
Quantitative Proteomics and Network Analysis of Differentially Expressed Proteins in Proteomes of Icefish Muscle Mitochondria Compared with Closely Related Red-Blooded Species

 , , , and
, , , and
Abstract
Simple Summary
Abstract
1. Introduction
2. Methodology
3. Results
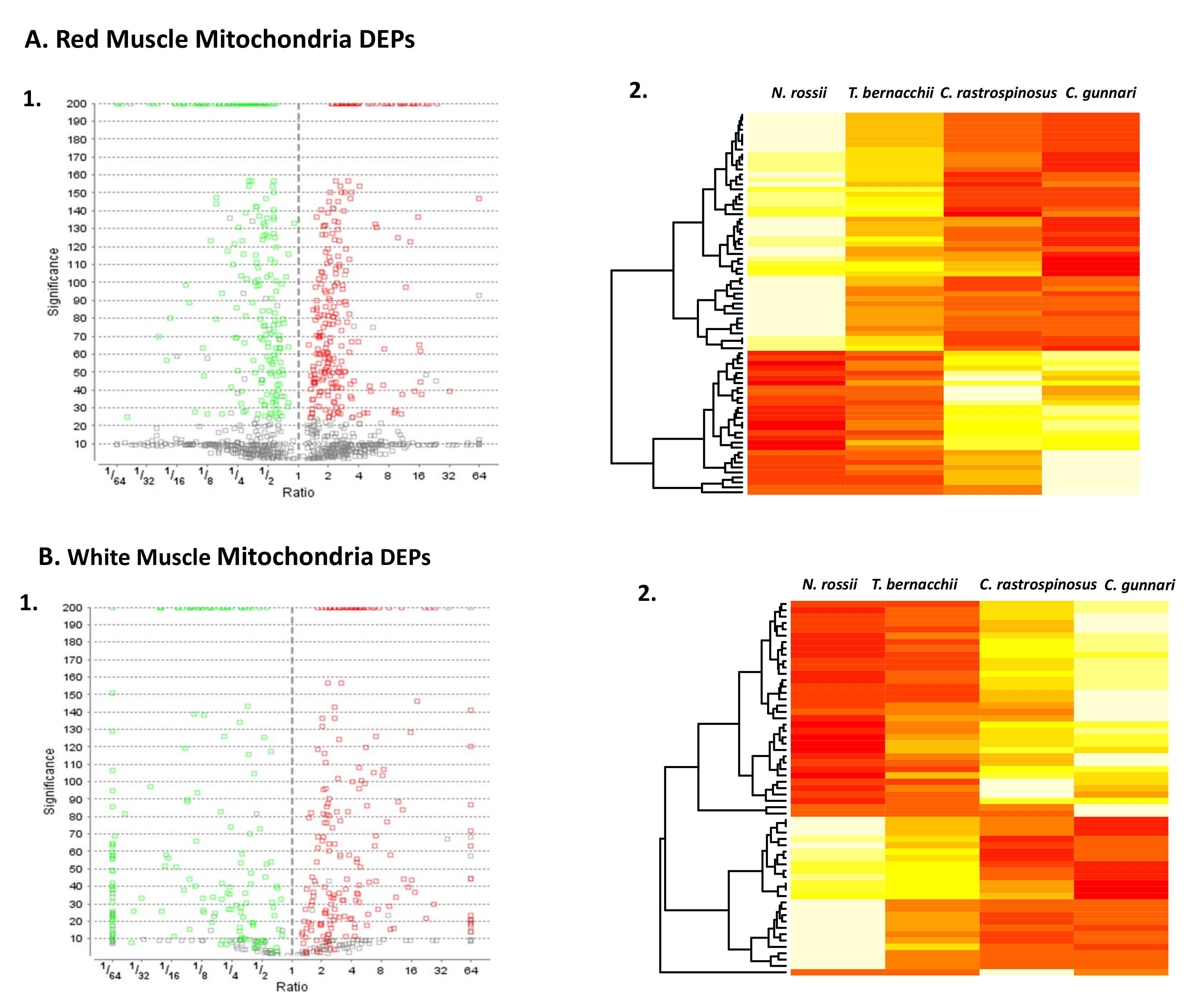
3.1. LC/LC-MS Data for RMM and WMM
3.2. Analysis of Proteins More in Abundance in Icefish
3.3. Proteins with Lower Abundance in Icefish
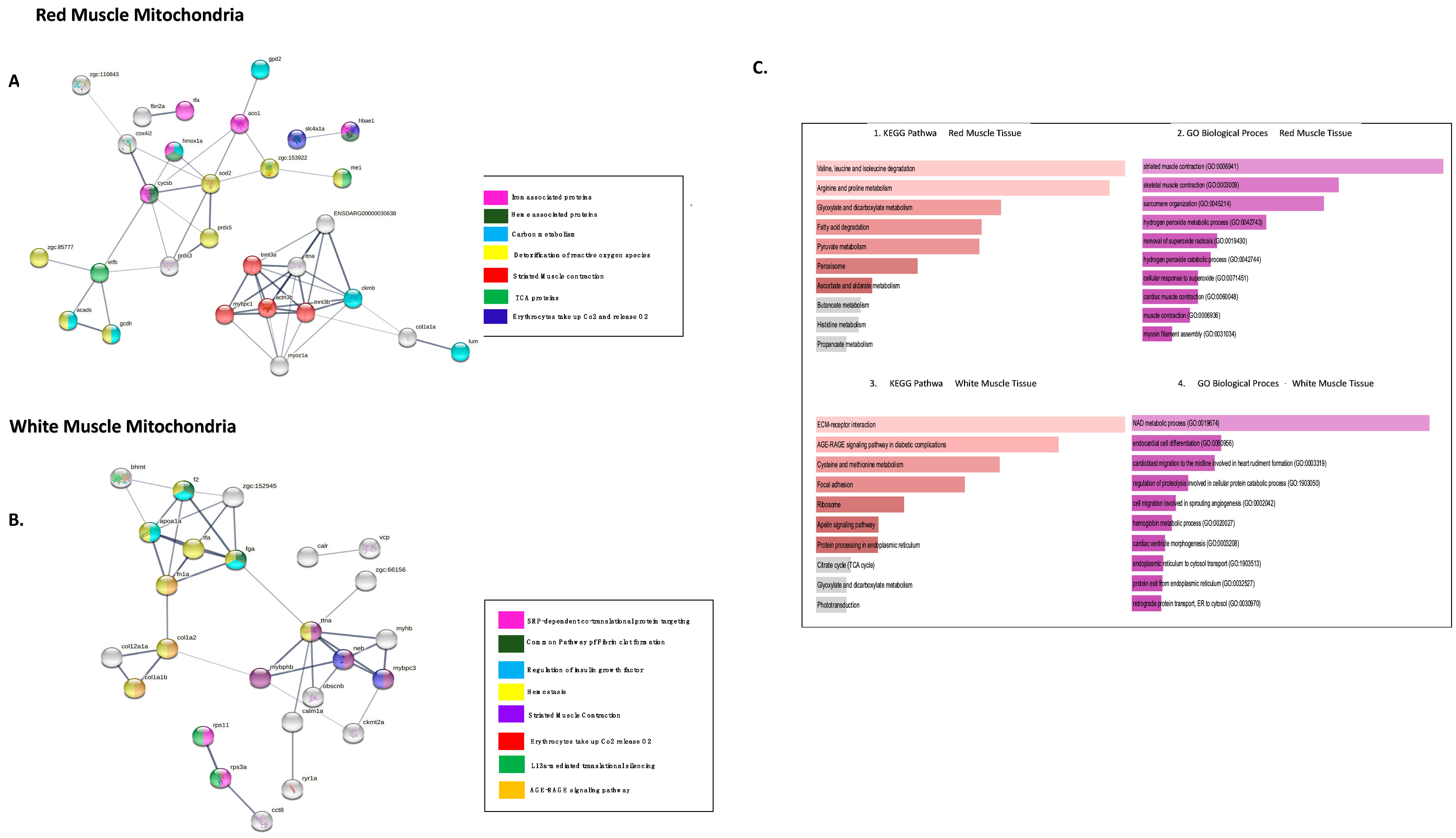
4. Discussion
4.1. Proteins Involved in Energy Metabolism
4.2. Muscle-Contraction Proteins
4.3. Redox
4.4. Haem-Associated Proteins
4.5. Ribosome Machinery in WMM
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Giorgi, C.; Agnoletto, C.; Bononi, A.; Bonora, M.; de Marchi, E.; Marchi, S.; Missiroli, S.; Patergnani, S.; Poletti, F.; Rimessi, A.; et al. Mitochondrial calcium homeostasis as potential target for mitochondrial medicine. Mitochondrion 2012, 12, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Murphy, M.P. How mitochondria produce reactive oxygen species. Biochem. J. 2009, 417, 1–13. [Google Scholar] [CrossRef]
- Tait, S.W.; Green, D.R. Mitochondrial regulation of cell death. Cold Spring Harb. Perspect. Biol. 2013, 53, a008706. [Google Scholar] [CrossRef] [PubMed]
- Chu, Q.; Martinez, T.F.; Novak, S.W.; Donaldson, C.J.; Tan, D.; Vaughan, J.M.; Chang, T.; Diedrich, J.K.; Andrade, L.; Kim, A.; et al. Regulation of the ER stress response by a mitochondrial microprotein. Nat. Commun. 2019, 10, 4883. [Google Scholar] [CrossRef] [PubMed]
- Azuma, K.; Ikeda, K.; Inoue, S. Functional mechanisms of mitochondrial respiratory chain supercomplex assembly factors and their involvement in muscle quality. Int. J. Mol. Sci. 2020, 21, 3182. [Google Scholar] [CrossRef]
- Guo, R.; Gu, J.; Zong, S.; Wu, M.; Yang, M. Structure and mechanism of mitochondrial electron transport chain. Biomed. J. 2018, 41, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Perutz, M.; Rossmann, M.; Cullis, A.F.; Muirhead, H.; Will, G.; North, A.C.T. Structure of hoemoglobin. Nature 1960, 185, 416–422. [Google Scholar] [CrossRef] [PubMed]
- Heinemann, I.U.; Jahn, M.; Jahn, D. The biochemistry of heme biosynthesis. Arch. Biochem. Biophys. 2008, 474, 238–251. [Google Scholar] [CrossRef]
- Straub, A.C.; Butcher, J.T.; Billaud, M.; Mutchler, S.M.; Artamonov, M.V.; Nguyen, A.T.; Johnson, T.; Best, A.K.; Miller, M.P.; Palmer, L.A.; et al. Hemoglobin α/eNOS coupling at myoendothelial junctions is required for nitric oxide scavenging during vasoconstriction. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 2594–2600. [Google Scholar] [CrossRef]
- Shephard, F.; Greville-Heygate, O.; Marsh, O.; Anderson, S.; Chakrabarti, L. A mitochondrial location for haemoglobins—Dynamic distribution in ageing and Parkinson’s disease. Mitochondrion 2014, 14, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Brown, N.; Alkhayer, K.; Clements, R.; Singhal, N.; Gregory, R.; Azzam, S.; Li, S.; Freeman, E.; McDonough, J. Neuronal Hemoglobin Expression and Its Relevance to Multiple Sclerosis Neuropathology. J. Mol. Neurosci. 2016, 59, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Lechauve, C.; Butcher, J.T.; Freiwan, A.; Biwer, L.A.; Keith, J.M.; Good, M.E.; Ackerman, H.; Tillman, H.S.; Kiger, L.; Isakson, B.E.; et al. Endothelial cell α-globin and its molecular chaperone α-hemoglobin-stabilizing protein regulate arteriolar contractility. J. Clin. Investig. 2018, 128, 5073–5082. [Google Scholar] [CrossRef]
- Corliss, B.A.; Delalio, L.J.; Stevenson Keller, T.C.; Keller, A.S.; Keller, D.A.; Corliss, B.H.; Beers, J.M.; Peirce, S.M.; Isakson, B.E. Vascular Expression of Hemoglobin Alpha in Antarctic Icefish Supports Iron Limitation as Novel Evolutionary Driver. Front. Physiol. 2019, 10, 1389. [Google Scholar] [CrossRef]
- Chuang, J.Y.; Lee, C.W.; Shih, Y.H.; Yang, T.; Yu, L.; Kuo, Y.M. Interactions between amyloid-β and hemoglobin: Implications for amyloid plaque formation in alzheimer’s disease. PLoS ONE 2012, 7, e33120. [Google Scholar] [CrossRef]
- Shephard, F.; Greville-Heygate, O.; Liddell, S.; Emes, R.; Chakrabarti, L. Analysis of Mitochondrial haemoglobin in Parkinson’s disease brain. Mitochondrion 2016, 29, 45–52. [Google Scholar] [CrossRef]
- Archer, S.D.; Johnston, I.A. Density of Cristae and Distribution of Mitochondria in the Slow Muscle Fibers of Antarctic Fish. Physiol. Zool. 1991, 64, 242–258. [Google Scholar] [CrossRef][Green Version]
- Duhamel, G.; Hulley, P.A.; Causse, R.; Koubbi, P.; Vacchi, M.; Pruvost, P.; Vigetta, S.; Irisson, J.-O.; Mormède, S.; Belchier, M.; et al. Chapter 7. Biogeographic Patterns of Fish. In Biogeographic Atlas of the Southern Ocean; Scientific Communitte on Antarctic Research: Cambridge, UK, 2014; ISBN 9780948277283. [Google Scholar]
- Johan, T. Ruud Vertebrates without Erythrocytes and Blood Pigment. Nature 1954, 173, 848–850. [Google Scholar]
- Kim, B.M.; Amores, A.; Kang, S.; Ahn, D.H.; Kim, J.H.; Kim, I.C.; Lee, J.H.; Lee, S.G.; Lee, H.; Lee, J.; et al. Antarctic blackfin icefish genome reveals adaptations to extreme environments. Nat. Ecol. Evol. 2019, 3, 469–478. [Google Scholar] [CrossRef] [PubMed]
- Beers, J.M.; Jayasundara, N. Antarctic notothenioid fish: What are the future consequences of “losses” and “gains” acquired during long-term evolution at cold and stable temperatures? J. Exp. Biol. 2015, 218, 1834–1845. [Google Scholar] [CrossRef] [PubMed]
