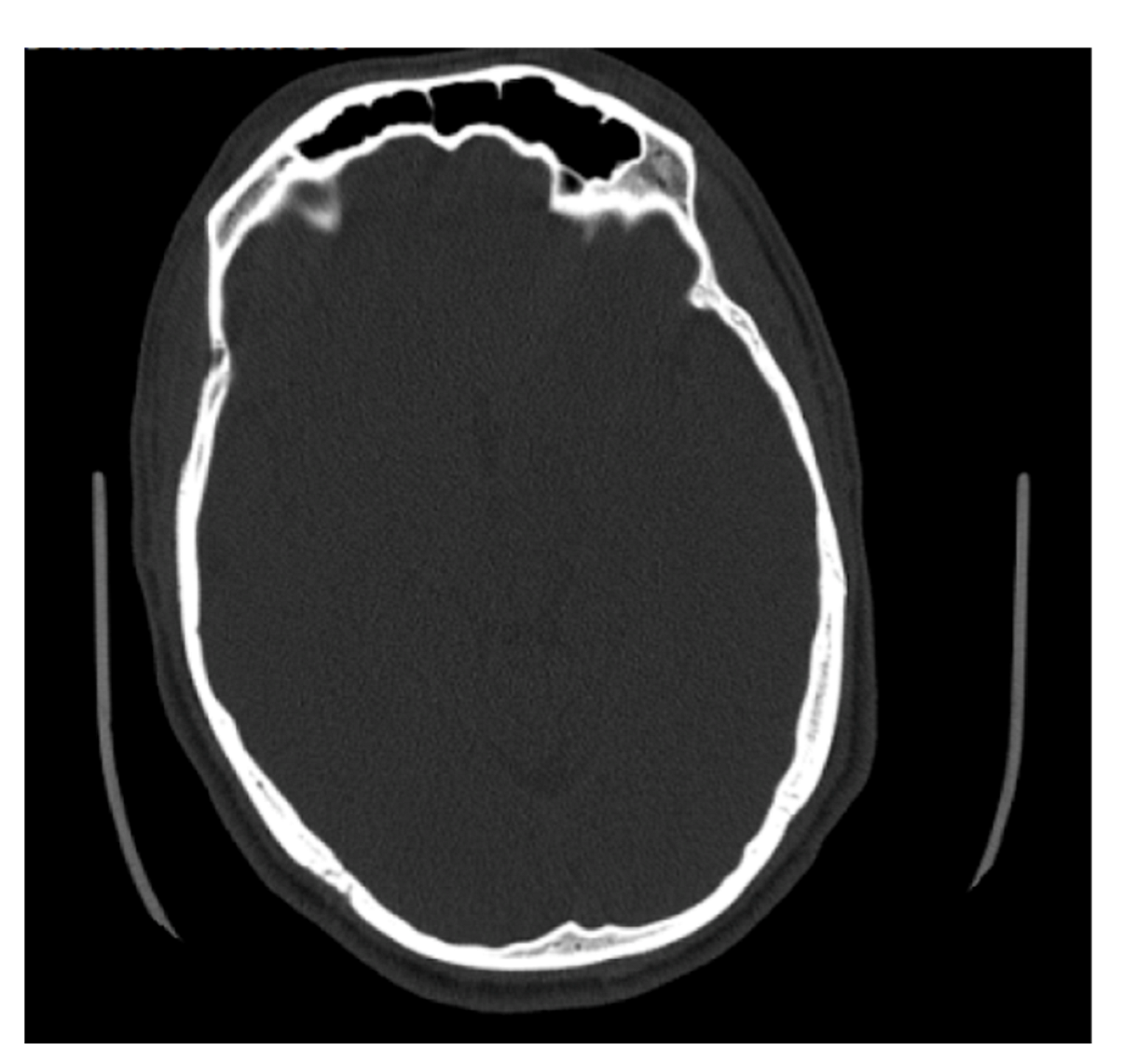
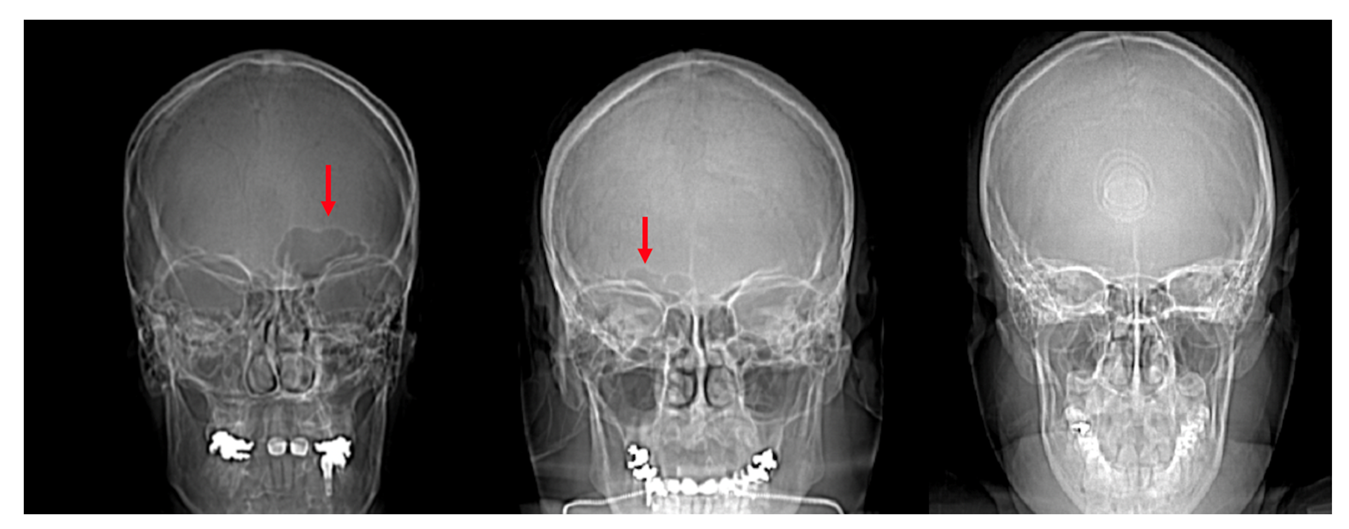
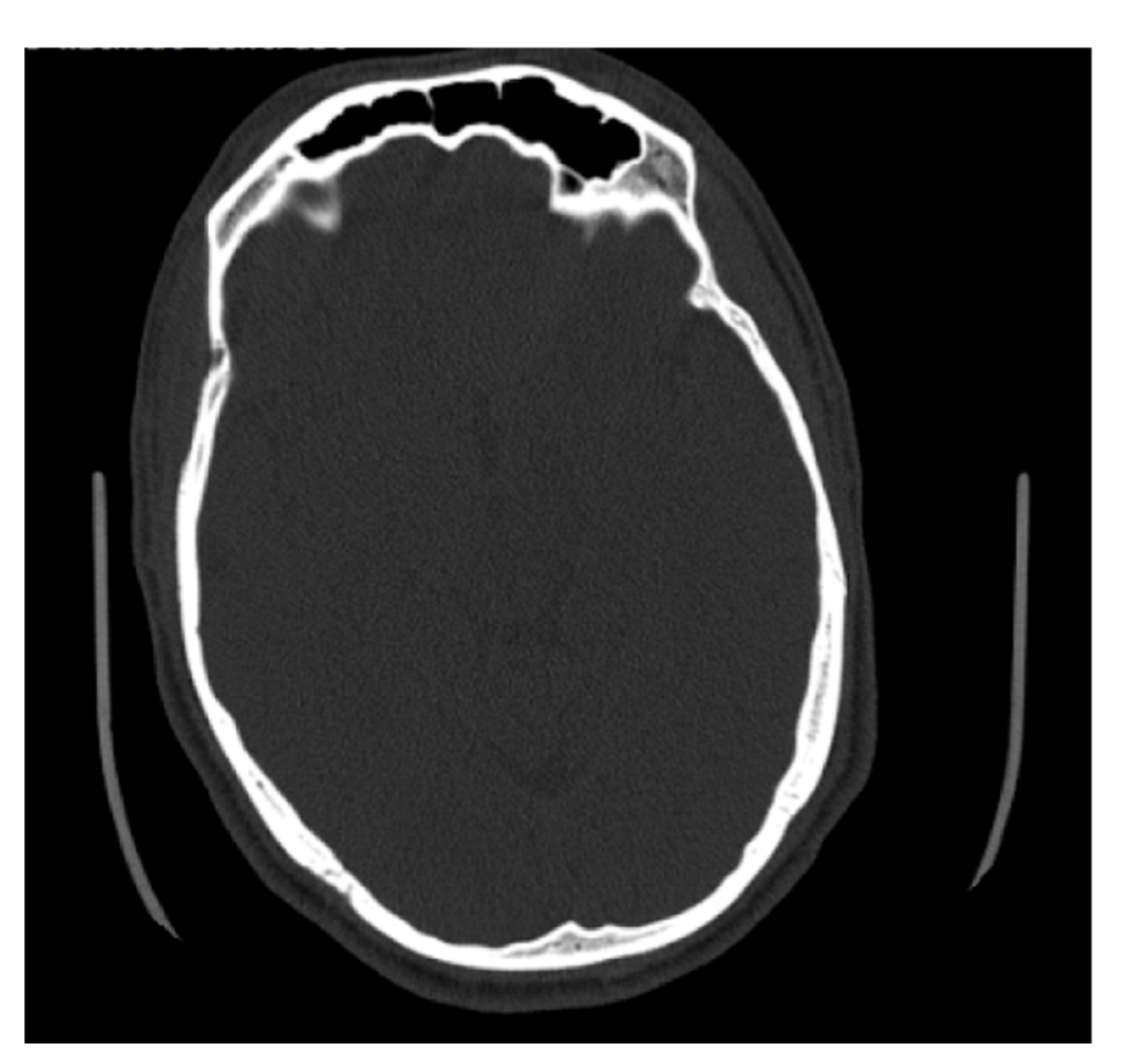
4.1. Frontal Sinus Absence and Unilateral Expression
4.2. Shape Variation
4.3. Dimensional Variation
4.4. Interactive Effects of Sexual Dimorphism and Ancestral Adaptions
4.5. The Use of CT Scans and Image Orientation/Quality
4.6. Limitations of Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bush, M.A.; Bush, P.J.; Sheets, H.D. Statistical evidence for the similarity of the human dentition. J. Forensic Sci. 2011, 56, 118–123. [Google Scholar] [CrossRef] [PubMed]
- Christensen, A.M. Testing the reliability of frontal sinuses in positive identification. J. Forensic Sci. 2005, 50, 1–5. [Google Scholar] [CrossRef]
- Christensen, A.M.; Hatch, G.M. Advances in the Use of Frontal Sinuses for Human Identification. In New Perspectives in Forensic Human Skeletal Identification; Academic Press: Cambridge, MA, USA, 2018. [Google Scholar]
- Da Silva, R.F.; Prado, F.B.; Caputo, I.G.C.; Devito, K.L.; de Luscena Botelho, T.; Júnior, E.D. The forensic importance of frontal sinus radiographs. J. Forensic Leg. Med. 2009, 16, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Flint, D.J.; Brent Dove, S.; Brumit, P.C.; White, M.; Senn, D.R. Computer-aided Dental Identification: An Objective Method for Assessment of Radiographic Image Similarity. J. Forensic Sci. 2009, 54, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Hurst, C.; Soler, A.; Fenton, T. Personal Identification in Forensic Anthropology. Encycl. Forensic Sci. 2013, 68–75. [Google Scholar] [CrossRef]
- Mann, R.W. Use of bone trabeculae to establish positive identification. Forensic Sci. Int. 1998, 98, 91–99. [Google Scholar] [CrossRef]
- Mundorff, A.Z.; Vidoli, G.; Melinek, J. Anthropological and Radiographic Comparison of Vertebrae for Identification of Decomposed Human Remains. J. Forensic Sci. 2006, 51, 1002–1004. [Google Scholar] [CrossRef] [PubMed]
- Nambiar, P.; Naidu, M.D.K.; Subramaniam, K. Anatomical variability of the frontal sinuses and their application in forensic identification. Clin. Anat. 1999, 12, 16–19. [Google Scholar] [CrossRef]
- Owsley, D.W.; Mann, R.W.; Chapman, R.E.; Moore, E.; Cox, W.A. Positive Identification in a Case of Intentional Extreme Fragmentation. J. Forensic Sci. 1993, 38, 985–996. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.P.; Santos, J.C. How to do identify single cases according to the quality assurance from IOFOS. The positive identification of an unidentified body by dental parameters: A case of homicide. J. Forensic Leg. Med. 2013, 20, 169–173. [Google Scholar] [CrossRef]
- Quatrehomme, G.; Fronty, P.; Sapanet, M.; Grévin, G.; Bailet, P.; Ollier, A. Identification by frontal sinus pattern in forensic anthropology. Forensic Sci. Int. 1996, 83, 147–153. [Google Scholar] [CrossRef]
- Reichs, K.J. Quantified comparison of frontal sinus patterns by means of computed tomography. Forensic Sci. Int. 1993, 61, 141–168. [Google Scholar] [CrossRef]
- Šlaus, M.; Strinović, D.; Pećina-Šlaus, N.; Brkić, H.; Baličević, D.; Petrovečki, V.; Pećina, T.C. Identification and analysis of human remains recovered from Wells from the 1991 war in Croatia. Forensic Sci. Int. 2007, 171, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Stephan, C.N.; Guyomarch, P. Quantification of Perspective-Induced Shape Change of Clavicles at Radiography and 3D Scanning to Assist Human Identification. J. Forensic Sci. 2013, 59, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Stephan, C.N.; Winburn, A.P.; Christensen, A.F.; Tyrrell, A.J. Skeletal identification BY radiographic Comparison: Blind tests of a Morphoscopic method using antemortem Chest Radiographs. J. Forensic Sci. 2011, 56, 320–332. [Google Scholar] [CrossRef]
