
Ecotourism Disturbance on an Endemic Endangered Primate in the Huangshan Man and the Biosphere Reserve of China: A Way to Move Forward

 , , , ,
, , , ,
Abstract
:Simple Summary
Abstract

1. Introduction
2. Materials and Methods
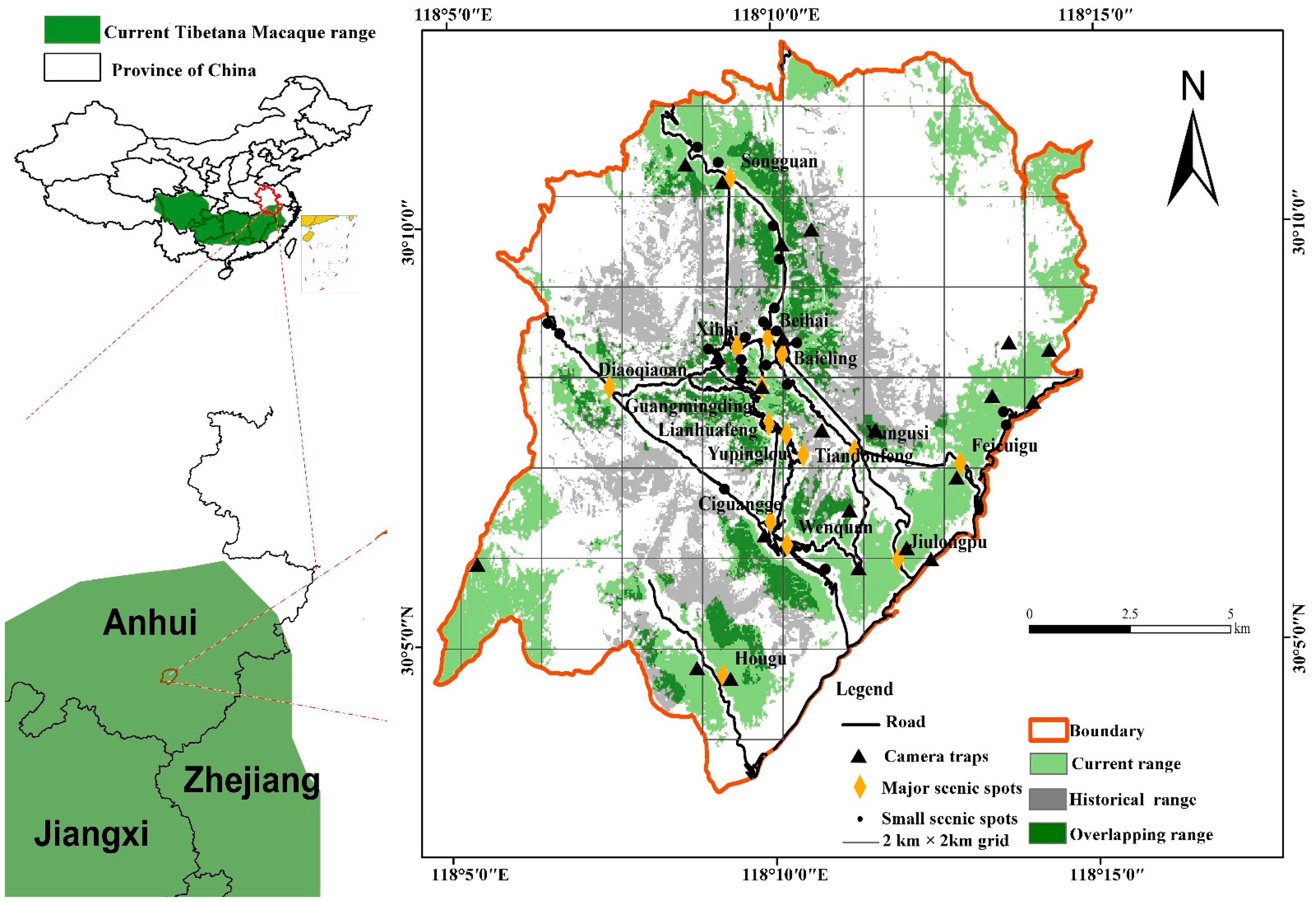
2.1. Study Site
2.2. Sampling Methods
- (1)
- Primate population distribution and density survey methods
- (2)
- Long-term macaque population dynamics
- (3)
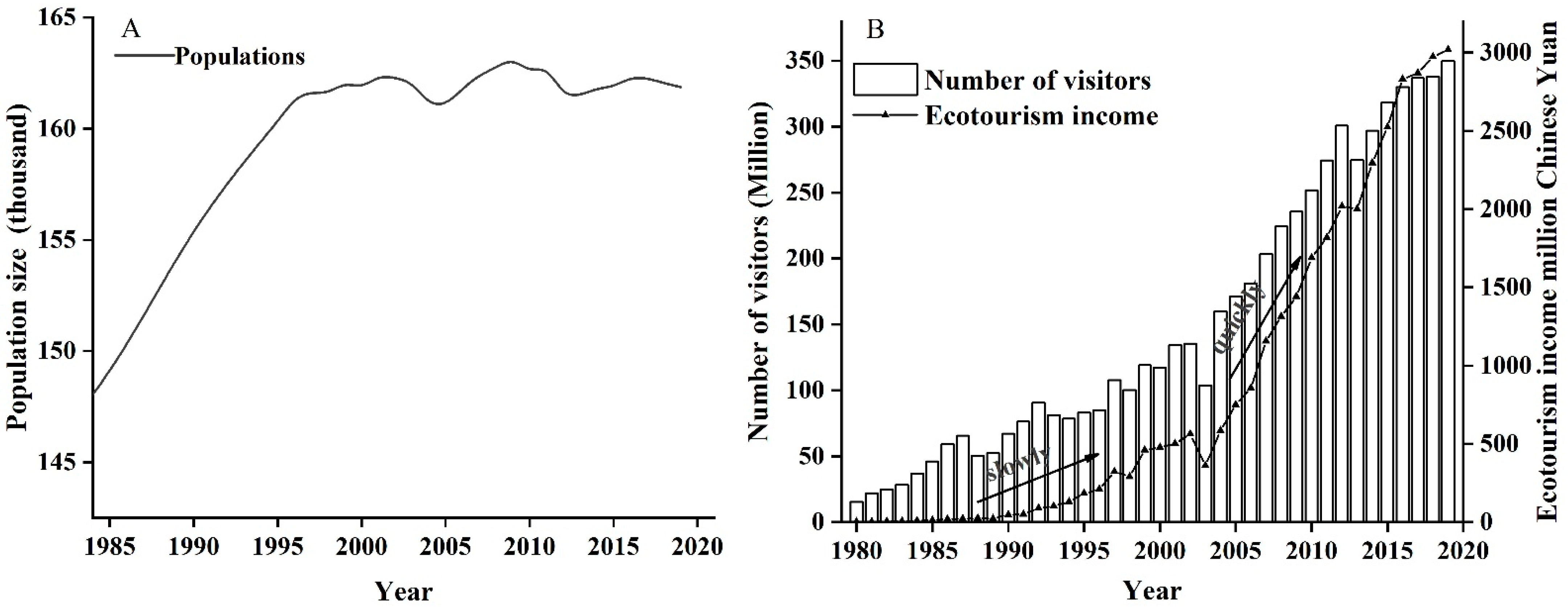
- Human population size, number of tourists, eco-tourism income, and GDP
2.3. Data Analysis and Modeling
- (1)
- Overall macaque population distribution and density
- (2)
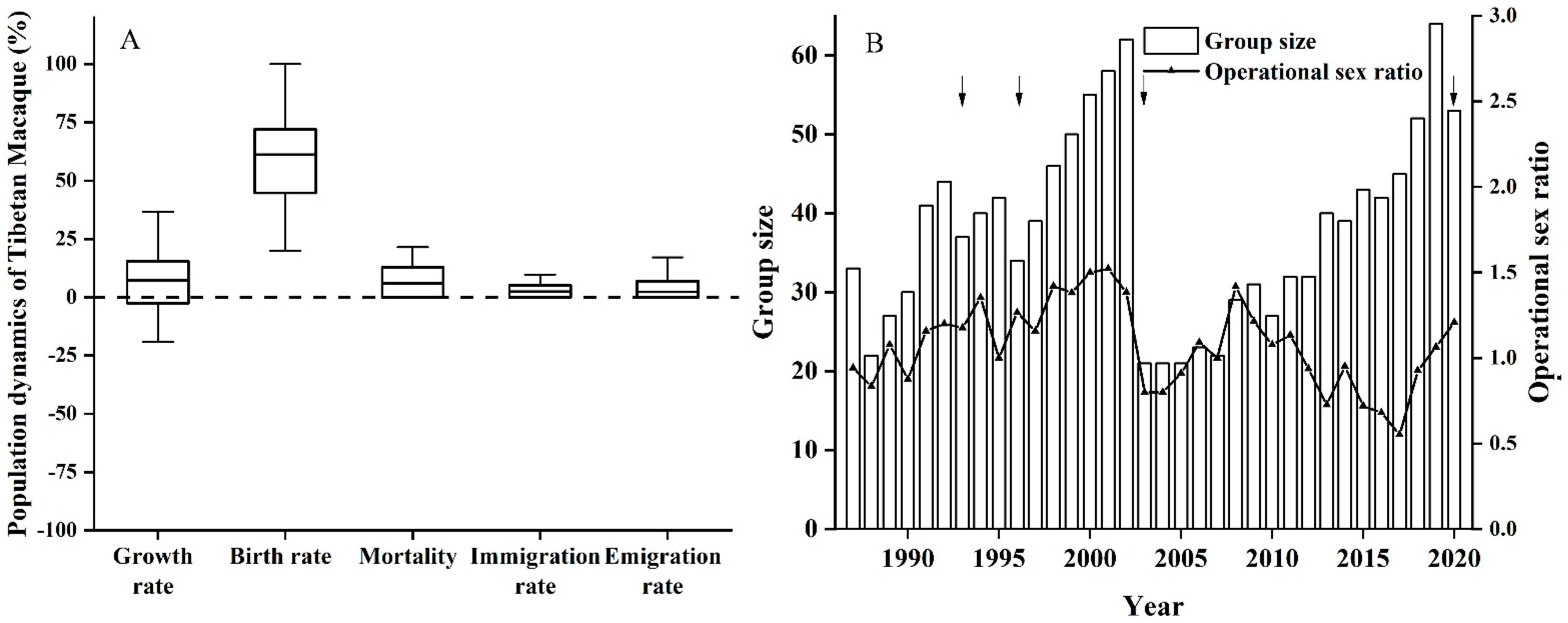
- Population dynamics
- (1)
- The annual net population growth rate is calculated as follows [48]: R = [(Nt+1/Nt − 1) × 100%.
- (2)
- Annual birth rate: b = It/Ft (Subcommittee on Conservation of Population, 1981). It is the number of births in year t, Ft is the number of adult females.
- (3)
- Annual mortality is the number of deaths within one year divided by group size at the beginning of that year.
- (4)
- The annual immigration and emigration rates were calculated as the number of immigrating and emigrating individuals divided by group size at the beginning of the year.
- (5)
- The operational sex ratio was calculated as the total number of adult males/ the total number of adult females in the group that year.