- Di Prisco, G.; Cocca, E.; Parker, S.K.; Detrich, H.W. Tracking the evolutionary loss of hemoglobin expression by the white-blooded Antarctic icefishes. Gene 2002, 295, 185–191. [Google Scholar] [CrossRef]
- Near, T.J.; Parker, S.K.; Detrich, H.W. A genomic fossil reveals key steps in hemoglobin loss by the Antarctic icefishes. Mol. Biol. Evol. 2006, 23, 2008–2016. [Google Scholar] [CrossRef] [PubMed]
- Scudiero, R.; Trinchella, F.; Riggio, M.; Parisi, E. Structure and expression of genes involved in transport and storage of iron in red-blooded and hemoglobin-less antarctic notothenioids. Gene 2007, 97, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Borley, K.A.; Sidell, B.D. Evolution of the myoglobin gene in Antarctic Icefishes (Channichthyidae). Polar Biol. 2011, 34, 659–665. [Google Scholar] [CrossRef]
- Wujcik, J.M.; Wang, G.; Eastman, J.T.; Sidell, B.D. Morphometry of retinal vasculature in Antarctic fishes is dependent upon the level of hemoglobin in circulation. J. Exp. Biol. 2007, 210, 815–824. [Google Scholar] [CrossRef] [PubMed][Green Version]
- O’Brien, K.M.; Mueller, I.A.; Orczewska, J.I.; Dullen, K.R.; Ortego, M. Hearts of some Antarctic fishes lack mitochondrial creatine kinase. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2014, 178, 30–36. [Google Scholar] [CrossRef]
- Sidell, B.D.; O’Brien, K.M. When bad things happen to good fish: The loss of hemoglobin and myoglobin expression in Antarctic icefishes. J. Exp. Biol. 2006, 209, 1791–1802. [Google Scholar] [CrossRef]
- Sidell, B.D. Intracellular oxygen diffusion: The roles of myoglobin and lipid at cold body temperature. J. Exp. Biol. 1998, 201, 1119–1128. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, K.M.; Mueller, I.A. The unique mitochondrial form and function of antarctic channichthyid icefishes. Integr. Comp. Biol. 2010, 50, 993–1008. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Zhou, Q.; He, J.; Jiang, Z.; Peng, C.; Tong, R.; Shi, J. Recent advances in the development of protein–protein interactions modulators: Mechanisms and clinical trials. Signal Transduct. Target. Ther. 2020, 5, 213. [Google Scholar] [CrossRef]
- Rachlin, J.; Cohen, D.D.; Cantor, C.; Kasif, S. Biological context networks: A mosaic view of the interactome. Mol. Syst. Biol. 2006, 2, 66. [Google Scholar] [CrossRef] [PubMed]
- Marín, M.; Esteban, F.J.; Ramírez-Rodrigo, H.; Ros, E.; Sáez-Lara, M.J. An integrative methodology based on protein-protein interaction networks for identification and functional annotation of disease-relevant genes applied to channelopathies. BMC Bioinformatics 2019, 20, 565. [Google Scholar] [CrossRef]
- Baldwin, K.M.; Winder, W.W.; Terjung, R.L.; Holloszy, J.O. Glycolytic enzymes in different types of skeletal muscle: Adaptation to exercise. Am. J. Physiol. Content 1973, 225, 962–966. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.C.; Dennis, R.G.; Baar, K. Cultured slow vs. fast skeletal muscle cells differ in physiology and responsiveness to stimulation. Am. J. Physiol. Cell Physiol. 2006, 291, 11–17. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Pruitt, K.D.; Hogue, C.W.; Groll, M.; Hartmann, C.; Huber, R.; Fields, S. Mcode. Nucleic Acids Res. 2001, 29, 137–140. [Google Scholar] [CrossRef]
- Scardoni, G.; Petterlini, M.; Laudanna, C. Analyzing biological network parameters with CentiScaPe. Bioinformatics 2009, 25, 2857–2859. [Google Scholar] [CrossRef]
- Franz, M.; Rodriguez, H.; Lopes, C.; Zuberi, K.; Montojo, J.; Bader, G.D.; Morris, Q. GeneMANIA update 2018. Nucleic Acids Res. 2018, 46, W60–W64. [Google Scholar] [CrossRef]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’ayan, A. Enrichr: Interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinformatics 2013, 14, 128. [Google Scholar] [CrossRef]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef]
- Glancy, B.; Balaban, R.S. Protein composition and function of red and white skeletal muscle mitochondria. Am. J. Physiol. Cell Physiol. 2011, 300, C1280–C1290. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein–protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed]
- Doncheva, N.T.; Assenov, Y.; Domingues, F.S.; Albrecht, M. Topological analysis and interactive visualization of biological networks and protein structures. Nat. Protoc. 2012, 7, 670–685. [Google Scholar] [CrossRef]
- Magnoni, L.J.; Crespo, D.; Ibarz, A.; Blasco, J.; Fernández-Borràs, J.; Planas, J.V. Effects of sustained swimming on the red and white muscle transcriptome of rainbow trout (Oncorhynchus mykiss) fed a carbohydrate-rich diet. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2013, 166, 510–521. [Google Scholar] [CrossRef]
- Raman, K.; Damaraju, N.; Joshi, G.K. The organisational structure of protein networks: Revisiting the centrality-lethality hypothesis. Syst. Synth. Biol. 2014, 8, 73–81. [Google Scholar] [CrossRef]
- He, X.; Zhang, J. Why do hubs tend to be essential in protein networks? PLoS Genet. 2006, 2, 0826–0834. [Google Scholar] [CrossRef]
- Zotenko, E.; Mestre, J.; O’Leary, D.P.; Przytycka, T.M. Why do hubs in the yeast protein interaction network tend to be essential: Reexamining the connection between the network topology and essentiality. PLoS Comput. Biol. 2008, 4, e1000140. [Google Scholar] [CrossRef]
