
Impact of Metacognitive and Psychological Factors in Learning-Induced Plasticity of Resting State Networks

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
1.1. Behavioral Relevance of Resting State Networks Connectivity
1.2. RS-Networks and Learning Traces
2. Material and Methods
2.1. Sample Size and Power Considerations
2.2. Participants
2.3. Psychological Evaluations
2.4. MRI Acquisition
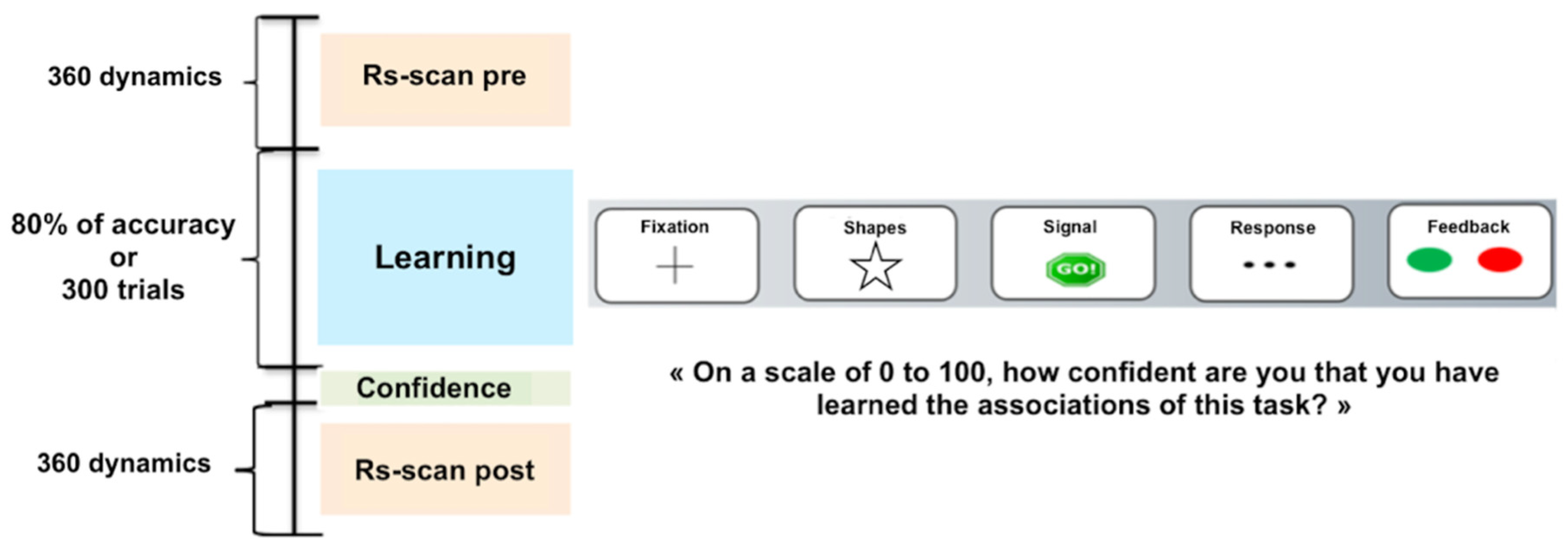
2.4.1. Learning Task
2.4.2. MRI Acquisition
2.4.3. Functional Imaging Pre-Processing
2.4.4. Whole Brain ROI to ROI Functional Connectivity
2.5. Statistical Analyses
2.5.1. Rs Functional Connectivity
2.5.2. Psychological Variables
2.5.3. Post-Hoc Analyses
3. Results
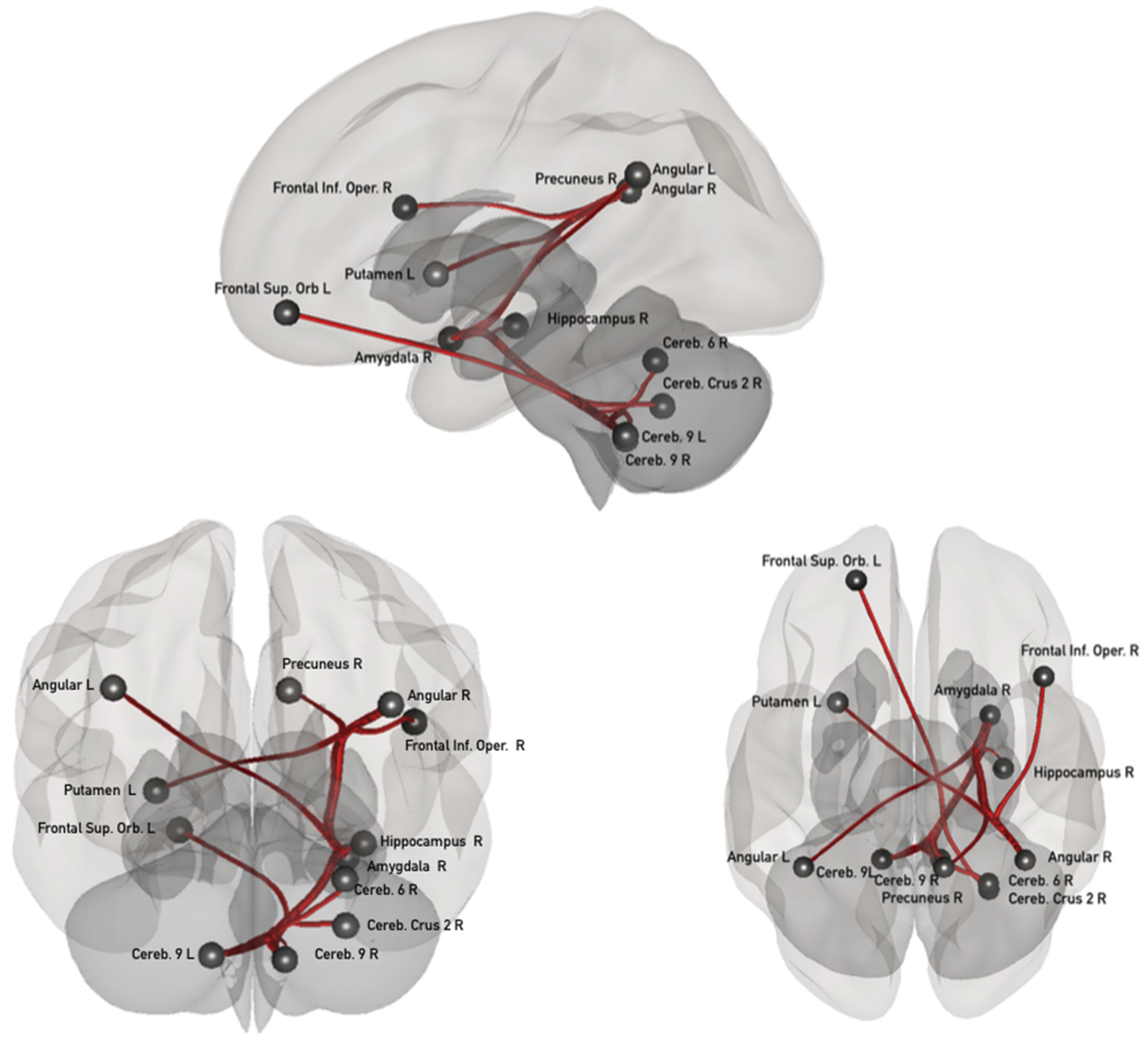
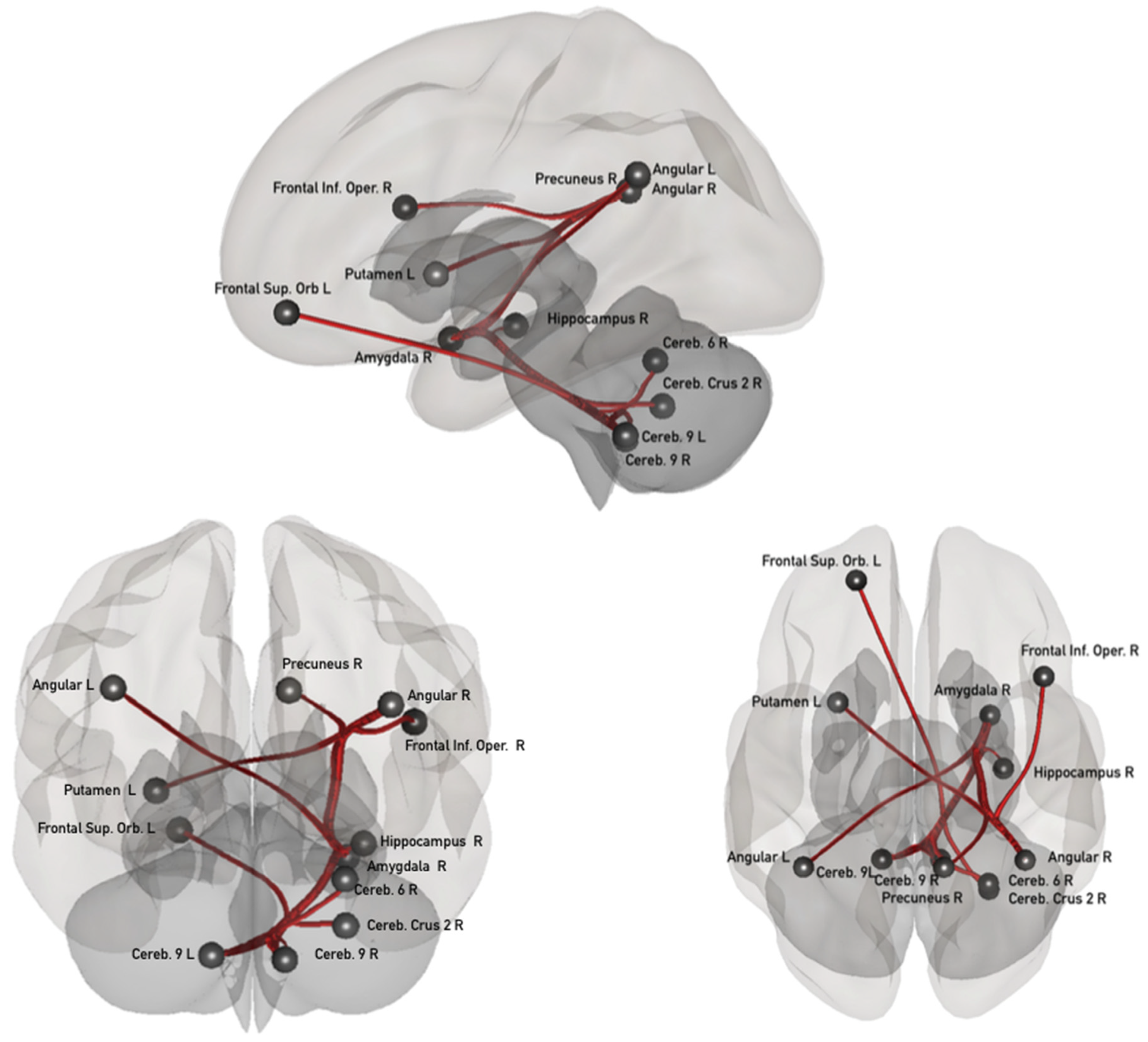
3.1. Task-Induced Resting State Network Modifications
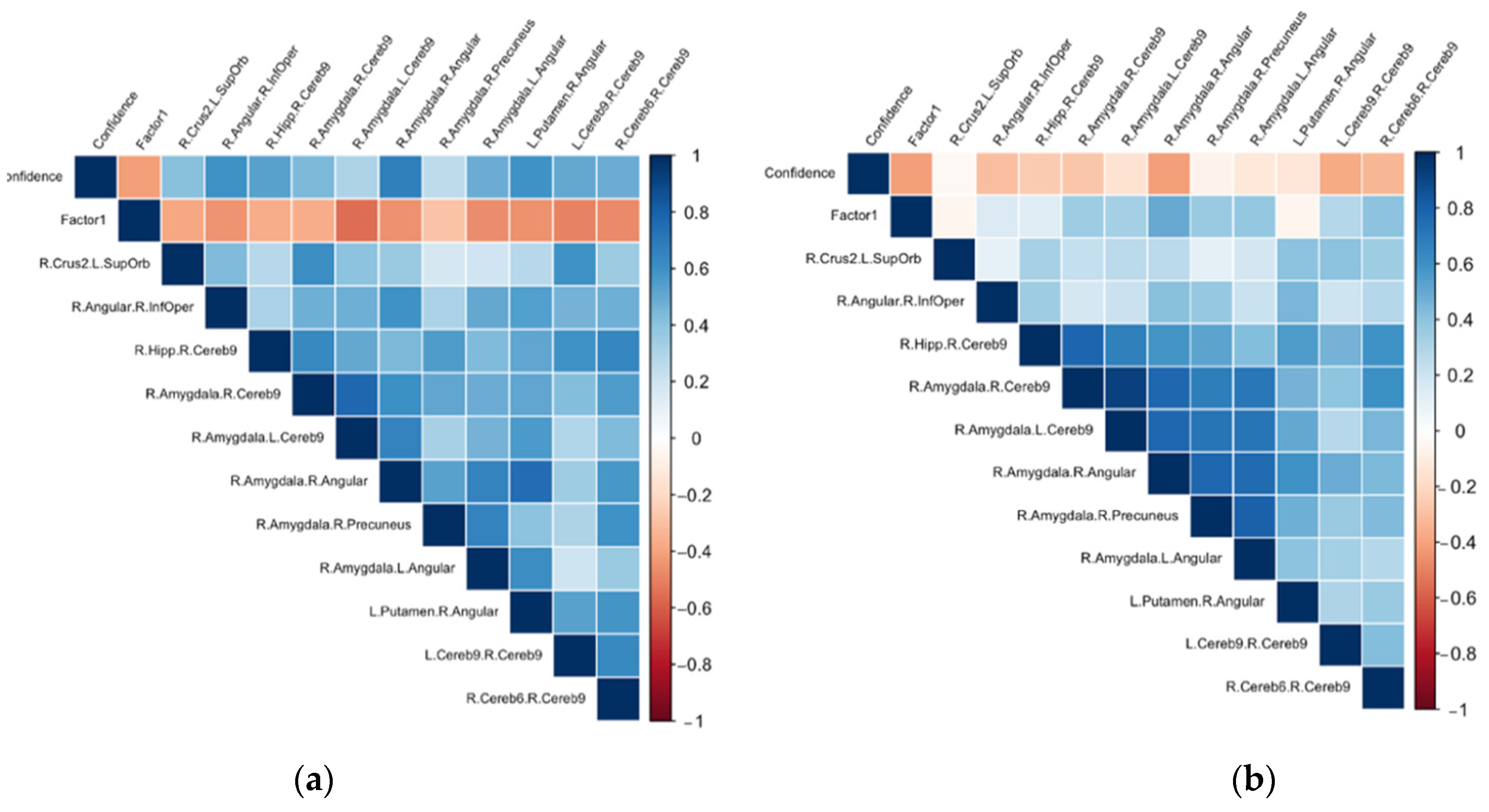
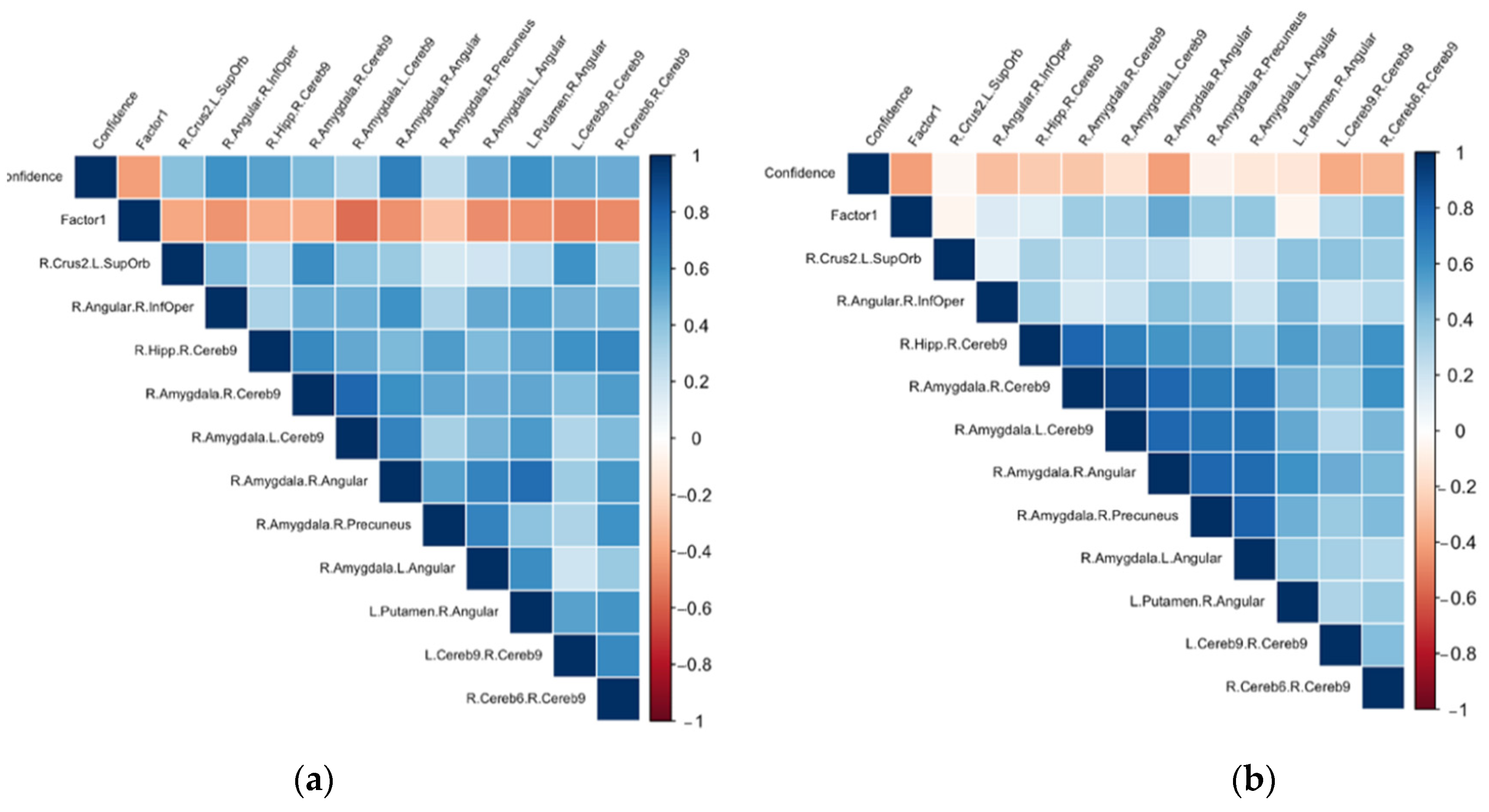
3.2. Associations between the ΔFC in the Confidence Network and the Psychological Dimensions Examined through Principal Component Analysis (PCA)
3.3. Post Hoc Analyses
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Damoiseaux, J.S.; Rombouts, S.R.B.; Barkhof, F.; Scheltens, P.; Stam, C.J.; Smith, S.M.; Beckmann, C.F. Consistent resting-state networks across healthy subjects. Proc. Natl. Acad. Sci. USA 2006, 103, 13848–13853. [Google Scholar] [CrossRef] [Green Version]
- Biswal, B.B.; Mennes, M.; Zuo, X.-N.; Gohel, S.; Kelly, C.; Smith, S.M.; Beckmann, C.F.; Adelstein, J.S.; Buckner, R.L.; Colcombe, S.; et al. Toward discovery science of human brain function. Proc. Natl. Acad. Sci. USA 2010, 107, 4734–4739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greicius, M. Resting-state functional connectivity in neuropsychiatric disorders. Curr. Opin. Neurol. 2008, 21, 424–430. [Google Scholar] [CrossRef] [PubMed]
- Hohenfeld, C.; Werner, C.J.; Reetz, K. Resting-state connectivity in neurodegenerative disorders: Is there potential for an imaging biomarker? NeuroImage Clin. 2018, 18, 849–870. [Google Scholar] [CrossRef] [PubMed]