- Tuceryan, M.; Li, F.; Blitzer, H.L.; Parks, E.T.; Platt, J.A. A framework for estimating probability of a match in forensic bite mark identification. J. Forensic Sci. 2011, 56, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela, A. Radiographic Comparison of the Lumbar Spine for Positive Identification of Human Remains. Am. J. Forensic Med. Pathol. 1997, 18, 215–217. [Google Scholar] [CrossRef]
- Yoshino, M.; Miyasaka, S.; Sato, H.; Seta, S. Classification system of frontal sinus patterns by radiography. Its application to identification of unknown skeletal remains. Forensic Sci. Int. 1987, 34, 289–299. [Google Scholar] [CrossRef]
- Zamora, A.C.; Tallman, S.D. The role of diffuse idiopathic skeletal hyperostosis (DISH) in positive identification. J. Forensic Sci. 2022, 67, 877–888. [Google Scholar] [CrossRef]
- Hatch, G.M.; Dedouit, F.; Christensen, A.M.; Thali, M.J.; Ruder, T.D. RADid: A pictorial review of radiologic identification using postmortem CT. J. Forensic Radiol. Imaging 2014, 2, 52–59. [Google Scholar] [CrossRef]
- Christensen, A.M. Assessing the Variation in Individual Frontal Sinus Outlines. Am. J. Phys. Anthropol. 2004, 127, 291–295. [Google Scholar] [CrossRef] [PubMed]
- Besana, J.L.; Rogers, T.L. Personal identification using the frontal sinus. J. Forensic Sci. 2010, 55, 584–589. [Google Scholar] [CrossRef]
- Baglivo, M.; Winklhofer, S.; Hatch, G.M.; Ampanozi, G.; Thali, M.J.; Ruder, T.D. The rise of forensic and post-mortem radiology—analysis of the literature between the year 2000 and 2011. J. Forensic Radiol. Imaging 2013, 1, 3–9. [Google Scholar] [CrossRef]
- Park, I.; Song, J.S.; Choi, H.; Kim, T.H.; Hoon, S.; Lee, S.H.; Lee, H.M. Volumetric study in the development of paranasal sinuses by CT imaging in Asian: A pilot study. Int. J. Pediatric Otorhinolaryngol. 2010, 74, 1347–1350. [Google Scholar] [CrossRef]
- Verma, S.; Mahima, V.G.; Patil, K. Radiomorphometric analysis of frontal sinus for sex determination. J. Forensic Dent. Sci. 2014, 6, 182–187. [Google Scholar]
- Aydinliogu, A.; Kavakli, A.; Erdem, S. Absence of Frontal Sinus in Turkish Individuals. Yonsei Med. J. 2003, 44, 215–218. [Google Scholar] [CrossRef] [Green Version]
- Cameriere, R.; Scendoni, R.; Lin, Z.; Milani, C.; Palacio, L.A.V.; Turiello, M.; Ferrante, L. Analysis of frontal sinuses for personal identification in a Chinese sample using new code number. J. Forensic Sci. 2019, 65, 45–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christensen, A.M. The Impact of Daubert: Implications for Testimony and Research in Forensic Anthropology (and the Use of Frontal Sinuses in Personal Identification). J. Forensic Sci. 2004, 49, 1–4. [Google Scholar] [CrossRef]
- Tang, J.; Hu, D.Y.; Jiang, F.H.; Yu, X.J. Assessing forensic applications of the frontal sinus in a Chinese Han population. Forensic Sci. Int. 2009, 183, 104.e1–104.e3. [Google Scholar] [CrossRef] [PubMed]
- Tubbs, R.S.; Elton, S.; Salter, G.; Blount, J.P.; Grabb, P.A.; Oakes, W.J. Superficial surgical landmarks for the frontal sinus. J. Neurosurg. 2002, 96, 320–322. [Google Scholar] [CrossRef] [PubMed]