- (3)
- Evaluation of habitat suitability spatiotemporal dynamics
2.3.1. Environmental Variables
2.3.2. Human Disturbance Factors
2.3.3. Habitat Suitability Change
2.3.4. Multilevel Spatial Utilization and Habitat Suitability Change
3. Results
3.1. Population Distribution and Density
3.2. Population Dynamics of the Yulinkeng I Group
3.3. Human Population, Number of Tourists, Eco-Tourism Income
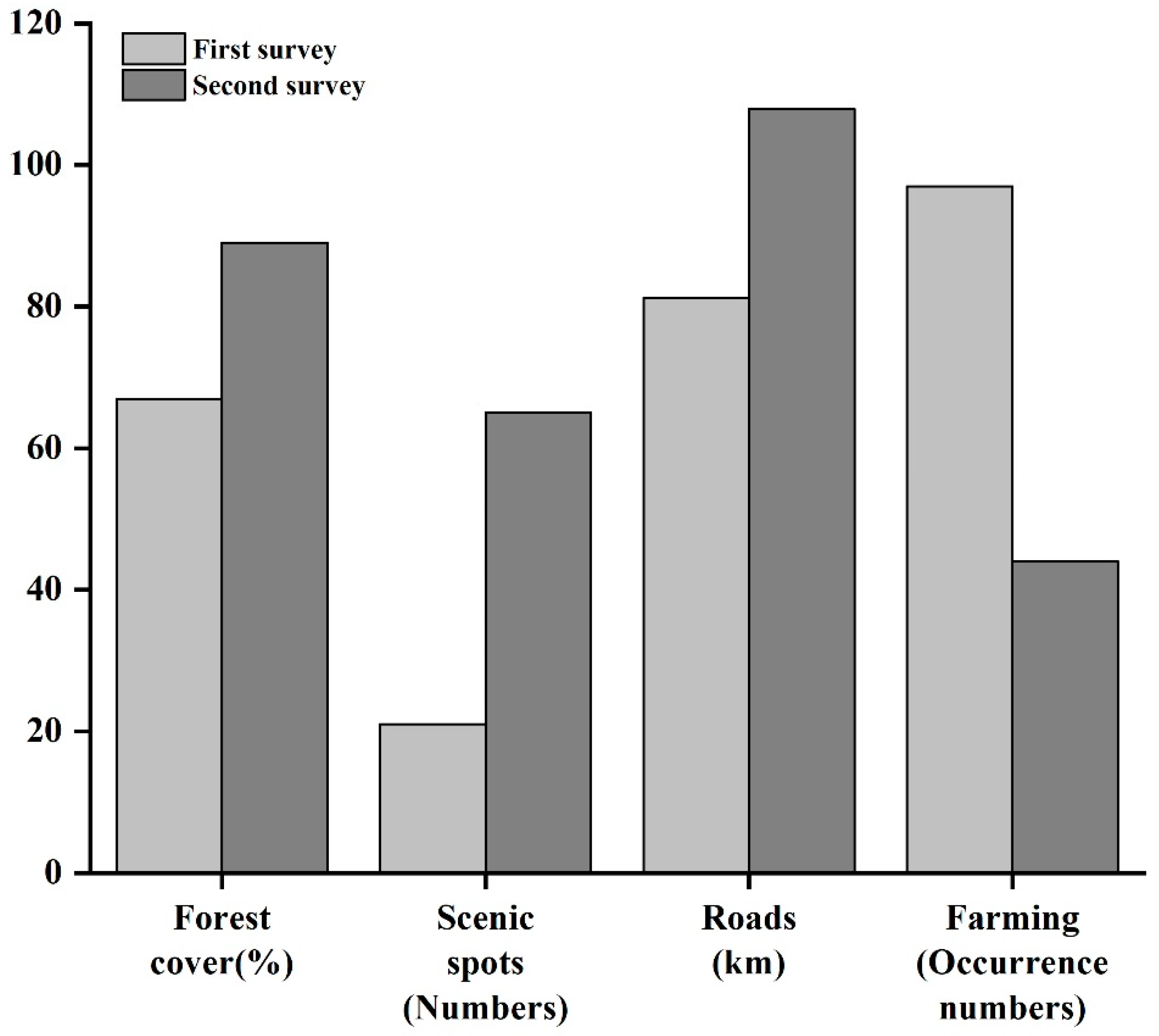
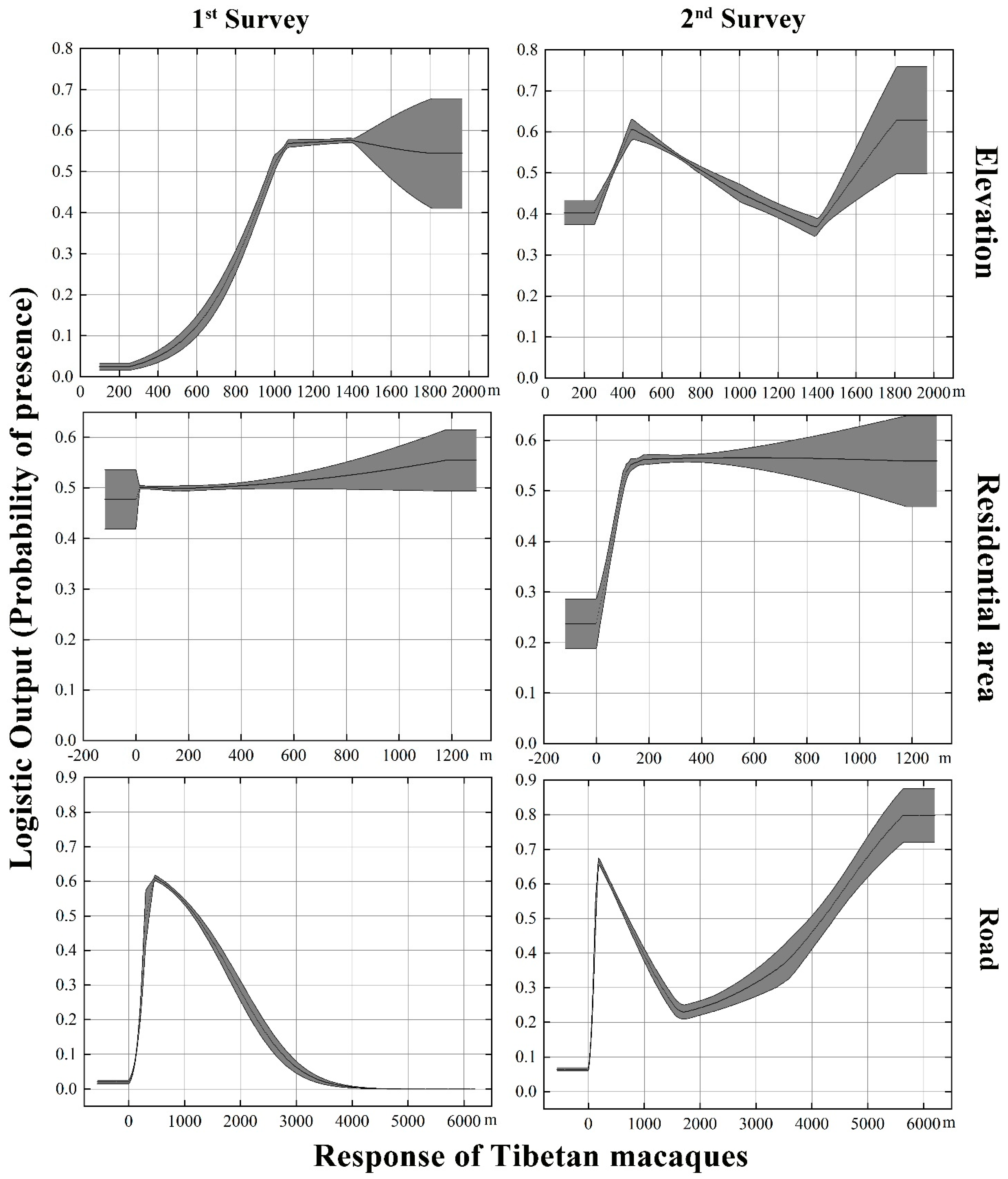
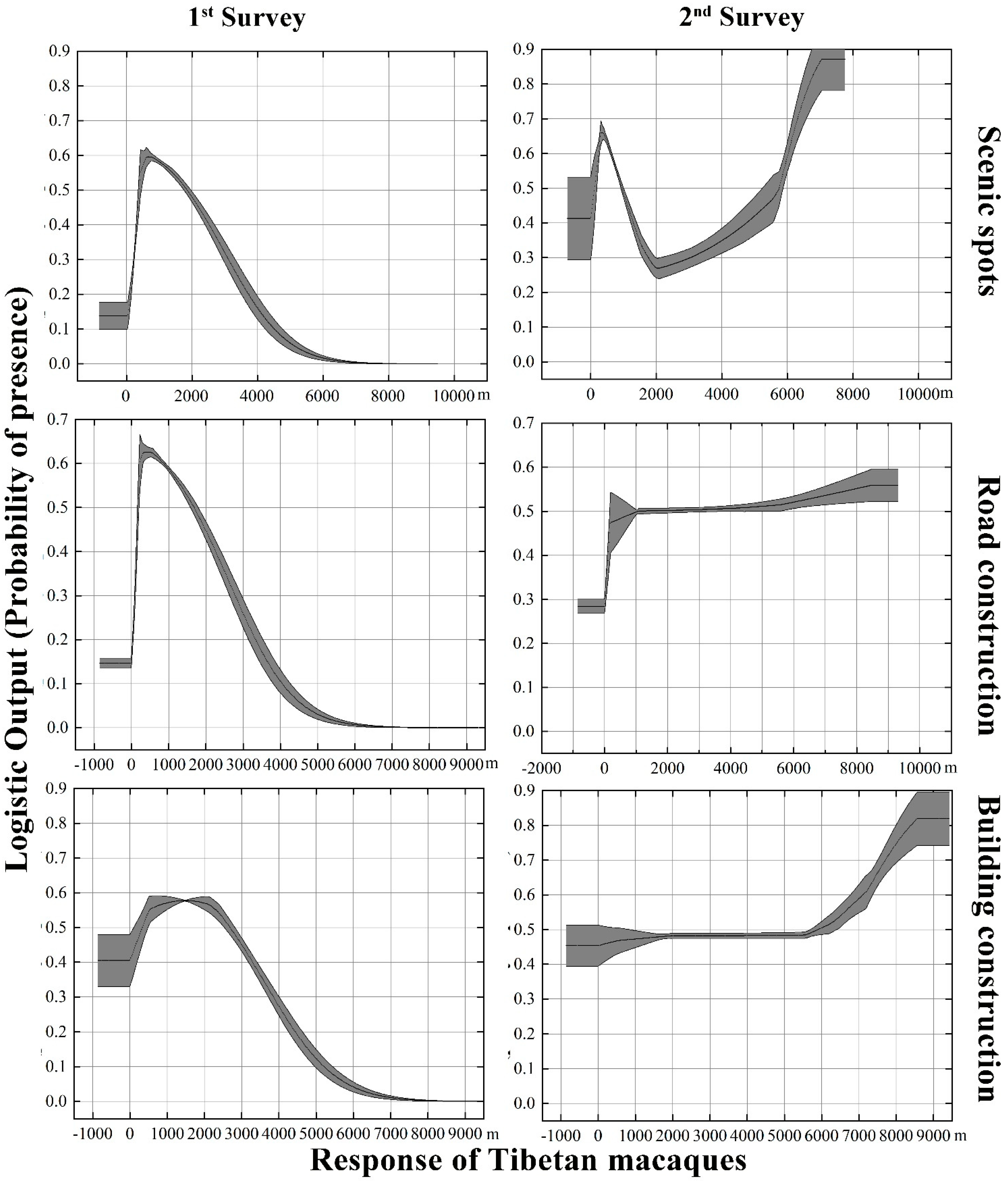
3.4. Predictor Variable Changes and Their Contributions
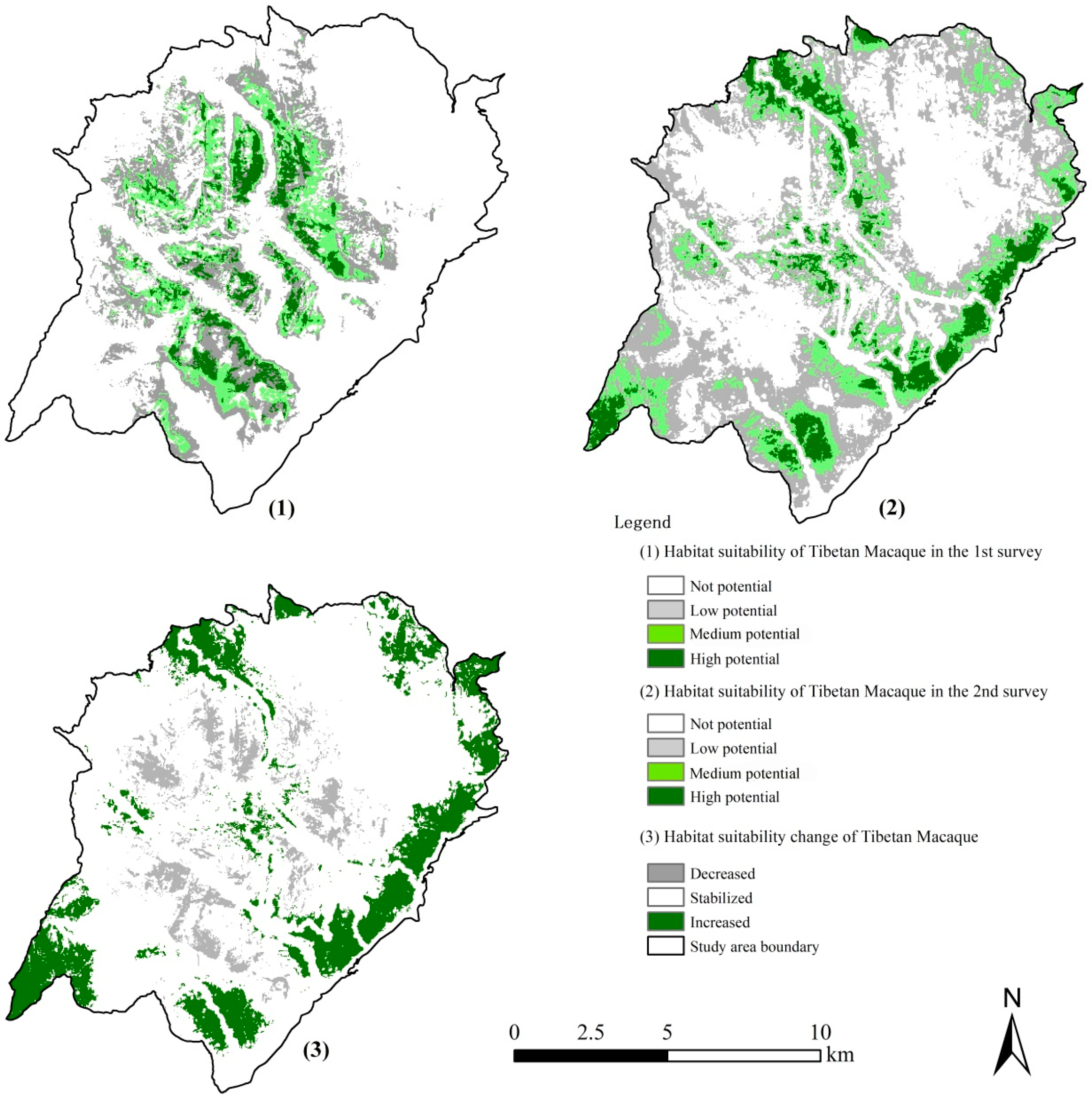
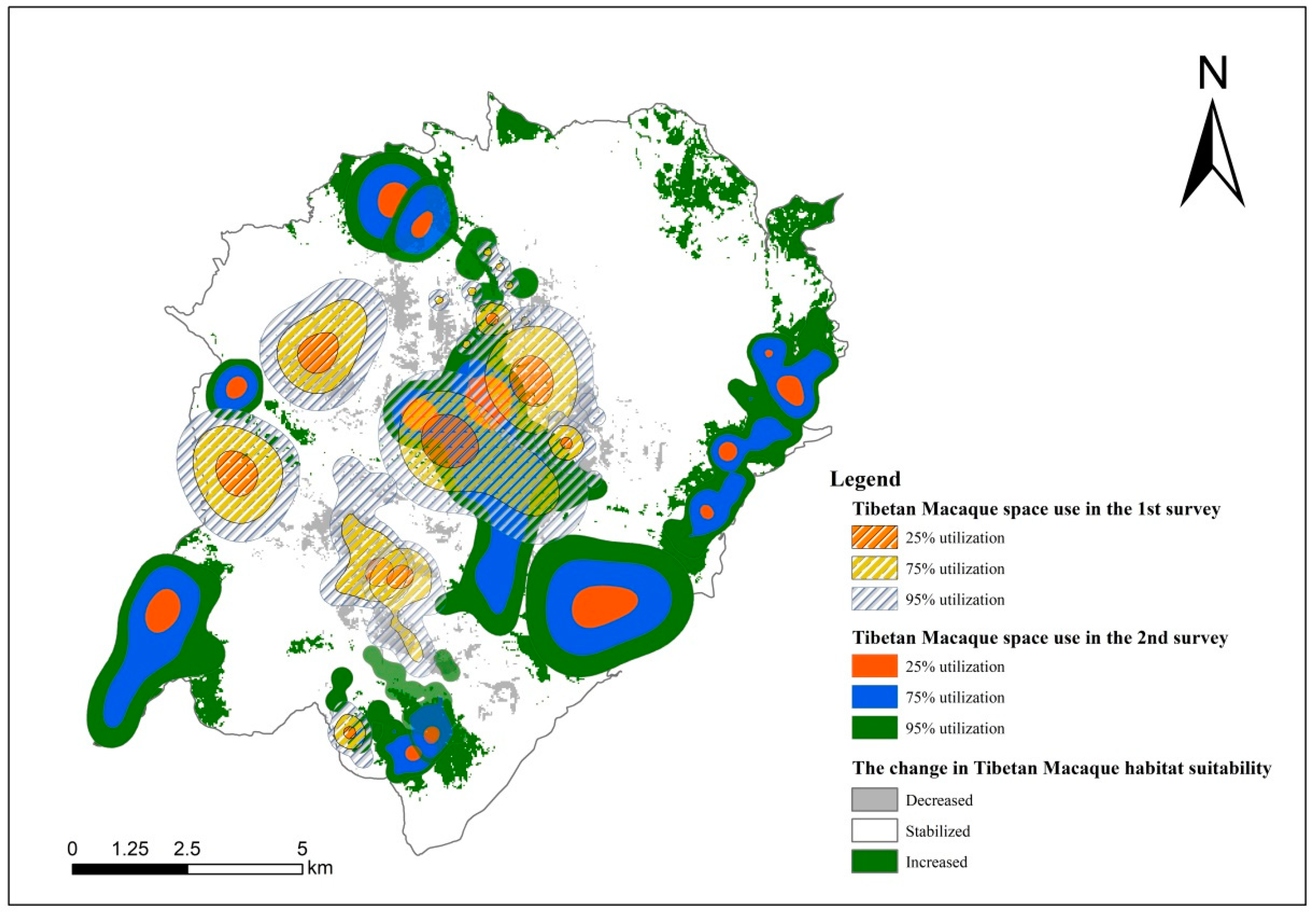
3.5. The Dynamics of Tibetan Macaque Habitat Suitability and Space Use
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reed, M.G. Conservation (In)Action: Renewing the relevance of UNESCO Biosphere Reserves. Conserv. Lett. 2016, 9, 448–456. [Google Scholar] [CrossRef]
- Franklin, J.F. The Biosphere Reserve Program in the United States. Science 1977, 195, 262–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- UNESCO MAB. Biosphere Reserves: The Seville Strategy and the Statutory Framework of the World Network of Biosphere Reserves; UNESCO: Paris, France, 1996. [Google Scholar]
- Yang, X.; Xu, M. Biodiversity conservation in Changbai Mountain Biosphere Reserve, northeastern China: Status, problem, and strategy. Biodivers. Conserv. 2003, 12, 883–903. [Google Scholar] [CrossRef]
- Ma, Z.; Li, B.; Li, W.; Han, N.; Chen, J.; Watkinson, A.R. Conflicts between biodiversity conservation and development in a biosphere reserve. J. Appl. Ecol. 2009, 46, 527–535. [Google Scholar] [CrossRef]
- Kiss, A. Is community-based ecotourism a good use of biodiversity conservation funds? Trends Ecol. Evol. 2004, 19, 232–237. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Buesching, C.D.; Newman, C.; Kaneko, Y.; Xie, Z.; Macdonald, D.W. Balancing the benefits of ecotourism and development: The effects of visitor trail-use on mammals in a Protected Area in rapidly developing China. Biol. Conserv. 2013, 165, 18–24. [Google Scholar] [CrossRef]
- Tran, L.; Walter, P. Ecotourism, gender and development in northern Vietnam. Ann. Tour. Res. 2014, 44, 116–130. [Google Scholar] [CrossRef]
- Poponi, S.; Palli, J.; Ferrari, S.; Filibeck, G.; Forte, T.a.G.W.; Franceschini, C.; Ruggieri, A.; Piovesan, G. Toward the development of sustainable ecotourism in Italian national parks of the Apennines: Insights from hiking guides. Ecol. Soc. 2020, 25, 1708–3087. [Google Scholar] [CrossRef]
- Ren, L.; Li, J.; Li, C.; Dang, P. Can ecotourism contribute to ecosystem? Evidence from local residents’ ecological behaviors. Sci. Total Environ. 2021, 757, 143814. [Google Scholar] [CrossRef]
- Hansen, A.J.; Burns, P.; Ervin, J.; Goetz, S.J.; Hansen, M.; Venter, O.; Watson, J.E.M.; Jantz, P.A.; Virnig, A.L.S.; Barnett, K.; et al. A policy-driven framework for conserving the best of Earth’s remaining moist tropical forests. Nat. Ecol. Evol. 2020, 4, 1377–1384. [Google Scholar] [CrossRef]