- Cole, M.W.; Ito, T.; Bassett, D.S.; Schultz, D.H. Activity flow over resting-state networks shapes cognitive task activations. Nat. Neurosci. 2016, 19, 1718–1726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosazza, C.; Minati, L. Resting-state brain networks: Literature review and clinical applications. Neurol. Sci. 2011, 32, 773–785. [Google Scholar] [CrossRef] [PubMed]
- Edde, M.; Leroux, G.; Altena, E.; Chanraud, S. Functional brain connectivity changes across the human life span: From fetal development to old age. J. Neurosci. Res. 2021, 99, 236–262. [Google Scholar] [CrossRef]
- Kelly, C.; Castellanos, F.X. Strengthening Connections: Functional Connectivity and Brain Plasticity. Neuropsychol. Rev. 2014, 24, 63–76. [Google Scholar] [CrossRef]
- Taubert, M.; Lohmann, G.; Margulies, D.S.; Villringer, A.; Ragert, P. Long-term effects of motor training on resting-state networks and underlying brain structure. NeuroImage 2011, 57, 1492–1498. [Google Scholar] [CrossRef] [PubMed]
- Vahdat, S.; Darainy, M.; Milner, T.E.; Ostry, D.J. Functionally Specific Changes in Resting-State Sensorimotor Networks after Motor Learning. J. Neurosci. 2011, 31, 16907–16915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edde, M.; Scala, G.D.; Dupuy, M.; Dilharreguy, B.; Catheline, G.; Chanraud, S. Learning-driven cerebellar intrinsic functional connectivity changes in men. J. Neurosci. Res. 2020, 98, 668–679. [Google Scholar] [CrossRef]
- Mehrkanoon, S.; Boonstra, T.W.; Breakspear, M.; Hinder, M.; Summers, J.J. Upregulation of cortico-cerebellar functional connectivity after motor learning. NeuroImage 2016, 128, 252–263. [Google Scholar] [CrossRef] [Green Version]
- Sami, S.; Robertson, E.M.; Miall, R.C. The Time Course of Task-Specific Memory Consolidation Effects in Resting State Networks. J. Neurosci. 2014, 34, 3982–3992. [Google Scholar] [CrossRef]