- Sawatari, Y.; Caceres, J. Frontal Sinus Fractures. In Current Therapy in Oral and Maxillofacial Surgery; Shahrokh, R.B., Bell, R.B., Khan, H.A., Eds.; Elsevier Health Sciences: St. Louis, MO, USA, 2012; pp. 346–353. [Google Scholar]
- Buyuk, S.K.; Karaman, A.; Yasa, Y. Association between frontal sinus morphology and craniofacial parameters: A forensic view. J. Forensic Leg. Med. 2017, 49, 20–23. [Google Scholar] [CrossRef]
- Cox, M.; Malcom, M.; Fairgrieve, S.I. A new digital method for the objective comparison of frontal sinuses for identification. J. Forensic Sci. 2009, 54, 761–772. [Google Scholar] [CrossRef] [PubMed]
- Hamed, S.S.; El-Badrawy, A.M.; Fattah, S.A. Gender Identification from frontal sinus using multi-detector computed tomography. J. Forensic Radiol. Imaging 2014, 2, 117–120. [Google Scholar] [CrossRef]
- Hacl, A.; Costa, A.L.F.; Oliveira, J.M.; Tucunduva, M.J.; Girondi, J.R.; Nahás-Scocate, A.C.R. Three-dimenstional volumetric analysis of frontal sinus using medical software. J. Forensic Radiol. Imaging 2017, 11, 1–5. [Google Scholar] [CrossRef]
- Noback, M.L.; Samo, E.; van Leeuwen, C.H.; Lynnerup, N.; Harvati, K. Paranasal sinuses: A problematic proxy for climate adaptation in Neanderthals. J. Hum. Evol. 2016, 97, 176–179. [Google Scholar] [CrossRef] [PubMed]
- Rae, T.C.; Koppe, T.; Stringer, C.B. The Neanderthal face is not cold adapted. J. Hum. Evol. 2011, 60, 234–239. [Google Scholar] [CrossRef]
- Arslan, H.; Aydınlıoğlu, A.; Bozkurt, M.; Egeli, E. Anatomic variations of the paranasal sinuses: CT examination for endoscopic sinus surgery. Auris Nasus Larynx 1999, 26, 39–48. [Google Scholar] [CrossRef]
- Taylor, C.R.; Robertshaw, D.; Hofmann, R. Thermal panting: A comparison of wildebeest and zebu cattle. Am. J. Physiol. Leg. Content 1969, 217, 907–910. [Google Scholar] [CrossRef] [Green Version]
- Robertshaw, D. Mechanisms for the control of respiratory evaporative heat loss in panting animals. J. Appl. Physiol. 2006, 101, 664–668. [Google Scholar] [CrossRef]
- Eichenholz, A. Respiratory alkalosis. Arch. Intern. Med. 1965, 11, 699–708. [Google Scholar] [CrossRef]
- Holton, N.E.; Yokley, T.R.; Franciscus, R.S. Climatic adaptation and Neanderthal facial evolution: A comment on Rae et al. (2011). J. Hum. Evol. 2011, 61, 624–627. [Google Scholar] [CrossRef]
- Lee, M.K.; Osamu, S.; Spiegel, J.H. CT measurement of the frontal sinus—Gender differences and implications for frontal cranioplasty. J. Cranio-Maxillo-Facial Surg. 2010, 38, 494–500. [Google Scholar] [CrossRef]
- Kullman, L.; Eklund, E.; Grundin, R. The value of frontal sinus in identification of unknown persons. J. Forensic Odonstostomatol. 1900, 8, 3–10. [Google Scholar]
- Suman, J.L.; Jaisanghar, N.; Elangovan, S.; Mahaboob, N.; Senthilkumar, B.; Yoithapprabhunath, T.R.; Srichinthu, K.K. Configuration of frontal sinuses: A forensic perspective. J. Pharm. BioAllied Sci. 2016, 8 (Suppl. 1), S90–S95. [Google Scholar] [PubMed]
- Butaric, L.N.; Richman, A.; Garvin, H.M. The Effects of Cranial Orientation on Forensic Frontal Sinus Identification as Assessed by Outline Analyses. Biology 2022, 11, 62. [Google Scholar] [CrossRef] [PubMed]
- Christensen, A.M.; Crowder, C.M.; Ousley, S.D.; Houck, M.M. Error and its Meaning in Forensic Science. J. Forensic Sci. 2013, 59, 123–126. [Google Scholar] [CrossRef]