- Scanes, C.G. Chapter 19—Human Activity and Habitat Loss: Destruction, Fragmentation, and Degradation. In Animals and Human Society; Scanes, C.G., Toukhsati, S.R., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 451–482. [Google Scholar] [CrossRef]
- Jiang, Z.; Ma, K. The Principles of Conservation Biology; Science Press: Beijing, China, 2014. [Google Scholar]
- Li, C.; Connor, T.; Bai, W.; Yang, H.; Zhang, J.; Qi, D.; Zhou, C. Dynamics of the giant panda habitat suitability in response to changing anthropogenic disturbance in the Liangshan Mountains. Biol. Conserv. 2019, 237, 445–455. [Google Scholar] [CrossRef]
- Yang, H.; Huang, Q.; Zhang, J.; Songer, M.; Liu, J. Range-wide assessment of the impact of China’s nature reserves on giant panda habitat quality. Sci. Total Environ. 2021, 769, 145081. [Google Scholar] [CrossRef] [PubMed]
- Geijzendorffer, I.R.; Cohen-Shacham, E.; Cord, A.F.; Cramer, W.; Guerra, C.; Martín-López, B. Ecosystem services in global sustainability policies. Environ. Sci. Policy 2017, 74, 40–48. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Jorge, S.; Gomes, I.; Hayes, K.; Corti, G.; Louzao, M.; Genovart, M.; Oro, D. Effects of nature-based tourism and environmental drivers on the demography of a small dolphin population. Biol. Conserv. 2016, 197, 200–208. [Google Scholar] [CrossRef]
- Ménard, N.; Foulquier, A.; Vallet, D.; Qarro, M.; Le Gouar, P.; Pierre, J.S. How tourism and pastoralism influence population demographic changes in a threatened large mammal species. Anim. Conserv. 2014, 17, 115–124. [Google Scholar] [CrossRef]
- Kappeler, P.M.; Watts, D.P. Long-Term Field Studies of Primates; Springer: New York, NY, USA, 2012. [Google Scholar]
- Ballantyne, M.; Pickering, C.M. Tourism and recreation: A common threat to IUCN red-listed vascular plants in Europe. Biodivers. Conserv. 2013, 22, 3027–3044. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, N.; Nakamichi, M.; Sugiura, H. The Japanese Macaques; Springer: New York, NY, USA, 2010. [Google Scholar]
- Penteriani, V.; Lamamy, C.; Kojola, I.; Heikkinen, S.; Bombieri, G.; del Mar Delgado, M. Does artificial feeding affect large carnivore behaviours? The case study of brown bears in a hunted and tourist exploited subpopulation. Biol. Conserv. 2021, 254, 108949. [Google Scholar] [CrossRef]
- Steven, R.; Pickering, C.; Guy Castley, J. A review of the impacts of nature based recreation on birds. J. Environ. Manag. 2011, 92, 2287–2294. [Google Scholar] [CrossRef]
- Saltz, D.; Berger-Tal, O.; Motro, U.; Shkedy, Y.; Raanan, N. Conservation implications of habituation in Nubian ibex in response to ecotourism. Anim. Conserv. 2018, 22, 220–227. [Google Scholar] [CrossRef]
- Waterman, J.O.; Campbell, L.A.D.; Maréchal, L.; Pilot, M.; Majolo, B. Effect of human activity on habitat selection in the endangered Barbary macaque. Anim. Conserv. 2019, 23, 373–385. [Google Scholar] [CrossRef]