- Devenish, R.J.; Prescott, M.; Boyle, G.M.; Nagley, P. The oligomycin axis of mitochondrial ATP synthase: OSCP and the proton channel. J. Bioenerg. Biomembr. 2000, 32, 507–515. [Google Scholar] [CrossRef]
- Yang, W.; Nagasawa, K.; Münch, C.; Xu, Y.; Satterstrom, K.; Jeong, S.; Hayes, S.D.; Jedrychowski, M.P.; Vyas, F.S.; Zaganjor, E.; et al. Mitochondrial Sirtuin Network Reveals Dynamic SIRT3-Dependent Deacetylation in Response to Membrane Depolarization. Cell 2016, 167, 985–1000.e21. [Google Scholar] [CrossRef]
- Okada, H.; Goldhaber, J.I.; Ross, R.S.; Okada, H.; Lai, N.C.; Kawaraguchi, Y.; Liao, P.; Copps, J.; Sugano, Y.; Okada-maeda, S.; et al. Integrins protect cardiomyocytes from ischemia/reperfusion injury Find the latest version : Integrins protect cardiomyocytes from ischemia/reperfusion injury. J. Clin. Investig. 2013, 123, 4294–4308. [Google Scholar] [CrossRef]
- Mueller, I.A.; Grim, J.M.; Beers, J.M.; Crockett, E.L.; O’Brien, K.M. Inter-relationship between mitochondrial function and susceptibility to oxidative stress in red-And white-blooded Antarctic notothenioid fishes. J. Exp. Biol. 2011, 214, 3732–3741. [Google Scholar] [CrossRef]
- Katyal, G.; Ebanks, B.; Lucassen, M.; Papetti, C. Sequence and structure comparison of ATP synthase F0 subunits 6 and 8 in notothenioid fish. PLoS ONE 2021, 16, e0245822. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Hulver, M.W.; McMillan, R.P.; Cline, M.A.; Gilbert, E.R. The pivotal role of pyruvate dehydrogenase kinases in metabolic flexibility. Nutr. Metab. 2014, 11, 10. [Google Scholar] [CrossRef] [PubMed]
- Guderley, H.; Storey, K.B.; Fields, J.H.A.; Hochachka, P.W. Pyruvate kinase from Oplophorus gracilirostris muscle. Comp. Biochem. Physiol. Part B Comp. Biochem. 1976, 55, 475–478. [Google Scholar] [CrossRef]
- Goodpaster, B.H.; Sparks, L.M. Metabolic Flexibility in Health and Disease. Cell Metab. 2017, 25, 1027–1036. [Google Scholar] [CrossRef] [PubMed]
- Feller, G.; Goessens, G.; Gerday, C.; Bassleer, R. Heart structure and ventricular ultrastructure of hemoglobin- and myoglobin-free icefish Channichthys rhinoceratus. Cell Tissue Res. 1985, 242, 669–676. [Google Scholar] [CrossRef] [PubMed]
- Feller, G.; Gerday, C. Metabolic pattern of the heart of haemoglobin- and myoglobin-free Antarctic fish Channichthys rhinoceratus. Polar Biol. 1987, 7, 225–229. [Google Scholar] [CrossRef]
- Feller, G.; Pauly, J.-P.; Smal, A.; O’Carra, P.; Gerday, C. The lactate dehydrogenase of the icefish heart: Biochemical adaptations to hypoxia tolerance. Biochim. Biophys. Acta-Protein Struct. Mol. Enzymol. 1991, 1079, 343–347. [Google Scholar] [CrossRef]
- Feller, G.; Gerday, C. Adaptations of the hemoglobinless Antarctic icefish (Channichthyidae) to hypoxia tolerance. Comp. Biochem. Physiol.-A Physiol. 1997, 118, 981–987. [Google Scholar] [CrossRef]
- Storey, K.B.; Storey, J.M. Metabolic rate depression in animals: Transcriptional and translational controls. Biol. Rev. Camb. Philos. Soc. 2004, 79, 207–233. [Google Scholar] [CrossRef]
- June, F.L. Volume 3, number 5 FEBS LETTERS June 1969. FEBS Lett. 2000, 3, 3–7. [Google Scholar]
- Hewitt, C.O.; Eszes, C.M.; Sessions, R.B.; Moreton, K.M.; Dafforn, T.R.; Takei, J.; Dempsey, C.E.; Clarke, A.R.; Holbrook, J.J. A general method for relieving substrate inhibition in lactate dehydrogenases. Protein Eng. 1999, 12, 491–496. [Google Scholar] [CrossRef] [PubMed]
- Soler, J.; de Arriaga, D.; Cadenas, Q.; Cadenas, E. Substrate inhibition of lactate dehydrogenase fromPhycomyces blakesleeanus: NADH dependence. Exp. Mycol. 1981, 5, 357–362. [Google Scholar] [CrossRef]
- Wallimann, T.; Wyss, M.; Brdiczka, D.; Nicolay, K.; Eppenberger, H.M. Intracellular compartmentation, structure and function of creatine kinase isoenzymes in tissues with high and fluctuating energy demands: The “phosphocreatine circuit” for cellular energy homeostasis. Biochem. J. 1992, 281, 21–40. [Google Scholar] [CrossRef] [PubMed]
- Albertson, R.C.; Yan, Y.L.; Titus, T.A.; Pisano, E.; Vacchi, M.; Yelick, P.C.; Detrich, H.W.; Postlethwait, J.H. Molecular pedomorphism underlies craniofacial skeletal evolution in Antarctic notothenioid fishes. BMC Evol. Biol. 2010, 10, 4. [Google Scholar] [CrossRef] [PubMed]
- Kraker, J.; Viswanathan, S.K.; Knöll, R.; Sadayappan, S. Recent advances in the molecular genetics of familial hypertrophic cardiomyopathy in South Asian descendants. Front. Physiol. 2016, 7, 499. [Google Scholar] [CrossRef] [PubMed]
- Witjas-Paalberends, E.R.; Güclü, A.; Germans, T.; Knaapen, P.; Harms, H.J.; Vermeer, A.M.C.; Christiaans, I.; Wilde, A.A.M.; DosRemedios, C.; Lammertsma, A.A.; et al. Gene-specific increase in the energetic cost of contraction in hypertrophic cardiomyopathy caused by thick filament mutations. Cardiovasc. Res. 2014, 103, 248–257. [Google Scholar] [CrossRef]
- Barefield, D.; Kumar, M.; de Tombe, P.P.; Sadayappan, S. Contractile dysfunction in a mouse model expressing a heterozygous MYBPC3 mutation associated with hypertrophic cardiomyopathy. Am. J. Physiol.-Heart Circ. Physiol. 2014, 306, 807–815. [Google Scholar] [CrossRef]
- Van Dijk, S.J.; Dooijes, D.; Dos Remedios, C.; Michels, M.; Lamers, J.M.J.; Winegrad, S.; Schlossarek, S.; Carrier, L.; Cate, F.J.T.; Stienen, G.J.M.; et al. Cardiac myosin-binding protein C mutations and hypertrophic ardiomyopathy haploinsufficiency, deranged phosphorylation, and cardiomyocyte dysfunction. Circulation 2009, 119, 1473–1483. [Google Scholar] [CrossRef]