- Caple, J.M.; Byrd, J.E.; Stephan, C.N. The utility of elliptical Fourier analysis for estimating ancestry and sex from lateral skull photographs. Forensic Sci. Int. 2018, 289, 352–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuhl, F.; Giardina, C. Elliptic Fourier features of a closed contour. Comput. Graph. Image Process 1982, 18, 236–258. [Google Scholar] [CrossRef]
- Lestrel, P.E. Method for analyzing complex two-dimensional forms: Elliptical Fourier functions. Am. J. Hum. Biol. 1989, 1, 149–164. [Google Scholar] [CrossRef]
- Crampton, J.S. Elliptic Fourier shape analysis of fossil bivalves: Some practical considerations. Lethaia 1995, 28, 179–186. [Google Scholar] [CrossRef]
- Yang, H.P.; Ma, C.S.; Wen, H.; Zhan, Q.B.; Wang, X.L. A tool for developing an automatic insect identification system based on wing outlines. Sci. Rep. 2015, 5, 12786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ioviţă, R. Comparing stone tool resharpening trajectories with the aid of elliptical Fourier analysis. In New Perspectives on Old Stones; Springer: New York, NY, USA, 2010; pp. 235–253. [Google Scholar]
- Rohlf, F.; Archie, J. A comparison of Fourier methods for the description of wing shape in mosquitoes (Diptera:Culicidae). Syst. Zool. 1984, 33, 302–317. [Google Scholar] [CrossRef]
- Tracey, S.R.; Lyle, J.M.; Duhamel, G. Application of elliptical Fourier analysis of otolith form as a tool for stock identification. Fish. Res. 2006, 77, 138–147. [Google Scholar] [CrossRef]
- Richards, R.A.; Esteves, C. Stock-specific variation in scale morphology of Atlantic striped bass. Trans. Am. Fish. Soc. 1997, 126, 908–918. [Google Scholar] [CrossRef]
- Chen, S.Y.; Lestrel, P.E.; Kerr, W.J.S.; McColl, J.H. Describing shape changes in the human mandible using elliptical Fourier functions. Eur. J. Orthod. 2000, 22, 205–216. [Google Scholar] [CrossRef]
- Daegling, D.J.; Jungers, W.L. Elliptical Fourier analysis of symphyseal shape in great ape mandibles. J. Hum. Evol. 2000, 39, 107–122. [Google Scholar] [CrossRef] [PubMed]
- Ferrario, V.F.; Sforza, C.; Schmitz, J.H.; Miani, A.; Taroni, G. Fourier analysis of human soft tissue facial shape: Sex differences in normal adults. J. Anat. 1995, 187 Pt 3 Pt 3, 593. [Google Scholar]
- Blanton, A.I. Examining Distal Humerus Morphological Variation in Thai Individuals Using Elliptical Fourier Analysis. Ph.D. Thesis, Boston University, Boston, MA, USA, 2020. [Google Scholar]
- Schmittbuhl, M.; Le Minor, J.M.; Taroni, F.; Mangin, P. Sexual dimorphism of the human mandible: Demonstration by elliptical Fourier analysis. Int. J. Leg. Med. 2001, 115, 100–101. [Google Scholar] [CrossRef]
- Lestrel, P.E. Fourier Descriptors and Their Applications in Biology; Cambridge University Press: Cambridge, UK, 1997. [Google Scholar]
- Lestrel, P.E.; Takahashi, O.; Kanazawa, E. A quantitative approach for measuring crowding in the dental arch: Fourier descriptors. Am. J. Orthod. Dentofac. Orthop. 2004, 125, 716–725. [Google Scholar] [CrossRef]
- McDowell, J.L.; L’Abbé, E.N.; Kenyhercz, M.W. Nasal aperture shape evaluation between black and white South Africans. Forensic Sci. Int. 2012, 222, 397.e1–397.e6. [Google Scholar] [CrossRef] [PubMed]
- Sheridan, C.S.; Thomas, C.L.; Clement, J.G. Quantification of ethnic differences in facial profile. Aust. Orthod. J. 1997, 14, 218–224. [Google Scholar] [PubMed]
- Urbanová, P. Variation of the orbital rim using elliptic fourier analysis. In Biological Shape Analysis; World Scientific: Tsukuba, Japan, 2011; pp. 221–241. [Google Scholar]