- Albert, N.B.; Robertson, E.M.; Miall, R.C. The Resting Human Brain and Motor Learning. Curr. Biol. 2009, 19, 1023–1027. [Google Scholar] [CrossRef] [Green Version]
- Lewis, C.M.; Baldassarre, A.; Committeri, G.; Romani, G.L.; Corbetta, M. Learning sculpts the spontaneous activity of the resting human brain. Proc. Natl. Acad. Sci. USA 2009, 106, 17558–17563. [Google Scholar] [CrossRef] [Green Version]
- Bassett, D.S.; Wymbs, N.F.; Porter, M.A.; Mucha, P.J.; Carlson, J.M.; Grafton, S.T. Dynamic reconfiguration of human brain networks during learning. Proc. Natl. Acad. Sci. USA 2011, 108, 7641–7646. [Google Scholar] [CrossRef] [Green Version]
- Bertolero, M.A.; Adebimpe, A.; Khambhati, A.N.; Mattar, M.G.; Romer, D.; Thompson-Schill, S.L.; Bassett, D.S. Learning differentially reorganizes brain activity and connectivity. arXiv 2018, arXiv:1810.08840. [Google Scholar]
- Meyniel, F.; Schlunegger, D.; Dehaene, S. The Sense of Confidence during Probabilistic Learning: A Normative Account. PLOS Comput. Biol. 2015, 11, e1004305. [Google Scholar] [CrossRef] [Green Version]
- Fleming, S.M.; Daw, N.D. Self-Evaluation of Decision-Making: A General Bayesian Framework for Metacognitive Computation. Psychol. Rev. 2017, 124, 91–114. [Google Scholar] [CrossRef]
- Meyniel, F.; Dehaene, S. Brain networks for confidence weighting and hierarchical inference during probabilistic learning. Proc. Natl. Acad. Sci. USA 2017, 114, E3859–E3868. [Google Scholar] [CrossRef] [Green Version]
- Guggenmos, M.; Sterzer, P. A confidence-based reinforcement learning model for perceptual learning. bioRxiv 2017, 136903. [Google Scholar] [CrossRef] [Green Version]
- Song, C.; Kanai, R.; Fleming, S.M.; Weil, R.S.; Schwarzkopf, D.S.; Rees, G. Relating inter-individual differences in metacognitive performance on different perceptual tasks. Conscious. Cogn. 2011, 20, 1787–1792. [Google Scholar] [CrossRef] [Green Version]
- Kanai, R.; Rees, G. The structural basis of inter-individual differences in human behaviour and cognition. Nat. Rev. Neurosci. 2011, 12, 231–242. [Google Scholar] [CrossRef]