- Dimitriadi, A.; Beis, D.; Arvanitidis, C.; Adriaens, D.; Koumoundouros, G. Developmental temperature has persistent, sexually dimorphic effects on zebrafish cardiac anatomy. Sci. Rep. 2018, 8, 8125. [Google Scholar] [CrossRef] [PubMed]
- Lynch, T.L.; Sivaguru, M.; Velayutham, M.; Cardounel, A.J.; Michels, M.; Barefield, D.; Govindan, S.; Dos Remedios, C.; Van Der Velden, J.; Sadayappan, S. Oxidative Stress in Dilated Cardiomyopathy Caused by MYBPC3 Mutation. Oxid. Med. Cell. Longev. 2015, 2015, 424751. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.S.; Proctor, D.N.; Geiger, P.C.; Sieck, G.C. Reserve capacity for ATP consumption during isometric contraction in human skeletal muscle fibers. J. Appl. Physiol. 2001, 90, 657–664. [Google Scholar] [CrossRef] [PubMed]
- Klitgaard, H.; Zhou, M.; Schiaffino, S.; Betto, R.; Salviati, G.; Saltin, B. Ageing alters the myosin heavy chain composition of single fibres from human skeletal muscle. Acta Physiol. Scand. 1990, 140, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Van Der Velden, J.; Tocchetti, C.G.; Varricchi, G.; Bianco, A.; Sequeira, V.; Hilfiker-Kleiner, D.; Hamdani, N.; Leite-Moreira, A.F.; Mayr, M.; Falcao-Pires, I.; et al. Metabolic changes in hypertrophic cardiomyopathies: Scientific update from the working group of myocardial function of the European Society of Cardiology. Cardiovasc. Res. 2018, 114, 1273–1280. [Google Scholar] [CrossRef] [PubMed]
- Marques, I.J.; Leito, J.T.D.; Spaink, H.P.; Testerink, J.; Jaspers, R.T.; Witte, F.; Van Den Berg, S.; Bagowski, C.P. Transcriptome analysis of the response to chronic constant hypoxia in zebrafish hearts. J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 2008, 178, 77–92. [Google Scholar] [CrossRef] [PubMed]
- Noormohammadi, A.; Khodakarami, A.; Gutierrez-Garcia, R.; Lee, H.J.; Koyuncu, S.; Konig, T.; Schindler, C.; Saez, I.; Fatima, A.; Dieterich, C.; et al. Somatic increase of CCT8 mimics proteostasis of human pluripotent stem cells and extends C. elegans lifespan. Nat. Commun. 2016, 7, 13649. [Google Scholar] [CrossRef] [PubMed]
- Brehme, M.; Voisine, C.; Rolland, T.; Wachi, S.; Soper, J.H.; Zhu, Y.; Orton, K.; Villella, A.; Garza, D.; Vidal, M.; et al. A chaperome subnetwork safeguards proteostasis in aging and neurodegenerative disease. Cell Rep. 2014, 9, 1135–1150. [Google Scholar] [CrossRef]
- Brocker, C.; Lassen, N.; Estey, T.; Pappa, A.; Cantore, M.; Orlova, V.V.; Chavakis, T.; Kavanagh, K.L.; Oppermann, U.; Vasiliou, V. Aldehyde dehydrogenase 7A1 (ALDH7A1) is a novel enzyme involved in cellular defense against hyperosmotic stress. J. Biol. Chem. 2010, 285, 18452–18463. [Google Scholar] [CrossRef]
- O’Brien, K.M.; Rix, A.S.; Egginton, S.; Farrell, A.P.; Crockett, E.L.; Schlauch, K.; Woolsey, R.; Hoffman, M.; Merriman, S. Cardiac mitochondrial metabolism may contribute to differences in thermal tolerance of red- and white-blooded Antarctic notothenioid fishes. J. Exp. Biol. 2018, 221, jeb177816. [Google Scholar] [CrossRef]
- Bilyk, K.T.; Zhuang, X.; Murphy, K.R.; Christina Cheng, C.H. A tale of two genes: Divergent evolutionary fate of haptoglobin and hemopexin in hemoglobinless Antarctic icefishes. J. Exp. Biol. 2019, 222, jeb188573. [Google Scholar] [CrossRef]
- Cai, J.; Yang, J.; Jones, D.P. Mitochondrial control of apoptosis: The role of cytochrome c. Biochim. Biophys. Acta-Bioenerg. 1998, 1366, 139–149. [Google Scholar] [CrossRef]
- Santucci, R.; Sinibaldi, F.; Cozza, P.; Polticelli, F.; Fiorucci, L. Cytochrome c: An extreme multifunctional protein with a key role in cell fate. Int. J. Biol. Macromol. 2019, 136, 1237–1246. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, K.M.; Skilbeck, C.; Sidell, B.D.; Egginton, S. Muscle fine structure may maintain the function of oxidative fibres in haemoglobinless Antarctic fishes. J. Exp. Biol. 2003, 206, 411–421. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Marchi, S.; Patergnani, S.; Pinton, P. The endoplasmic reticulum-mitochondria connection: One touch, multiple functions. Biochim. Biophys. Acta-Bioenerg. 2014, 1837, 461–469. [Google Scholar] [CrossRef]
- Bakker, C.E.; De Diego Otero, Y.; Bontekoe, C.; Raghoe, P.; Luteijn, T.; Hoogeveen, A.T.; Oostra, B.A.; Willemsen, R. Immunocytochemical and biochemical characterization of FMRP, FXR1P, and FXR2P in the mouse. Exp. Cell Res. 2000, 258, 162–170. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. No. | Protein Abundance—Higher in Icefish (Red Muscle Mitochondria) | Relative Percent of Total Ion Area (Mapped)—Converted from Total Sum of Ion Area to Relative Percent of Total Ion Area | ||||||
|---|---|---|---|---|---|---|---|---|
| Protein Name | Gene Name | Quant Significance H&B Multiple Test Corrected q-Value | N. rossii | T. bernacchii | C. rastrospinosus | C. gunnari | Accession Number | |
| 1 | 26S protease regulatory subunit 4 isoform X1 & X2 | psmc1a | 1.25 × 10−6 | 4.9 | 6.6 | 7 | 81.4 | XP_010780333.1 |
| 2 | Apolipoprotein B-100-like partial | apobb | 3.76 × 10−19 | 3.2 | 3.3 | 27.9 | 65.6 | XP_010781933.1 |
| 3 | NAD(P) transhydrogenase mitochondrial-like | nnt | 2.95 × 10−19 | 5.5 | 5.8 | 29.6 | 59.2 | XP_010786020.1 |
| 4 | Hyaluronan and proteoglycan link protein 1 | hapln1 | 7.26 × 10−20 | 9.1 | 5.2 | 30 | 55.7 | XP_010767902.1 |
| 5 | Sarcolemmal membrane-associated protein-like isoform X1 & X2 | slmapa | 9.73 × 10−3 | 15.2 | 15.7 | 24 | 45.1 | XP_010782086.1 |