- Fancourt, H.S.; Lynch, J.J.; Byrd, J.E.; Stephan, C.N. Next-generation osteometric sorting: Using 3D shape, elliptical Fourier analysis, and Hausdorff distance to optimize osteological pair-matching. J. Forensic Sci. 2021, 66, 821–836. [Google Scholar] [CrossRef] [PubMed]
- Paolello, J.; Cabo-Perez, L. Elliptical Fourier analysis of vertebral outlines for victim identification. In Proceedings of the American Academy of Forensic Sciences 60th Annual Scientific Meeting, Washington, DC, USA, 18–23 February 2008. [Google Scholar]
- Sanders, W.J.; Bodenbender, B.E. Morphometric analysis of lumbar vertebra UMP 67-28: Implications for spinal function and phylogeny of the Miocene Moroto hominoid. J. Hum. Evol. 1994, 26, 203–237. [Google Scholar] [CrossRef] [Green Version]
- Winburn, A.P.; Algee-Hewitt, B. Evaluating population affinity estimates in forensic anthropology: Insights from the forensic anthropology database for assessing methods accuracy (FADAMA). J. Forensic Sci. 2021, 66, 1210–1219. [Google Scholar] [CrossRef]
- DiGangi, E.A.; Bethard, J.D. Uncloaking a lost cause: Decolonizing ancestry estimation in the United States. Am. J. Phys. Anthropol. 2021, 175, 422–436. [Google Scholar] [CrossRef] [PubMed]
- Bethard, J.D.; DiGangi, E.A. Moving beyond a lost cause: Forensic anthropology and ancestry estimates in the United States. J. Forensic Sci. 2020, 65, 1791. [Google Scholar] [CrossRef] [PubMed]
- Tallman, S.D.; Parr, N.M.; Winburn, A.P. Assumed differences; unquestioned typologies: The oversimplification of race and ancestry in forensic anthropology. Forensic Anthropol. 2021, 4, 73. [Google Scholar] [CrossRef]
- Kelley, S.R.; Tallman, S.D. Population-Inclusive Assigned-Sex-at-Birth Estimation from Skull Computed Tomography Scans. Forensic Sci. 2022, 2, 321–348. [Google Scholar] [CrossRef]
- Tallman, S.D.; Kincer, C.D.; Plemons, E.D. Centering transgender individuals in forensic anthropology and expanding binary sex estimation in casework and research. Forensic Anthropol. 2021, 5. [Google Scholar] [CrossRef]
- Iwata, H.; Ukai, Y. SHAPE: A computer program package for quantitative evaluation of biological shapes based on elliptic Fourier descriptors. J. Hered. 2002, 93, 384–385. [Google Scholar] [CrossRef] [Green Version]
- Iwata, H. Tutorial for SHAPE v1.3; N.A.R. Organization: Ibaraki, Japan, 2006. [Google Scholar]
- Çakur, B.; Sumbullu, M.A.; Durna, N.B. Aplasia and agenesis of the frontal sinus in Turkish individuals: A retrospective study using dental volumetric tomography. Int. J. Med. Sci. 2011, 8, 278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gadekar, N.B.; Kotrashetti, V.S.; Hosmani, J.; Nayak, R. Forensic application of frontal sinus measurement among the Indian population. J. Oral Maxillofac. Pathol. 2019, 23, 147. [Google Scholar] [PubMed]
- Goyal, M.; Acharya, A.B.; Sattur, A.P.; Naikmasur, V.G. Are frontal sinuses useful indicators of sex? J. Forensic Leg. Med. 2013, 20, 91–94. [Google Scholar] [CrossRef] [PubMed]
- Danesh-Sani, S.A.; Bavandi, R.; Esmaili, M. Frontal sinus agenesis using computed tomography. J. Craniofacial Surg. 2011, 22, e48–e51. [Google Scholar] [CrossRef] [PubMed]
- Harris, A.M.P.; Wood, R.E.; Nortjé, C.J.; Thomas, C.J. The fronal sinus: Forensic fingerprint?—A pilot study. J. Forensic Odontostomatol. 1987, 5, 9–15. [Google Scholar]