- Toro, R. On the Possible Shapes of the Brain. Evol. Biol. 2012, 39, 600–612. [Google Scholar] [CrossRef] [Green Version]
- Spada, M.M.; Georgiou, G.A.; Wells, A. The Relationship among Metacognitions, Attentional Control, and State Anxiety. Cogn. Behav. Ther. 2010, 39, 64–71. [Google Scholar] [CrossRef]
- Modi, S.; Kumar, M.; Kumar, P.; Khushu, S. Aberrant functional connectivity of resting state networks associated with trait anxiety. Psychiatry Res. Neuroimaging 2015, 234, 25–34. [Google Scholar] [CrossRef]
- Saviola, F.; Pappaianni, E.; Monti, A.; Grecucci, A.; Jovicich, J.; De Pisapia, N. Trait and state anxiety are mapped differently in the human brain. Sci. Rep. 2020, 10, 11112. [Google Scholar] [CrossRef]
- Balsters, J.H.; Whelan, C.D.; Robertson, I.H.; Ramnani, N. Cerebellum and Cognition: Evidence for the Encoding of Higher Order Rules. Cereb. Cortex 2013, 23, 1433–1443. [Google Scholar] [CrossRef] [Green Version]
- Whitfield-Gabrieli, S.; Nieto-Castanon, A. Conn: A Functional Connectivity Toolbox for Correlated and Anticorrelated Brain Networks. Brain Connect. 2012, 2, 125–141. [Google Scholar] [CrossRef] [Green Version]
- Behzadi, Y.; Restom, K.; Liau, J.; Liu, T.T. A component based noise correction method (CompCor) for BOLD and perfusion based fMRI. NeuroImage 2007, 37, 90–101. [Google Scholar] [CrossRef] [Green Version]
- Rolls, E.T.; Joliot, M.; Tzourio-Mazoyer, N. Implementation of a new parcellation of the orbitofrontal cortex in the automated anatomical labeling atlas. Neuroimage 2015, 122, 1–5. [Google Scholar] [CrossRef]
- Tzourio-Mazoyer, N.; Landeau, B.; Papathanassiou, D.; Crivello, F.; Etard, O.; Delcroix, N.; Mazoyer, B.; Joliot, M. Automated anatomical labeling of activations in SPM using a macroscopic anatomical parcellation of the MNI MRI single-subject brain. Neuroimage 2002, 15, 273–289. [Google Scholar] [CrossRef]
- Kelly, A.M.C.; Garavan, H. Human functional neuroimaging of brain changes associated with practice. Cereb Cortex 2005, 15, 1089–1102. [Google Scholar] [CrossRef] [Green Version]