- Grünewald, C.; Schleuning, M.; Böhning-Gaese, K. Biodiversity, scenery and infrastructure: Factors driving wildlife tourism in an African savannah national park. Biol. Conserv. 2016, 201, 60–68. [Google Scholar] [CrossRef]
- Li, B.G.; Li, M.; Li, J.H.; Fan, P.F.; Ni, Q.Y.; Lu, J.Q.; Zhou, X.M.; Long, Y.C.; Jiang, Z.G.; Zhang, P.; et al. The primate extinction crisis in China: Immediate challenges and a way forward. Biodivers. Conserv. 2018, 27, 3301–3327. [Google Scholar] [CrossRef]
- Razafindratsima, O.H.; Sato, H.; Tsuji, Y.; Culot, L. Advances and frontiers in primate seed dispersal. Int. J. Primatol. 2018, 39, 315–320. [Google Scholar] [CrossRef] [Green Version]
- Heymann, E.W.; Culot, L.; Knogge, C.; Noriega Pina, T.E.; Tirado Herrera, E.R.; Klapproth, M.; Zinner, D. Long-term consistency in spatial patterns of primate seed dispersal. Ecol. Evol. 2017, 7, 1435–1441. [Google Scholar] [CrossRef] [Green Version]
- Wessling, E.G.; Dieguez, P.; Llana, M.; Pacheco, L.; Pruetz, J.D.; Kühl, H.S. Chimpanzee (Pan troglodytes verus) density and environmental gradients at their biogeographical range edge. Int. J. Primatol. 2020, 41, 822–848. [Google Scholar] [CrossRef]
- Lebrão, C.; Rosa, L.M.V.; Paim, F.P.; Nassar, P.M.; El Bizri, H.R.; Silva, F.E. Community-Based Ecotourism and Primate Watching as a Conservation Tool in the Amazon Rainforest. Int. J. Primatol. 2021, 42, 523–527. [Google Scholar] [CrossRef]
- Trapanese, C.; Robira, B.; Tonachella, G.; di Gristina, S.; Meunier, H.; Masi, S. Where and what? Frugivory is associated with more efficient foraging in three semi-free ranging primate species. R. Soc. Open Sci. 2019, 6, 181722. [Google Scholar] [CrossRef] [Green Version]
- Boonratana, R. Asian primates in fragments: Understanding causes and consequences of fragmentation, and predicting primate population viability. Am. J. Primatol. 2020, 82, e23082. [Google Scholar] [CrossRef]
- Condro, A.A.; Prasetyo, L.B.; Rushayati, S.B.; Santikayasa, I.P.; Iskandar, E. Predicting hotspots and prioritizing protected areas for endangered primate species in Indonesia under changing climate. Biology 2021, 10, 154. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Ren, B.; Li, D.; Garber, P.A.; Zhu, P.; Xiang, Z.; Grueter, C.C.; Liu, Z.; Li, M. Climate change, grazing, and collecting accelerate habitat contraction in an endangered primate. Biol. Conserv. 2019, 231, 88–97. [Google Scholar] [CrossRef]
- Mercado Malabet, F.; Peacock, H.; Razafitsalama, J.; Birkinshaw, C.; Colquhoun, I. Realized distribution patterns of crowned lemurs (Eulemur coronatus) within a human-dominated forest fragment in northern Madagascar. Am. J. Primatol. 2020, 82, e23125. [Google Scholar] [CrossRef] [PubMed]
- Ginath Yuh, Y.; N’Goran, P.K.; Dongmo, Z.N.; Tracz, W.; Tangwa, E.; Agunbiade, M.; Kuhl, H.S.; Sop, T.; Fotang, C. Mapping suitable great ape habitat in and around the Lobeke National Park, South-East Cameroon. Ecol. Evol. 2020, 10, 14282–14299. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Wu, F.; Tian, M.; Li, W. From Geopark to Sustainable Development: Heritage Conservation and Geotourism Promotion in the Huangshan UNESCO Global Geopark (China). Geoheritage 2017, 10, 79–91. [Google Scholar] [CrossRef]