- Ubelaker, D.H. Positive Identification from the Radiographic Comparison of Frontal Sinus Patterns. In Human Identification: Case Studies in Forensic Anthropology; Rathbun, T.A., Buikstra, J., Eds.; Charles, C. Thomas: Springfield, IL, USA, 1984; pp. 399–411. [Google Scholar]
- Libersa, C.; Faber, M. Etude anatomo-radiologique du sinus frontal chez l’enfant. Lille Med. 1958, 3, 453. [Google Scholar]
- Buikstra, J.E.; Ubelaker, D.H. Standards for Data Collection from Human Skeletal Remains. Ark. Archeol. Surv. Res. Ser. 1994, 44, 20. [Google Scholar]
- Walker, P.L. Sexing skulls using discriminant function analysis of visually assessed traits. Am. J. Phys. Anthropol. 2008, 136, 39–50. [Google Scholar] [CrossRef]
- Tallman, S.D. Cranial nonmetric sexual dimorphism and sex estimation in East and Southeast Asian individuals. Forensic Anthropol. 2019, 2, 204–221. [Google Scholar] [CrossRef]
- Koertvelyessy, T. Relationships between the frontal sinus and climatic conditions: A skeletal approach to cold adaptation. Am. J. Phys. Anthropol. 1972, 37, 161–172. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, C. Ober die Verhaltnisse der Warmeokonomie der Thiere zu ihrer Grosse. Gottinger Stud. 1847, 3, 595–708. [Google Scholar]
- Allen, J.A. The influence of physical conditions in the genesis of species. Radic. Rev. 1877, 1, 108–140. [Google Scholar]
- Ruff, C.B. Climatic adaptation and hominid evolution: The thermoregulatory imperative. Evol. Anthropol. Issues News Rev. 1993, 2, 53–60. [Google Scholar] [CrossRef]
- Lazenby, R.; Smashnuk, A. Osteometric variation in the Inuit second metacarpal: A test of Allen’s Rule. Int. J. Osteoarchaeol. 1999, 9, 182–188. [Google Scholar] [CrossRef]
- Holliday, T.W. Postcranial evidence of cold adaptation in European Neandertals. Am. J. Phys. Anthropol. 1997, 104, 245–258. [Google Scholar] [CrossRef]
- Brun, C.N.; Christensen, A.M.; Kravarski, M.; Gorincour, G.; Schweitzer, W.; Thali, M.J.; Ruder, T.D. Comparative radiologic identification with standardized single CT images of the paranasal sinuses—Evaluation of inter-rater reliability. Forensic Sci. Int. 2017, 280, 81–86. [Google Scholar] [CrossRef]
- Veneziano, A. Indiana Bones [R Package]. 2021. Available online: https://github.com/AlessioVeneziano/IndianaBones (accessed on 20 October 2021).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Initial Sample n = 325 | |
|---|---|
AFAB n = 176
| AMAB n = 149
|
| Frontal sinuses absent n = 12 ↓ | |
AFAB n = 6
| AMAB n = 6
|
| Sample used in dimensional analyses n = 313 | |
AFAB n = 170
| AMAB n = 143
|
| Frontal sinuses not connected medially n = 6 ↓ | |
AFAB n = 3
| AMAB n = 3
|
| Sample used in morphological analyses n = 307 | |
AFAB n = 167
| AMAB n = 140
|
| Variables | Results | Statistical Significance |
|---|---|---|
| Assigned sex vs. maximum height | X2 = 290.88 df = 291 p-value = 0.4909 | Fail to reject null hypothesis |
| Assigned sex vs. maximum width | X2 = 310.99 df = 306 p-value = 0.4100 | Fail to reject null hypothesis |
| Assigned sex vs. maximum depth | X2 = 273.45 df = 274 p-value = 0.4980 | Fail to reject null hypothesis |
| Population affinity vs. maximum height | X2 = 879.17 df = 873 p-value = 0.4351 | Fail to reject null hypothesis |
| Population affinity vs. maximum width | X2 = 914.2 df = 918 p-value = 0.5292 | Fail to reject null hypothesis |
| Population affinity vs. maximum depth | X2 = 850.28 df = 822 p-value = 0.2401 | Fail to reject null hypothesis |