- Molchan, S.E.; Sunderland, T.; McIntosh, A.R.; Herscovitch, P.; Schreurs, B.G. A functional anatomical study of associative learning in humans. Proc. Natl. Acad. Sci. USA 1994, 91, 8122–8126. [Google Scholar] [CrossRef] [Green Version]
- Sehlmeyer, C.; Schöning, S.; Zwitserlood, P.; Pfleiderer, B.; Kircher, T.; Arolt, V.; Konrad, C. Human Fear Conditioning and Extinction in Neuroimaging: A Systematic Review. PLoS ONE 2009, 4, e5865. [Google Scholar] [CrossRef]
- Ekhtiari, H.; Nasseri, P.; Yavari, F.; Mokri, A.; Monterosso, J. Neuroscience of drug craving for addiction medicine. In Progress in Brain Research; Elsevier: Amsterdam, The Netherlands, 2016; Volume 223, pp. 115–141. ISBN 978-0-444-63545-7. [Google Scholar]
- Ren, Y.; Nguyen, V.T.; Sonkusare, S.; Lv, J.; Pang, T.; Guo, L.; Eickhoff, S.B.; Breakspear, M.; Guo, C.C. Effective connectivity of the anterior hippocampus predicts recollection confidence during natural memory retrieval. Nat. Commun. 2018, 9, 4875. [Google Scholar] [CrossRef] [Green Version]
- Luttrell, A.; Briñol, P.; Petty, R.E.; Cunningham, W.; Díaz, D. Metacognitive confidence: A neuroscience approach. Rev. Psicol. Soc. 2013, 28, 317–332. [Google Scholar] [CrossRef] [Green Version]
- Seghier, M.L. The angular gyrus: Multiple functions and multiple subdivisions. Neuroscientist 2013, 19, 43–61. [Google Scholar] [CrossRef] [Green Version]
- Sreekumar, V.; Nielson, D.M.; Smith, T.A.; Dennis, S.J.; Sederberg, P.B. The experience of vivid autobiographical reminiscence is supported by subjective content representations in the precuneus. Sci. Rep. 2018, 8, 14899. [Google Scholar] [CrossRef]
- Daniel, R.; Pollmann, S. Striatal activations signal prediction errors on confidence in the absence of external feedback. Neuroimage 2012, 59, 3457–3467. [Google Scholar] [CrossRef]
- Hsu, M.; Bhatt, M.; Adolphs, R.; Tranel, D.; Camerer, C.F. Neural Systems Responding to Degrees of Uncertainty in Human Decision-Making. Science 2005, 310, 1680–1683. [Google Scholar] [CrossRef] [Green Version]