| 6 | ADP/ATP translocase 2-like | slc25a5 | 1.03 × 10−18 | 13 | 6.2 | 36.5 | 44.3 | XP_010765274.1 |
| 7 | Dihydrolipoyllysine-residue acetyltransferase component of pyruvate dehydrogenase complex | dlat | 9.09 × 10−12 | 15.9 | 16.3 | 27.3 | 40.5 | XP_010773292.1 |
| 8 | Alpha-2-macroglobulin-like partial | a2ml | 1.23 × 10−4 | 14.7 | 22.1 | 23.1 | 40.1 | XP_010771939.1 |
| 9 | Inter-alpha-trypsin inhibitor heavy chain H4-like | itih3a.2 | 1.37 × 10−11 | 12.1 | 12.7 | 35.9 | 39.3 | XP_010793736.1 |
| 10 | Stress-70 protein mitochondrial-like | hspa9 | 1.58 × 10−5 | 20 | 18.3 | 33.7 | 28.1 | XP_010766277.1 |
| 11 | Epoxide hydrolase 1 | ephx1l | 3.79 × 10−20 | 10.2 | 21.6 | 31.1 | 37.1 | XP_010790338.1 |
| 12 | Sorting and assembly machinery component 50 homolog | samm50 | 1.65 × 10−19 | 17.4 | 8.9 | 37.3 | 36.4 | XP_010773400.1 |
| 13 | Calcium-binding mitochondrial carrier protein Aralar1 | slc25a12 | 1.21 × 10−19 | 23 | 7.8 | 32.8 | 36.4 | XP_010768357.1 |
| 14 | Long chain fatty acyl CoA synthetase | acsl1a | 5.88 × 10−7 | 2.6 | 19.1 | 42.1 | 36.1 | AAK07470.1 |
| 15 | Malonyl-CoA decarboxylase mitochondrial | mlycd | 3.73 × 10−20 | 10.9 | 18.7 | 34.7 | 35.6 | XP_010792494.1 |
| 16 | NADH dehydrogenase [ubiquinone] 1 beta subcomplex subunit 8 mitochondrial | ndufb8 | 6.625 × 10−7 | 13.1 | 19.5 | 31.9 | 35.6 | XP_010779169.1 |
| 17 | 60S ribosomal protein L27a isoform X1 & X2 | rpl27a | 5.17 × 10−20 | 10.7 | 14.8 | 38.9 | 35.6 | XP_010790259.1 |
| 18 | Fatty acid binding protein H8-isoform | fabp3 | 2.43 × 10−19 | 2.4 | 25.6 | 37.2 | 34.8 | AAC60356.1 |
| 19 | ATP-dependent 6-phosphofructokinase muscle type-like | pfkm | 2.02 × 10−10 | 22.3 | 12.5 | 31 | 34.2 | XP_010794434.1 |
| 20 | Carnitine/acylcarnitine carrier protein | slc25a20 | 1.2 × 10−19 | 20 | 9.9 | 36.2 | 33.9 | XP_010773584.1 |
| 21 | ATP synthase subunit O | atp5o | 4.06 × 10−8 | 24.6 | 13.4 | 29.1 | 32.9 | XP_010772138.1 |
| 22 | Proliferation-associated protein 2G4-like | pa2g4b | 4.31 × 10−3 | 19 | 21.8 | 27.3 | 32 | XP_010782747.1 |
| 23 | ATP synthase subunit g mitochondrial | atp5l | 2.26 × 10−7 | 23.1 | 10.6 | 34.4 | 31.9 | XP_010779232.1 |
| 24 | Voltage-dependent anion-selective channel protein 3 | vdac3 | 4.6 × 10−19 | 13.2 | 12.1 | 43.2 | 31.6 | XP_010782516.1 |
| 25 | ATP synthase subunit alpha mitochondrial | atp5fa1 | 7.02 × 10−9 | 22.2 | 14.9 | 31.7 | 31.2 | XP_010779868.1 |
| 26 | ATP synthase subunit beta mitochondrial | zgc:163069 | 1.07 × 10−8 | 24.7 | 13.4 | 31.1 | 30.8 | XP_010765728.1 |
| 27 | ATP synthase subunit gamma mitochondrial isoform X1 | atp5g | 2.53 × 10−14 | 18.3 | 10 | 41.5 | 30.1 | XP_010778067.1 |
| 28 | 60S ribosomal protein L35 | rpl35 | 2.93 × 10−15 | 14.9 | 15.2 | 40.2 | 29.6 | XP_010790499.1 |
| 29 | ATP synthase F(0) complex subunit B1 mitochondrial | atp5pb | 1.54 × 10−15 | 23 | 11.6 | 36 | 29.5 | XP_010786327.1 |
| 30 | ATP synthase subunit delta mitochondrial | atp5d | 6.63 × 10−3 | 18.8 | 23.4 | 28.4 | 29.4 | XP_010775450.1 |
| 31 | Prothrombin partial | f2 | 7.41 × 10−20 | 14.3 | 23.2 | 33.2 | 29.3 | XP_010786167.1 |
| 32 | ADP/ATP translocase 3 | slc25a6 | 5.06 × 10−12 | 23.5 | 13.6 | 33.9 | 29 | XP_010784438.1 |
| 33 | 60S ribosomal protein L18a-like | rpl18a | 3.97 × 10−8 | 18.2 | 18.8 | 34.1 | 28.9 | XP_010774792.1 |
| 34 | ATP synthase subunit d mitochondrial | atp5pd | 8.44 × 10−20 | 25.2 | 8.6 | 37.5 | 28.7 | XP_010766730.1 |
| 35 | Malate dehydrogenase cytoplasmic-like partial | mdh1aa | 7.88 × 10−16 | 13.2 | 16.9 | 42.1 | 27.8 | XP_010766317.1 |
| 36 | Cytochrome c oxidase subunit 5A mitochondrial isoform X2 | cox5a | 5.67 × 10−20 | 16.5 | 9.2 | 46.4 | 27.8 | XP_010766309.1 |
| 37 | 60S ribosomal protein L23 | rpl23 | 1.13 × 10−12 | 15.9 | 20.4 | 37 | 26.7 | XP_010783746.1 |
| 38 | Prohibitin | phb | 2.72 × 10−6 | 18.1 | 18.4 | 39.1 | 24.5 | XP_010773724.1 |
| 39 | Ubiquinone biosynthesis protein COQ9 | coq9 | 8.44 × 10−11 | 17.2 | 17.6 | 42.1 | 23 | XP_010793356.1 |
| 40 | 26S proteasome non-ATPase regulatory subunit 6 | psmd6 | 3.76 × 10−2 | 8.5 | 15 | 28.3 | 48.1 | XP_010773228.1 |
| 41 | Fragile X mental retardation syndrome-related protein 2 | fxr2 | 7.06 × 10−2 | 9.6 | 10.5 | 16.4 | 63.5 | XP_010770797.1 |
| 42 | Reticulon-4-interacting protein 1 homolog mitochondrial-like | rtn4ip1 | 1.04 × 10−1 | 19.8 | 20.5 | 28.7 | 31 | XP_010790805.1 |
| 43 | Kininogen-1 | kng1 | 1.28 × 10−1 | 13.7 | 16.6 | 25.8 | 43.8 | XP_010787469.1 |
| Protein Abundance—Higher in Icefish (White Muscle) | Relative Percent of Total Ion Area (Mapped)—Converted from Total Sum of Ion Area to Relative Percent of Total Ion Area | |||||||
|---|---|---|---|---|---|---|---|---|
| S. No. | Protein Name | Gene Name | Quant Significance H&B Multiple Test Corrected q-Value | N. rossii | T. bernacchii | C. rastrospinosus | C. gunnari | Accession Number |
| 1 | Myosin regulatory light chain 2 skeletal muscle isoform-like | mylpfb | 3.42 × 10−19 | 1 | 2.2 | 2.3 | 94.4 | XP_010770965.1 |
| 2 | Myozenin-1 isoform X1 & X2 | myoz1b | 1.54 × 10−7 | 0 | 7.8 | 13.3 | 78.9 | XP_010791910.1 |
| 3 | Heat shock protein beta 1 | hspb1 | 9.48 × 10−5 | 9.9 | 4.1 | 11.5 | 74.6 | XP_010788098.1 |
| 4 | 60S ribosomal protein L35 | rpl35 | 4.58 × 10−20 | 3.4 | 7.9 | 32.7 | 56 | XP_010790499.1 |
| 5 | 40S ribosomal protein S16 isoform X1 | rps16 | 9.29 × 10−3 | 12.3 | 16.9 | 17.9 | 52.8 | XP_010773777.1 |