| Assigned sex vs. H × W × D | X2 = 313 df = 312 p-value = 0.4734 | Fail to reject null hypothesis |
| Population affinity vs. H × W × D | X2 = 939 df = 936 p-value = 0.4663 | Fail to reject null hypothesis |
| Variables | Results | Statistical Significance |
|---|---|---|
| Assigned sex and population affinity vs. maximum height | Residual Deviance = 9.499 df = 3 p-value = −1.0658 × 10−8 | Reject null hypothesis |
| Assigned sex and population affinity vs. maximum width | Residual Deviance = 418.7 df = 3 p-value = 0.223 | Fail to reject hypothesis |
| Assigned sex and population affinity vs. maximum depth | Residual Deviance = 9.499 df = 3 p-value = −1.0658 × 10−8 | Reject null hypothesis |
| Assigned sex and population affinity vs. H × W × D | Residual Deviance = 9.499 df = 3 p-value = −1.0658 × 10−8 | Reject null hypothesis |
| Variables | Results | Statistical Significance | Significant Adjusted p-Values |
|---|---|---|---|
| Assigned sex and population affinity vs. maximum height | Assigned sex as a factor p-value = 0.00157 | Reject null hypothesis | AFAB vs. AMAB: p-value = 0.0015679 |
| Population affinity as a factor p-value = 0.13536 | Fail to reject null hypothesis | - | |
| Assigned sex and population affinities as factors p-value = 0.01191 | Reject null hypothesis | African-derived AFAB vs. Latin-derived AMAB: p-value = 0.0296342; Asian-derived AFAB vs. African-derived AMAB: p-value = 0.0204191; Asian-derived AFAB vs. Latin-derived AMAB: p-value = 0.0008630 | |
| Assigned sex and population affinity vs. maximum depth | Assigned sex as a factor p-value = 4.3 × 10−10 | Reject null hypothesis | AFAB vs. AMAB: p-value = 4.3 × 10−10 |
| Population affinity as a factor p-value = 0.499 | Fail to reject null hypothesis | - | |
| Assigned sex and population affinities as factors p-value = 0.146 | Reject null hypothesis | African-derived AFAB vs. African-derived AMAB: p-value = 0.0059593; African-derived AFAB vs. Asian-derived AMAB: p-value = 0.0022128; African-derived AFAB vs. Latin-derived AMAB: p-value = 0.0000058; African-derived AFAB vs. European-derived AMAB: p-value = 0.0151888; Latin-derived AFAB vs. African-derived AMAB: p-value = 0.0303812; Asian-derived AFAB vs. Latin-derived AMAB: p-value = 0.0057280; Latin-derived AFAB vs. Asian-derived AMAB: p-value = 0.0135587; Latin-derived AFAB vs. Latin-derived AMAB: p-value = 0.0000928; European-derived AFAB vs. Latin-derived AMAB: p-value = 0.0055286 | |
| Assigned sex and population affinity vs. (H × W × D) | Assigned sex as a factor p-value = 3.25 × 10−5 | Reject null hypothesis | AFAB vs. AMAB: p-value = 0.0000325 |
| Population affinity as a factor p-value = 0.8747 | Fail to reject null hypothesis | - | |
| Assigned sex and population affinities as factors p-value = 0.0177 | Reject null hypothesis | African-derived AFAB vs. African-derived AMAB: p-value = 0.0033445; African-derived AFAB vs. Asian-derived AMAB: p-value = 0.0200277; African-derived AFAB vs. Latin-derived AMAB: p-value = 0.0117864; Asian-derived AFAB vs. African-derived AMAB: p-value = 0.0336140 |
| Maximum Height (in mm) | ||||
|---|---|---|---|---|
| Group | n | Mean | Range | Standard Deviation |
| African-derived AFABs | 42 | 22.98 a | 10.53–38.66 | 7.36 |
| African-derived AMABs | 27 | 28.44 b | 8.68–58.71 | 11.84 |
| African-derived AFABs and AMABs | 69 | 25.12 | 8.68–58.71 | 9.67 |
| Asian-derived AFABs | 43 | 21.13 b, c | 7.02–43.06 | 9.83 |
| Asian-derived AMABs | 29 | 27.53 | 11.53–58.81 | 10.28 |