- Sommer, S.; Pollmann, S. Putamen Activation Represents an Intrinsic Positive Prediction Error Signal for Visual Search in Repeated Configurations. Open Neuroimag J. 2016, 10, 126–138. [Google Scholar] [CrossRef] [Green Version]
- Seymour, B.; Dolan, R. Emotion, Decision Making, and the Amygdala. Neuron 2008, 58, 662–671. [Google Scholar] [CrossRef] [Green Version]
- Ito, M. Bases and implications of learning in the cerebellum-adaptive control and internal model mechanism. In Progress in Brain Research; Elsevier: Amsterdam, The Netherlands, 2005; Volume 148, pp. 95–109, Creating coordination in the cerebellum. [Google Scholar]
- D’Angelo, E.; Casali, S. Seeking a unified framework for cerebellar function and dysfunction: From circuit operations to cognition. Front. Neural Circuits 2013, 6, 116. [Google Scholar] [CrossRef] [Green Version]
- Habas, C.; Kamdar, N.; Nguyen, D.; Prater, K.; Beckmann, C.F.; Menon, V.; Greicius, M.D. Distinct Cerebellar Contributions to Intrinsic Connectivity Networks. J. Neurosci. 2009, 29, 8586–8594. [Google Scholar] [CrossRef]
- Conant, R.C.; Ross Ashby, W. Every good regulator of a system must be a model of that system †. Int. J. Syst. Sci. 1970, 1, 89–97. [Google Scholar] [CrossRef]
- Nelson, T.O. Metamemory: A Theoretical Framework and New Findings. In Psychology of Learning and Motivation; Elsevier: Amsterdam, The Netherlands, 1990; Volume 26, pp. 125–173. ISBN 978-0-12-543326-6. [Google Scholar]
- Kentridge, R.W.; Heywood, C.A. Metacognition and Awareness. Conscious. Cogn. 2000, 9, 308–312. [Google Scholar] [CrossRef] [Green Version]
- Stoodley, C.J. The Cerebellum and Cognition: Evidence from Functional Imaging Studies. Cerebellum 2012, 11, 352–365. [Google Scholar] [CrossRef]
- DeBerry, L. The Relation Between Cognitive Inflexibility and Obsessive-Compulsive Personality Traits in Adults: Depression and Anxiety as Potential Mediators 2012. Master’s Thesis, University of Southern Mississippi, Hattiesburg, MS, USA, 2012. [Google Scholar]