- Li, W.B.; Yang, P.P.; Li, B.W.; Liu, C.; Sun, L.X.; Li, J.H. Habitat characteristics or protected area size: What is more important for the composition and diversity of mammals in nonprotected areas? Ecol. Evol. 2021, 11, 7250–7263. [Google Scholar] [CrossRef] [PubMed]
- Li, J.H.; Sun, L.X.; Kappeler, P.M. The Behavioral Ecology of the Tibetan Macaque; Springer: New York, NY, USA, 2020; pp. 1–303. [Google Scholar]
- IUCN. The IUCN Red List of Threatened Species. 2020. Available online: https://dx.doi.org/10.2305/IUCN.UK.2020-2.RLTS.T12562A17948236.en (accessed on 1 June 2022).
- Zhao, Q.K.; Deng, Z.Y.; Xu, J.M. Natural foods and their ecological implications for Macaca thibetana Mt. Emai, China. Folia Primatol. 1991, 57, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Xiong, C. The study of seasonal home range of Tibetan monkey’s yulinkeng troop in Huangshan mountain. Acta Theriol. Sin. 1989, 9, 239–246. [Google Scholar]
- Wada, K.; Xiong, C.; Wang, Q. On the distribution of Tibetan and Rhesus monkeys in Southern Anhui, China. Acta Theriol. Sin. 1987, 7, 168–176. [Google Scholar]
- Bezerra, B.M.; Bastos, M.; Souto, A.; Keasey, M.P.; Eason, P.; Schiel, N.; Jones, G. Camera trap observations of nonhabituated critically endangered wild blonde capuchins, Sapajus flavius (Formerly Cebus flavius). Int. J. Primatol. 2014, 35, 895–907. [Google Scholar] [CrossRef]
- He, G.; Yang, H.T.; Pan, R.L.; Sun, Y.W.; Zheng, P.B.; Wang, J.H.; Jin, X.L.; Zhang, J.J.; Li, B.G.; Guo, S.T. Using unmanned aerial vehicles with thermal-image acquisition cameras for animal surveys: A case study on the Sichuan snub-nosed monkey in the Qinling Mountains. Integr. Zool. 2020, 15, 79–86. [Google Scholar] [CrossRef]
- Wilmet, L.; Beudels-Jamar, R.C.; Schwitzer, C.; Rakotondrainibe, H.; Devillers, P.; Vermeulen, C. Use of space and home range characteristics of Lepilemur mittermeieri, an endangered sportive lemur endemic to the Ampasindava peninsula, north-west Madagascar. Am. J. Primatol. 2019, 81, e23017. [Google Scholar] [CrossRef]
- Wang, Q.; Li, J.; Li, M. Studies on population ecology of Tibetan monkeys (Macaca thibetana) I. population dynamics and analysis of Tibetan monkeys. Acta Theriol. Sin. 1994, 14, 161–165. [Google Scholar]
- Sharma, P.; Chettri, N.; Uddin, K.; Wangchuk, K.; Joshi, R.; Tandin, T.; Pandey, A.; Gaira, K.S.; Basnet, K.; Wangdi, S.; et al. Mapping human–wildlife conflict hotspots in a transboundary landscape, Eastern Himalaya. Glob. Ecol. Conserv. 2020, 24, e01284. [Google Scholar] [CrossRef]
- Cable, A.B.; O’Keefe, J.M.; Deppe, J.L.; Hohoff, T.C.; Taylor, S.J.; Davis, M.A. Habitat suitability and connectivity modeling reveal priority areas for Indiana bat (Myotis sodalis) conservation in a complex habitat mosaic. Landsc. Ecol. 2020, 36, 119–137. [Google Scholar] [CrossRef]