| Asian-derived AFABS and AMABs | 72 | 23.71 | 7.02–58.81 | 10.43 |
| European-derived AFABs | 43 | 26.75 | 9.45–44.43 | 8.05 |
| European-derived AMABs | 46 | 24.68 | 12.58–41.48 | 7.05 |
| European-derived AFABs and AMABs | 89 | 25.68 | 9.45–44.43 | 7.58 |
| Latin American-derived AFABs | 40 | 25.52 | 9.31–44.83 | 8.09 |
| Latin American-derived AMABs | 43 | 29.21 a, c | 13.36–50.71 | 9.41 |
| Latin American-derived AFABs and AMABs | 83 | 27.43 | 9.31–50.71 | 8.94 |
| All AFABs | 168 | 24.08 d | 7.02–44.83 | 8.61 |
| All AMABs | 145 | 27.29 d | 8.68–58.81 | 9.53 |
| Maximum Width (in mm) | ||||
|---|---|---|---|---|
| Group | n | Mean | Range | Standard Deviation |
| African-derived AFABs | 42 | 50.08 | 14.89–79.39 | 15.99 |
| African-derived AMABs | 27 | 57.38 | 10.62–108.12 | 26.66 |
| African-derived AFABs and AMABs | 69 | 52.94 | 10.62–108.12 | 20.95 |
| Asian-derived AFABs | 43 | 49.89 | 9.95–98.15 | 23.07 |
| Asian-derived AMABs | 29 | 58.06 | 13.15–96.01 | 22.55 |
| Asian-derived AFABs and AMABs | 72 | 53.18 | 9.95–98.15 | 23.06 |
| European-derived AFABs | 43 | 58.53 | 20.67–114.02 | 21.48 |
| European-derived AMABs | 46 | 55.07 | 15.46–95.01 | 19.09 |
| European-derived AFABs and AMABs | 89 | 56.74 | 15.46–114.02 | 20.24 |
| Latin American-derived AFABs | 40 | 56.19 | 26.48–87.64 | 18.12 |
| Latin American-derived AMABs | 43 | 56.29 | 20.06–88.45 | 17.97 |
| Latin American-derived AFABs and AMABs | 83 | 56.24 | 20.06–88.45 | 17.93 |
| All AFABs | 168 | 53.65 | 9.95–114.02 | 20.10 |
| All AMABs | 145 | 56.46 | 10.62–108.12 | 20.89 |
| Maximum Depth (in mm) | ||||
|---|---|---|---|---|
| Group | n | Mean | Range | Standard Deviation |
| African-derived AFABs | 42 | 9.85 a, b, c, d | 5.69–15.04 | 2.49 |
| African-derived AMABs | 27 | 13.27 a, e | 5.53–25.61 | 5.28 |
| African-derived AFABs and AMABs | 69 | 11.19 | 5.53–25.61 | 4.15 |
| Asian-derived AFABs | 43 | 11.16 f | 3.65–23.19 | 4.98 |
| Asian-derived AMABs | 29 | 13.44 b, g | 8.29–21.78 | 4.01 |
| Asian-derived AFABs and AMABs | 72 | 12.08 | 3.65–23.19 | 4.72 |
| European-derived AFABs | 43 | 11.15 i | 4.36–18.04 | 3.15 |
| European-derived AMABs | 46 | 12.59 d | 8.78–22.47 | 3.10 |
| European-derived AFABs and AMABs | 89 | 11.90 | 4.36–22.47 | 3.19 |
| Latin American-derived AFABs | 40 | 10.27 e, g, h | 6.36–15.63 | 2.26 |
| Latin American-derived AMABs | 43 | 14.15 c, f, h, i | 7.02–28.26 | 4.18 |
| Latin American-derived AFABs and AMABs | 83 | 12.28 | 6.36–49.84 | 3.90 |
| All AFABs | 168 | 10.62 j | 3.65–23.19 | 3.43 |
| All AMABs | 145 | 13.35 j | 5.53–28.26 | 4.08 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shamlou, A.A.; Tallman, S.D. Frontal Sinus Morphological and Dimensional Variation as Seen on Computed Tomography Scans. Biology 2022, 11, 1145. https://doi.org/10.3390/biology11081145
Shamlou AA, Tallman SD. Frontal Sinus Morphological and Dimensional Variation as Seen on Computed Tomography Scans. Biology. 2022; 11(8):1145. https://doi.org/10.3390/biology11081145
Chicago/Turabian StyleShamlou, Austin A., and Sean D. Tallman. 2022. "Frontal Sinus Morphological and Dimensional Variation as Seen on Computed Tomography Scans" Biology 11, no. 8: 1145. https://doi.org/10.3390/biology11081145
APA StyleShamlou, A. A., & Tallman, S. D. (2022). Frontal Sinus Morphological and Dimensional Variation as Seen on Computed Tomography Scans. Biology, 11(8), 1145. https://doi.org/10.3390/biology11081145