- Wilson, C.G.; Nusbaum, A.T.; Whitney, P.; Hinson, J.M. Trait anxiety impairs cognitive flexibility when overcoming a task acquired response and a preexisting bias. PLoS ONE 2018, 13, e0204694. [Google Scholar] [CrossRef]
- Williams, L.M. Defining biotypes for depression and anxiety based on large-scale circuit dysfunction: A theoretical review of the evidence and future directions for clinical translation. Depress. Anxiety 2017, 34, 9–24. [Google Scholar] [CrossRef]
- Arnold Anteraper, S.; Triantafyllou, C.; Sawyer, A.T.; Hofmann, S.G.; Gabrieli, J.D.; Whitfield-Gabrieli, S. Hyper-Connectivity of Subcortical Resting-State Networks in Social Anxiety Disorder. Brain Connect. 2013, 4, 81–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Yin, D.; Cheng, W.; Fan, M.; You, M.; Men, W.; Zang, L.; Shi, D.; Zhang, F. Abnormal Functional Connectivity of the Amygdala-Based Network in Resting-State fMRI in Adolescents with Generalized Anxiety Disorder. Med. Sci. Monit. 2015, 21, 459–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.J.; Guell, X.; Hubbard, N.A.; Siless, V.; Frosch, I.R.; Goncalves, M.; Lo, N.; Nair, A.; Ghosh, S.S.; Hofmann, S.G.; et al. Functional Alterations in Cerebellar Functional Connectivity in Anxiety Disorders. Cerebellum 2020, 20, 392–401. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Yin, S.; Zhu, X.; Ren, W.; Yu, J.; Wang, P.; Zheng, Z.; Niu, Y.-N.; Huang, X.; Li, J. Linking Inter-Individual Variability in Functional Brain Connectivity to Cognitive Ability in Elderly Individuals. Front. Aging Neurosci. 2017, 9, 385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koziol, L.F.; Budding, D.; Andreasen, N.; D’Arrigo, S.; Bulgheroni, S.; Imamizu, H.; Ito, M.; Manto, M.; Marvel, C.; Parker, K.; et al. Consensus Paper: The Cerebellum’s Role in Movement and Cognition. Cerebellum 2014, 13, 151–177. [Google Scholar] [CrossRef] [PubMed]