| 6 | 60S ribosomal protein L7 | rpl7 | 1.08 × 10−6 | 6.9 | 7.8 | 33.1 | 52.3 | XP_010770361.1 |
| 7 | Fragile X mental retardation syndrome-related protein 2 | fxr2 | 6.55 × 10−4 | 4.6 | 11.6 | 32.2 | 51.6 | XP_010770797.1 |
| 8 | 40S ribosomal protein S12 | rps12 | 1.13 × 10−4 | 8.4 | 13.8 | 27 | 50.8 | XP_010783785.1 |
| 9 | 60S ribosomal protein L30 | rpl30 | 8.33 × 10−5 | 3.1 | 22.6 | 24 | 50.3 | XP_010765856.1 |
| 10 | 60S ribosomal protein L12 isoform X2 | rpl12 | 5.13 × 10−20 | 9.9 | 11.7 | 28.5 | 49.8 | XP_010779104.1 |
| 11 | Reticulon | rtn1a | 5.51 × 10−20 | 14.9 | 13 | 22.7 | 49.4 | XP_010790870.1 |
| 12 | Palladin-like | palld | 1.07 × 10−14 | 2.7 | 15.9 | 32.1 | 49.2 | XP_010785200.1 |
| 13 | 60S ribosomal protein L9 | rpl9 | 4.70 × 10−20 | 4.2 | 3.4 | 43.7 | 48.7 | XP_010776310.1 |
| 14 | AMP deaminase 1 isoform X1 & X2 | ampd1 | 1.65 × 10−19 | 18.6 | 10.9 | 22.8 | 48.4 | XP_010793467.1 |
| 15 | ADP/ATP translocase 3 | slc25a5 | 3.22 × 10−10 | 7.8 | 10.6 | 33.7 | 47.8 | XP_010784438.1 |
| 16 | 40S ribosomal protein S13 | rps13 | 9.87 × 10−4 | 13.1 | 10.6 | 29.1 | 47.3 | XP_010794693.1 |
| 17 | 40S ribosomal protein S8-like partial | rps8 | 2.91 × 10−10 | 9.1 | 11.8 | 33 | 46 | XP_010787537.1 |
| 18 | Alpha-actinin-2 | actn2 | 1.42 × 10−8 | 12.1 | 18.2 | 23.6 | 46 | XP_010791686.1 |
| 19 | Kelch-like protein 41b | klhl41b | 5.57 × 10−20 | 4.2 | 11.6 | 38.5 | 45.7 | XP_010791686.1 |
| 20 | 60S ribosomal protein L6 | rpl6 | 4.536 × 10−20 | 7 | 11.3 | 36.3 | 45.4 | XP_010774286.1 |
| 21 | 40S ribosomal protein S5 | rps5 | 7.75 × 10−4 | 17 | 12.2 | 25.5 | 45.3 | XP_010782543.1 |
| 22 | Cytochrome c oxidase subunit II | mt-co2 | 5.91 × 10−13 | 2.6 | 26.8 | 25.9 | 44.7 | XP_010783741.1 |
| 23 | 40S ribosomal protein S25 | rps25 | 6.64 × 10−3 | 13 | 14.5 | 28.4 | 44.2 | YP_004581500.1 |
| 24 | 60S ribosomal protein L11 | rpl11 | 4.98 × 10−20 | 15.6 | 11.7 | 29.1 | 43.7 | XP_010776714.1 |
| 25 | Voltage-dependent anion-selective channel protein 2 | vdac2 | 8.76 × 10−7 | 15.6 | 10.6 | 30.4 | 43.5 | XP_010779161.1 |
| 26 | 40S ribosomal protein S4 | rps4x | 6.41 × 10−20 | 7.5 | 16 | 33.6 | 43 | XP_010767141.1 |
| 27 | 40S ribosomal protein S2 | rps2 | 5.4 × 10−20 | 6.8 | 11.4 | 39 | 42.8 | XP_010792965.1 |
| 28 | 60S ribosomal protein L10a | rp10a | 2.26 × 10−14 | 14.8 | 15.1 | 29.7 | 40.4 | XP_010783756.1 |
| 29 | 40S ribosomal protein S9 | rps9 | 4.40 × 10−8 | 12.5 | 17.9 | 29.3 | 40.3 | XP_010791484.1 |
| 30 | ATP synthase subunit g | atp5l | 4.66 × 10−20 | 18.7 | 11.6 | 32.7 | 37 | XP_010786813.1 |
| 31 | Vinculin | vcla | 1.15 × 10−7 | 18.7 | 11.6 | 32.7 | 37 | XP_010794136.1 |
| 32 | 60S ribosomal protein L13 | rpl13 | 5.51 × 10−11 | 8.2 | 16 | 39.4 | 36.5 | XP_010787927.1 |
| 33 | Fructose-1 6-bisphosphatase isozyme 2-like | fbp2 | 5.63 × 10−20 | 9.8 | 26 | 28.1 | 36.1 | XP_010789836.1 |
| 34 | 40S ribosomal protein SA isoform X2 | rpsa | 6.10 × 10−20 | 10 | 17.7 | 37.1 | 35.3 | XP_010781656.1 |
| 35 | Succinyl-CoA ligase | suclg1 | 6.66 × 10−11 | 5.5 | 10.2 | 49.4 | 35 | XP_010768032.1 |
| 36 | Tubulin alpha chain-like isoform X1 & X2 | tuba8l2 | 4.49 × 10−4 | 11.3 | 23.9 | 29.9 | 34.9 | XP_010778226.1 |
| 37 | 60S ribosomal protein L23a | rpl23a | 5.29 × 10−8 | 3.5 | 9.8 | 56.9 | 34.8 | XP_010766070.1 |
| 38 | NADH-ubiquinone oxidoreductase | mt-nd1 | 8.21 × 10−7 | 3.5 | 9.8 | 51.9 | 34.7 | XP_010791811.1 |
| 39 | Alpha-enolase-like | eno1a | 4.27 × 10−4 | 13 | 22.3 | 30.3 | 34.5 | XP_010777506.1 |
| 40 | Isocitrate dehydrogenase | idh1 | 1.22 × 10−19 | 20.6 | 11 | 35.7 | 32.6 | XP_010765339.1 |
| 41 | 26S proteasome non-ATPase regulatory subunit 12 | psmd12 | 1.19 × 10−3 | 10 | 11.8 | 47.6 | 30.6 | XP_010791048.1 |
| 42 | ATP synthase F(0) complex subunit B1 | atp5pb | 2.06 × 10−10 | 19 | 6.4 | 44.2 | 30.2 | XP_010777584.1 |
| 43 | Peptidyl-prolyl cis-trans isomerase-like | pplb | 1.02 × 10−3 | 8 | 27.2 | 35.3 | 29.5 | XP_010786327.1 |
| 44 | Malate dehydrogenase | mdh1aa | 1.71 × 10−19 | 15.1 | 9 | 46.6 | 29.4 | XP_010790691.1 |
| 45 | ATP synthase subunit O | atp5o | 5.16 × 10−4 | 16.1 | 9.4 | 45.9 | 28.6 | XP_010780749.1 |
| 46 | NAD(P) transhydrogenase | nnt | 5.81 × 10−8 | 15.9 | 16.7 | 39 | 28.3 | XP_010772138.1 |
| 47 | Glycogen phosphorylase muscle form-like | pygma | 8.47 × 10−6 | 17.7 | 25.3 | 30 | 26.9 | XP_010776087.1 |
| 48 | Ubiquitin carboxyl-terminal hydrolase 5 isoform X1 | usp5 | 4.8 × 10−4 | 9.3 | 19.1 | 44.8 | 26.7 | XP_010788355.1 |
| 49 | Cytochrome b-c1 complex subunit 2 | uqcrfs1 | 4.32 × 10−4 | 11.7 | 10.4 | 52.9 | 25 | XP_010769500.1 |
| 50 | ATP synthase subunit gamma | atp5g | 1.41 × 10−12 | 13.4 | 12.3 | 49.6 | 24.7 | XP_010784571.1 |
| 51 | ATP synthase subunit beta | zgc:163069 | 9.98 × 10−16 | 17.5 | 12.4 | 46.2 | 23.8 | XP_010778067.1 |
| 52 | Pyruvate kinase PKM | pkma | 5.13 × 10−19 | 11 | 10.3 | 56 | 22.7 | XP_010765728.1 |
| 53 | Vitellogenin-1-like | vtg2 | 1.38 × 10−2 | 0 | 2 | 93.6 | 4.4 | XP_010766216.1 |
| 54 | Proteasome subunit alpha type-7-like | psma8 | 1.96 × 10−2 | 9.6 | 11.7 | 55.1 | 23.6 | XP_010779640.1 |
| 55 | Guanine nucleotide-binding protein subunit beta-2-like 1 | gnb2l1 | 1.57 × 10−2 | 13.2 | 21 | 29.6 | 36.2 | XP_010783619.1 |
| 56 | ATP-dependent 6-phosphofructokinase muscle type-like | pfkmb | 1.84 × 10−2 | 16.7 | 17.3 | 26.1 | 39.9 | XP_010780163.1 |
| 57 | Inter-alpha-trypsin inhibitor heavy chain H3-like | zgc:110377 | 1.92 × 10−2 | 0 | 14.4 | 46.5 | 39.1 | XP_010782695.1 |