- Farashi, A.; Parvian, N.; Najafabadi, M.S. Land use and land cover change in protected areas: Using remote sensing to survey suitable habitats of brown bear Ursus arctos. Pol. J. Ecol. 2016, 64, 420–430. [Google Scholar] [CrossRef]
- Li, C.; Yu, J.; Wu, W.; Hou, R.; Yang, Z.; Owens, J.R.; Gu, X.; Xiang, Z.; Qi, D. Evaluating the efficacy of zoning designations for national park management. Glob. Ecol. Conserv. 2021, 27, e01562. [Google Scholar] [CrossRef]
- Zhang, K.; Yao, L.; Meng, J.; Tao, J. Maxent modeling for predicting the potential geographical distribution of two peony species under climate change. Sci. Total Environ. 2018, 634, 1326–1334. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.; Zhang, J.; Gao, H.; Cai, Z.; Zhou, X.; Li, S.; Zhang, T. Musk deer (Moschus spp.) face redistribution to higher elevations and latitudes under climate change in China. Sci. Total Environ. 2020, 704, 135335. [Google Scholar] [CrossRef] [PubMed]
- Hanya, G.; Yoshihiro, S.i.; Hayaishi, S.; Takahata, Y. Ranging patterns of Japanese macaques in the coniferous forest of Yakushima: Home range shift and travel rate. Am. J. Primatol. 2020, 82, e23185. [Google Scholar] [CrossRef]
- Ruppert, N.; Holzner, A.; See, K.W.; Gisbrecht, A.; Beck, A. Activity budgets and habitat use of wild southern pig-tailed macaques (Macaca nemestrina) in oil palm plantation and forest. Int. J. Primatol. 2018, 39, 237–251. [Google Scholar] [CrossRef]
- Silvestre, S.M.; Setchell, J.M.; Calle-Rendon, B.R.; de Toledo, J.J.; Hilario, R.R. The occurrence of the red-handed howler monkey (Alouatta belzebul) in amazonian savannas is related to forest patch area and density of flooded area palms. Am. J. Primatol. 2020, 82, e23210. [Google Scholar] [CrossRef]
- Seiler, N.; Robbins, M.M. Ecological correlates of space use patterns in wild western lowland gorillas. Am. J. Primatol. 2020, 82, e23168. [Google Scholar] [CrossRef] [PubMed]
- Schreier, A.L.; Bolt, L.M.; Russell, D.G.; Readyhough, T.S.; Jacobson, Z.S.; Merrigan-Johnson, C.; Coggeshall, E.M.C. Mantled howler monkeys (Alouatta palliata) in a Costa Rican forest fragment do not modify activity budgets or spatial cohesion in response to anthropogenic edges. Folia Primatol. 2021, 92, 49–57. [Google Scholar] [CrossRef]
- Ademola, O.J.; Vanden Broecke, B.; Leirs, H.; Mulungu, L.S.; Massawe, A.W.; Makundi, R.H. Effects of forest disturbance on the fitness of an endemic rodent in a biodiversity hotspot. Ecol. Evol. 2021, 11, 2391–2401. [Google Scholar] [CrossRef]
- Yang, L.; Shi, K.C.; Ma, C.; Ren, G.P.; Fan, P.F. Mechanisms underlying altitudinal and horizontal range contraction: The western black crested gibbon. J. Biogeogr. 2020, 48, 321–331. [Google Scholar] [CrossRef]
- Harcourt, A.H.; Schreier, B.M. Diversity, Body Mass, and Latitudinal Gradients in Primates. Int. J. Primatol. 2009, 30, 283–300. [Google Scholar] [CrossRef] [Green Version]
- Xiong, C. Ecological studies of the stump-tailed Macaque. Acta Theriol. Sin. 1984, 4, 2–9. [Google Scholar]