- Adamaszek, M.; D’Agata, F.; Ferrucci, R.; Habas, C.; Keulen, S.; Kirkby, K.C.; Leggio, M.; Mariën, P.; Molinari, M.; Moulton, E.; et al. Consensus Paper: Cerebellum and Emotion. Cerebellum 2017, 16, 552–576. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| N | Mean | Sd | Min | Max | |
| Learning accuracy (%) | 30 | 71.80 | 14.10 | 31.70 | 81.10 |
| Self-confidence (%) | 30 | 69.30 | 26.20 | 25 | 100 |
| FMPS: concerns over mistake | 26 | 21.20 | 7.66 | 12 | 43 |
| FMPS: doubt about actions | 26 | 11.00 | 3.34 | 5 | 18 |
| FMPS: personal standards | 26 | 22.00 | 4.82 | 11 | 34 |
| Sensitivity to punishment | 26 | 42.20 | 8.82 | 26 | 60 |
| Sensitivity to rewards | 26 | 36.10 | 6.75 | 21 | 52 |
| State anxiety | 26 | 33.60 | 9.85 | 20 | 55 |
| Trait anxiety | 26 | 40.90 | 12.30 | 21 | 67 |
| Depression | 26 | 5.00 | 3.62 | 0 | 14 |
| Anxiety | 26 | 6.38 | 3.72 | 2 | 17 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chirokoff, V.; Di Scala, G.; Swendsen, J.; Dilharreguy, B.; Berthoz, S.; Chanraud, S. Impact of Metacognitive and Psychological Factors in Learning-Induced Plasticity of Resting State Networks. Biology 2022, 11, 896. https://doi.org/10.3390/biology11060896
Chirokoff V, Di Scala G, Swendsen J, Dilharreguy B, Berthoz S, Chanraud S. Impact of Metacognitive and Psychological Factors in Learning-Induced Plasticity of Resting State Networks. Biology. 2022; 11(6):896. https://doi.org/10.3390/biology11060896
Chicago/Turabian StyleChirokoff, Valentine, Georges Di Scala, Joel Swendsen, Bixente Dilharreguy, Sylvie Berthoz, and Sandra Chanraud. 2022. "Impact of Metacognitive and Psychological Factors in Learning-Induced Plasticity of Resting State Networks" Biology 11, no. 6: 896. https://doi.org/10.3390/biology11060896
APA StyleChirokoff, V., Di Scala, G., Swendsen, J., Dilharreguy, B., Berthoz, S., & Chanraud, S. (2022). Impact of Metacognitive and Psychological Factors in Learning-Induced Plasticity of Resting State Networks. Biology, 11(6), 896. https://doi.org/10.3390/biology11060896