| Protein Abundance—Lower in Icefish (Red Muscle) | Relative Percent of Total Ion Area (Mapped)—Converted from Total Sum of Ion Area to Relative Percent of Total Ion Area | |||||||
|---|---|---|---|---|---|---|---|---|
| S. No. | Protein Name | Gene Name | Quant Significance H&B Multiple Test Corrected q-Value | N. rossii | T. bernacchii | C. rastrospinosus | C. gunnari | Accession Number |
| 1 | Hemoglobin subunit alpha-1 | hbae1 | 1.88 × 10−19 | 60 | 38.7 | 0.8 | 0.5 | NP_001290227.1 |
| 2 | Calsequestrin-1 | casq1 | 6.08 × 10−20 | 33.3 | 44.4 | 18.6 | 3.7 | XP_010782377.1 |
| 3 | Perilipin-3 | plin3 | 1.53 × 10−19 | 48.8 | 35.6 | 11.1 | 4.5 | XP_010778108.1 |
| 4 | creatine kinase S-type mitochondrial-like | ckmt2a | 4.35 × 10−19 | 71.4 | 22.4 | 1.4 | 4.8 | XP_010772488.1 |
| 5 | Transferrin | tfa | 8.36 × 10−20 | 34 | 54.2 | 17.3 | 5.5 | CAL92189.1 |
| 6 | Dehydrogenase/reductase SDR family member 7C | dhrs7cb | 3.01 × 10−9 | 68.9 | 18.2 | 7.1 | 5.7 | XP_010784042.1 |
| 7 | Myosin-binding protein H-like | mybpha | 9.09 × 10-20 | 66.6 | 20.7 | 5.9 | 6.8 | XP_010764981.1 |
| 8 | Band 3 anion transport protein | slc4a1 | 1.91 × 10−8 | 39 | 52 | 2.1 | 6.9 | XP_010785995.1 |
| 9 | Cytochrome c oxidase subunit 4 isoform 2 mitochondrial-like | cox4i2 | 5.79 × 10−20 | 60.1 | 22.5 | 9.6 | 7.7 | XP_010770791.1 |
| 10 | Titin-like | ttna | 2.47 × 10−11 | 50.9 | 21.6 | 19.4 | 8.1 | XP_010787367.1 |
| 11 | Glutaryl-CoA dehydrogenase mitochondrial-like | gcdh | 4.92 × 10−20 | 34 | 28.8 | 28.2 | 9 | XP_010795730.1 |
| 12 | Fibrillin-1-like isoform X1 | fbn2a | 5.14 × 10−20 | 67.5 | 17.5 | 6 | 9 | XP_010767938.1 |
| 13 | heme oxygenase 2 | hmox1 | 1.18 × 10−4 | 48.5 | 20.6 | 21.3 | 9.7 | XP_010786435.1 |
| 14 | NADP-dependent malic enzyme | me1 | 1.29 × 10−19 | 33.9 | 40.3 | 16 | 9.8 | XP_010776993.1 |
| 15 | Pyruvate dehydrogenase phosphatase regulatory subunit | pdpr | 8.30 × 10−9 | 24.9 | 48.6 | 15.4 | 11.1 | XP_010773093.1 |
| 16 | Dolichyl-diphosphooligosaccharide--protein glycosyltransferase subunit 1 | rpn1 | 7.17 × 10−12 | 38.5 | 30.1 | 20.2 | 11.2 | XP_010777725.1 |
| 17 | CDGSH iron-sulfur domain-containing protein 1 | zgc:110843 | 4.54 × 10−20 | 41 | 36 | 11.6 | 11.5 | XP_010767760.1 |
| 18 | Glutamate dehydrogenase | gdh | 3.06 × 10−19 | 29.9 | 48.6 | 11.1 | 11.9 | P82264.1 |
| 19 | Thioredoxin-dependent peroxide reductase mitochondrial | prdx3 | 6.46 × 10−20 | 31.2 | 29.5 | 27.2 | 12.1 | XP_010779546.1 |
| 20 | malate dehydrogenase | mdh | 1.178 × 10−12 | 32.5 | 37 | 18.4 | 12.1 | XP_010765488.1 |
| 21 | Cytochrome c | cycsb | 5.55 × 10−20 | 42.8 | 28.7 | 16.1 | 12.5 | XP_010792793.1 |
| 22 | Troponin T fast skeletal muscle isoforms-like isoform X1 to X3 | tnnt3a | 4.89 × 10−20 | 37.2 | 29.6 | 20.5 | 12.7 | XP_010784864.1 |
| 23 | Superoxide dismutase [Mn] mitochondrial | sod2 | 1.01 × 10−10 | 42 | 33.4 | 11.5 | 13.1 | XP_010771234.1 |
| 24 | Carnitine O-acetyltransferase | crat | 6.46 × 10−13 | 29.1 | 38.8 | 19 | 13.1 | XP_010795330.1 |
| 25 | PDZ and LIM domain protein 7 | ENSDARG00000030638 | 1.44 × 10−8 | 45.5 | 21 | 20.3 | 13.3 | XP_010765699.1 |
| 26 | Protein FAM162B-like | fam162a | 4.13 × 10−4 | 41.5 | 32 | 13 | 13.6 | XP_010783349.1 |
| 27 | Aconitate hydratase mitochondrial | aco1 | 4.14 × 10−19 | 45.3 | 26.8 | 14 | 14 | XP_010781940.1 |
| 28 | Retinol dehydrogenase 13-like isoform X1 & X2 | wu:fd55e03 | 2.59 × 10−13 | 42.1 | 26.2 | 17.6 | 14.1 | XP_010791045.1 |
| 29 | Myosin-binding protein C slow-type isoform X1 to X17 | mybpc | 2.39 × 10−3 | 39.8 | 27.3 | 18.8 | 14.1 | XP_010774860.1 |
| 30 | Lumican | lum | 2.21 × 10−7 | 47.8 | 26.9 | 11.3 | 14.1 | XP_010795529.1 |
| 31 | Retinol dehydrogenase 13-like isoform X1 | rdh13 | 2.59 × 10−13 | 26.2 | 42.1 | 17.6 | 14.1 | XP_010791045.1 |
| 32 | PDZ and LIM domain protein 7-like isoform X2 | ENSDARG00000030638 | 4.70 × 10−20 | 40.6 | 26.1 | 18.9 | 14.4 | XP_010785930.1 |
| 33 | Glycogen phosphorylase | pygm | 7.81 × 10−20 | 30.6 | 44.1 | 10.9 | 14.4 | XP_010788472.1 |
| 34 | Alpha-aminoadipic semialdehyde dehydrogenase | aldh7a1 | 1.25 × 10−19 | 36.3 | 28.9 | 20 | 14.7 | XP_010772035.1 |
| 35 | Myozenin-1-like | myoz1a | 7.07 × 10−20 | 46.5 | 26.8 | 12 | 14.7 | XP_010764663.1 |
| 36 | Collagen alpha-1(I) chain-like | col1a1a | 9.40 × 10−20 | 52.6 | 27.3 | 5.3 | 14.8 | XP_010768975.1 |
| 37 | Peroxiredoxin-5 mitochondrial | prdx5 | 8.32 × 10−7 | 30.5 | 28.9 | 25.2 | 15.4 | XP_010783999.1 |
| 38 | Short-chain specific acyl-CoA dehydrogenase mitochondrial | acads | 4.54 × 10−8 | 32.5 | 26.7 | 25.1 | 15.8 | XP_010779541.1 |
| 39 | Troponin alpha-3 chain-like | tnnt3b | 1.56 × 10−19 | 36.9 | 35.9 | 11 | 16.1 | XP_010771394.1 |
| 40 | Creatine kinase M-type | ckmb | 4.11 × 10−5 | 34.8 | 25.6 | 23.3 | 16.3 | XP_010791917.1 |
| 41 | Glycerol-3-phosphate dehydrogenase mitochondrial | gpd2 | 1.13 × 10−10 | 42.6 | 23.5 | 17 | 16.9 | XP_010791177.1 |
| 42 | electron transfer flavoprotein subunit beta | etfb | 2.38 × 10−5 | 33.9 | 24.8 | 24.2 | 17.1 | XP_010791064.1 |
| 43 | 1 4-alpha-glucan-branching enzyme | gbe1b | 2.20 × 10−10 | 28.1 | 44.4 | 10.4 | 17.1 | XP_010775191.1 |
| 44 | Ubiquitin carboxyl-terminal hydrolase 5 isoform X1 & X2 | usp5 | 8.40 × 10−5 | 34.9 | 25.9 | 21.1 | 18 | XP_010769508.1 |
| 45 | Methylmalonyl-CoA mutase mitochondrial | mut | 2.48 × 10−4 | 33.7 | 26.7 | 21.5 | 18 | XP_010784587.1 |