- Murray, M.H.; Becker, D.J.; Hall, R.J.; Hernandez, S.M. Wildlife health and supplemental feeding: A review and management recommendations. Biol. Conserv. 2016, 204, 163–174. [Google Scholar] [CrossRef]
- Shutt, K.; Heistermann, M.; Kasim, A.; Todd, A.; Kalousova, B.; Profosouva, I.; Petrzelkova, K.; Fuh, T.; Dicky, J.-F.; Bopalanzognako, J.-B.; et al. Effects of habituation, research and ecotourism on faecal glucocorticoid metabolites in wild western lowland gorillas: Implications for conservation management. Biol. Conserv. 2014, 172, 72–79. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group Name | Group Size | Range Size (km2) | Range/Altitude (m) | Data Sources |
|---|---|---|---|---|
| YAI | 52 | 2.44 | 600–1200 | Long-term study |
| YAII | 40–50 | 3.09 | 600–1300 | The long-term study, Line transects, Camera traps |
| TBSII | 40–50 | 7.07 | 500–1300 | Line transects, Camera traps |
| JLP | 40–50 | 8.81 | 400–1100 | Line transects, Camera traps |
| TLG | 30–40 | 2.06 | 700–1600 | Line transects, Camera traps |
| FRL | 40–50 | 3.63 | 500–1500 | Line transects, Camera traps |
| SG | 20–30 | 3.44 | 500–1600 | Line transects, Camera traps |
| HX | 30–40 | 14.13 | 1100–1700 | Line transects, Camera traps |
| THII | 35 | 7.13 | 300–600 | Long-term study |
| Total | 327–397 | 51.8 | - | - |
| Predictors Variables | Contribution Value | |
|---|---|---|
| First Survey (1973–1987) | Second Survey (2020–2021) | |
| Elevation | 20.60% | 2.05% |
| Slope | 1.55% | 3.30% |
| Aspect | 1.67% | 3.92% |
| Water sources | 3.25% | 8.80% |
| Forest cover | 17.14% | 3.25% |
| Farming | 0.67% | 9.97% |
| Residential area | 2.90% | 14.46% |
| Road | 19.23% | 14.80% |
| Scenic spots | 0.71% | 32.88% |
| Road construction | 18.91% | 0.59% |
| Building construction | 13.36% | 5.97% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, W.-B.; Yang, P.-P.; Xia, D.-P.; Huffman, M.A.; Li, M.; Li, J.-H. Ecotourism Disturbance on an Endemic Endangered Primate in the Huangshan Man and the Biosphere Reserve of China: A Way to Move Forward. Biology 2022, 11, 1042. https://doi.org/10.3390/biology11071042
Li W-B, Yang P-P, Xia D-P, Huffman MA, Li M, Li J-H. Ecotourism Disturbance on an Endemic Endangered Primate in the Huangshan Man and the Biosphere Reserve of China: A Way to Move Forward. Biology. 2022; 11(7):1042. https://doi.org/10.3390/biology11071042
Chicago/Turabian StyleLi, Wen-Bo, Pei-Pei Yang, Dong-Po Xia, Michael A. Huffman, Ming Li, and Jin-Hua Li. 2022. "Ecotourism Disturbance on an Endemic Endangered Primate in the Huangshan Man and the Biosphere Reserve of China: A Way to Move Forward" Biology 11, no. 7: 1042. https://doi.org/10.3390/biology11071042
APA StyleLi, W.-B., Yang, P.-P., Xia, D.-P., Huffman, M. A., Li, M., & Li, J.-H. (2022). Ecotourism Disturbance on an Endemic Endangered Primate in the Huangshan Man and the Biosphere Reserve of China: A Way to Move Forward. Biology, 11(7), 1042. https://doi.org/10.3390/biology11071042