| 46 | probable acyl-CoA dehydrogenase 6 | zgc:85777 | 1.82 × 10−3 | 31.3 | 26.9 | 23.2 | 18.6 | XP_010772948.1 |
| 47 | Alpha-actinin-3 | actn3b | 5.52 × 10−19 | 39.5 | 28.5 | 13.3 | 18.7 | XP_010784415.1 |
| 48 | Carbonic anhydrase 1 | ca1 | 7.88 × 10−20 | 49.4 | 20 | 10.9 | 19.6 | XP_010765900.1 |
| Protein Abundance—Lower in Icefish (White Muscle) | Relative Percent of Total Ion Area (Mapped)—Converted from Total Sum of Ion Area to Relative Percent of Total Ion Area | |||||||
|---|---|---|---|---|---|---|---|---|
| S. No. | Protein Name | Gene Name | Quant Significance H&B Multiple Test Corrected q-Value | N. rossii | T. bernacchii | C. rastrospinosus | C. gunnari | Accession Number |
| 1 | Calreticulin | calr | 5.72 × 10−3 | 75.4 | 4.7 | 19.8 | 0 | XP_010773398.1 |
| 2 | Creatine kinase S-type mitochondrial-like | ckmt2a | 1.19 × 10−19 | 78.2 | 14.8 | 6.4 | 0.7 | XP_010772488.1 |
| 3 | Myosin heavy chain fast skeletal 13 | zgc:66156 | 3.68 × 10−3 | 37.8 | 34.7 | 26.6 | 0.9 | XP_010791001.1 |
| 4 | Prothrombin partial | f2 | 7.77 × 10−4 | 37.3 | 30.3 | 31.2 | 1.1 | XP_010786167.1 |
| 5 | Fibronectin | fn1a | 1.33 × 10−5 | 33.1 | 30.2 | 34.9 | 1.8 | XP_010794764.1 |
| 6 | Transmembrane protease serine 2-like | LOC571565 | 6.81 × 10−3 | 81.2 | 16.8 | 0 | 2 | XP_010778161.1 |
| 7 | Hemoglobin subunit zeta | hbz | 2.98 × 10−4 | 80.5 | 16.4 | 0.3 | 2.5 | XP_010778322.1 |
| 8 | Fibrinogen alpha chain-like | fga | 2.67 × 10−5 | 51.4 | 39.8 | 5.2 | 2.8 | XP_010771898.1 |
| 9 | Alpha globin | hbaa1 | 7.77 × 10−20 | 79.6 | 16.5 | 0.4 | 3.3 | AAC25100.1 |
| 10 | Beta-globin | ba-1 | 7.45 × 10−9 | 87.5 | 9.1 | 0 | 3.4 | AAC60372.1 |
| 11 | Haemoglobin | hb | 7.33 × 10−9 | 87.5 | 9.1 | 0 | 3.4 | NP_001290226.1 |
| 12 | Ryanodine receptor 1- | ryr1a | 3.16 × 10−4 | 54.5 | 19.7 | 21.1 | 4.5 | XP_010787188.1 |
| 13 | Keratin type I cytoskeletal 19-like | zgc:153629 | 8.84 × 10−20 | 54.7 | 31.8 | 8.9 | 4.6 | XP_010787448.1 |
| 14 | Myosin heavy chain fast skeletal muscle-like | myhb | 4.58 × 10−12 | 46.7 | 27.6 | 19.2 | 6.5 | NP_001290213.1 |
| 15 | Malate dehydrogenase cytoplasmic | mdh1aa | 5.56 × 10−7 | 54.8 | 24.1 | 13.3 | 7.8 | XP_010765488.1 |
| 16 | Apolipoprotein A | apoa1 | 1.282 × 10−19 | 54.4 | 20 | 16.1 | 9.3 | XP_010792180.1 |
| 17 | Myomesin-1-like | myom1 | 1.31 × 10−19 | 31.1 | 29.8 | 21.8 | 17.3 | XP_010789743.1 |
| 18 | Betaine-homocysteine S-methyltransferase 1-like | bhmt | 7.32 × 10−20 | 53.1 | 20.8 | 14.4 | 11.6 | XP_010794476.1 |
| 19 | Collagen alpha-1(I) chain-like | col1a1b | 6.56 × 10−4 | 37.2 | 34.6 | 15.8 | 12.4 | XP_010768975.1 |
| 20 | Hemopexin | zgc:152945 | 3.14 × 10−6 | 40.7 | 26.6 | 19.7 | 13 | XP_010788340.1 |
| 21 | T-complex protein 1 subunit theta | cct8 | 3.34 × 10−6 | 60.6 | 18.2 | 7.4 | 13.9 | NP_001290219.1 |
| 22 | Transitional endoplasmic reticulum ATPase-like | vcp | 3.10 × 10−3 | 51.4 | 20.5 | 12.7 | 15.4 | XP_010770092.1 |
| 23 | 40S ribosomal protein S11 | rps11 | 8.261 × 10−4 | 43.2 | 32.7 | 8.7 | 15.5 | XP_010791578.1 |
| 24 | Titin-like partial | ttna | 1.66 × 10−3 | 32.9 | 28.7 | 21.1 | 17.2 | XP_010790363.1 |
| 25 | Calmodulin | calm1a | 1.197 × 10−10 | 36.2 | 30.9 | 14 | 18.9 | XP_010768524.1 |
| 26 | 40S ribosomal protein S3a | rps3a | 9.93 × 10−12 | 35.1 | 30.1 | 15.3 | 19.5 | XP_010773841.1 |
| 27 | Obscurin isoform X2 | obscnb | 2.1 × 10−2 | 36.8 | 30.2 | 14.9 | 18.1 | XP_010790854.1 |
| 28 | Transferrin | tfa | 3.14 × 10−2 | 36.3 | 33.5 | 22.7 | 7.6 | CAL92189.1 |
| 29 | Collagen alpha 1(XII) chain isoform X1, X2, X3, X4 & X5 | col12a1a | 8.53 × 10−2 | 68.8 | 18.2 | 8.5 | 4.5 | XP_010777236.1 |
| 30 | Nebulin-like isoform X4 | neb | 2.23 × 10−1 | 54.2 | 23.8 | 9.2 | 12.7 | XP_010772593.1 |
| 31 | Myosin-binding protein C slow-type isoform X1 to X17 | mybpc3 | 1.31 × 10−1 | 45.6 | 26.9 | 12.6 | 14.9 | XP_010774870.1 |
| 32 | Collagen alpha-2(I) chain isoform X1 | col1a2 | 2.91 × 10−1 | 54.6 | 20.8 | 9.6 | 14.9 | XP_010772950.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Katyal, G.; Ebanks, B.; Dowle, A.; Shephard, F.; Papetti, C.; Lucassen, M.; Chakrabarti, L. Quantitative Proteomics and Network Analysis of Differentially Expressed Proteins in Proteomes of Icefish Muscle Mitochondria Compared with Closely Related Red-Blooded Species. Biology 2022, 11, 1118. https://doi.org/10.3390/biology11081118
Katyal G, Ebanks B, Dowle A, Shephard F, Papetti C, Lucassen M, Chakrabarti L. Quantitative Proteomics and Network Analysis of Differentially Expressed Proteins in Proteomes of Icefish Muscle Mitochondria Compared with Closely Related Red-Blooded Species. Biology. 2022; 11(8):1118. https://doi.org/10.3390/biology11081118
Chicago/Turabian StyleKatyal, Gunjan, Brad Ebanks, Adam Dowle, Freya Shephard, Chiara Papetti, Magnus Lucassen, and Lisa Chakrabarti. 2022. "Quantitative Proteomics and Network Analysis of Differentially Expressed Proteins in Proteomes of Icefish Muscle Mitochondria Compared with Closely Related Red-Blooded Species" Biology 11, no. 8: 1118. https://doi.org/10.3390/biology11081118
APA StyleKatyal, G., Ebanks, B., Dowle, A., Shephard, F., Papetti, C., Lucassen, M., & Chakrabarti, L. (2022). Quantitative Proteomics and Network Analysis of Differentially Expressed Proteins in Proteomes of Icefish Muscle Mitochondria Compared with Closely Related Red-Blooded Species. Biology, 11(8), 1118. https://doi.org/10.3390/biology11081118