
Reproductive Biology of the Sea Cucumber Holothuria mammata (Echinodermata: Holothuroidea)

 ,
,  ,
,  , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling Strategy
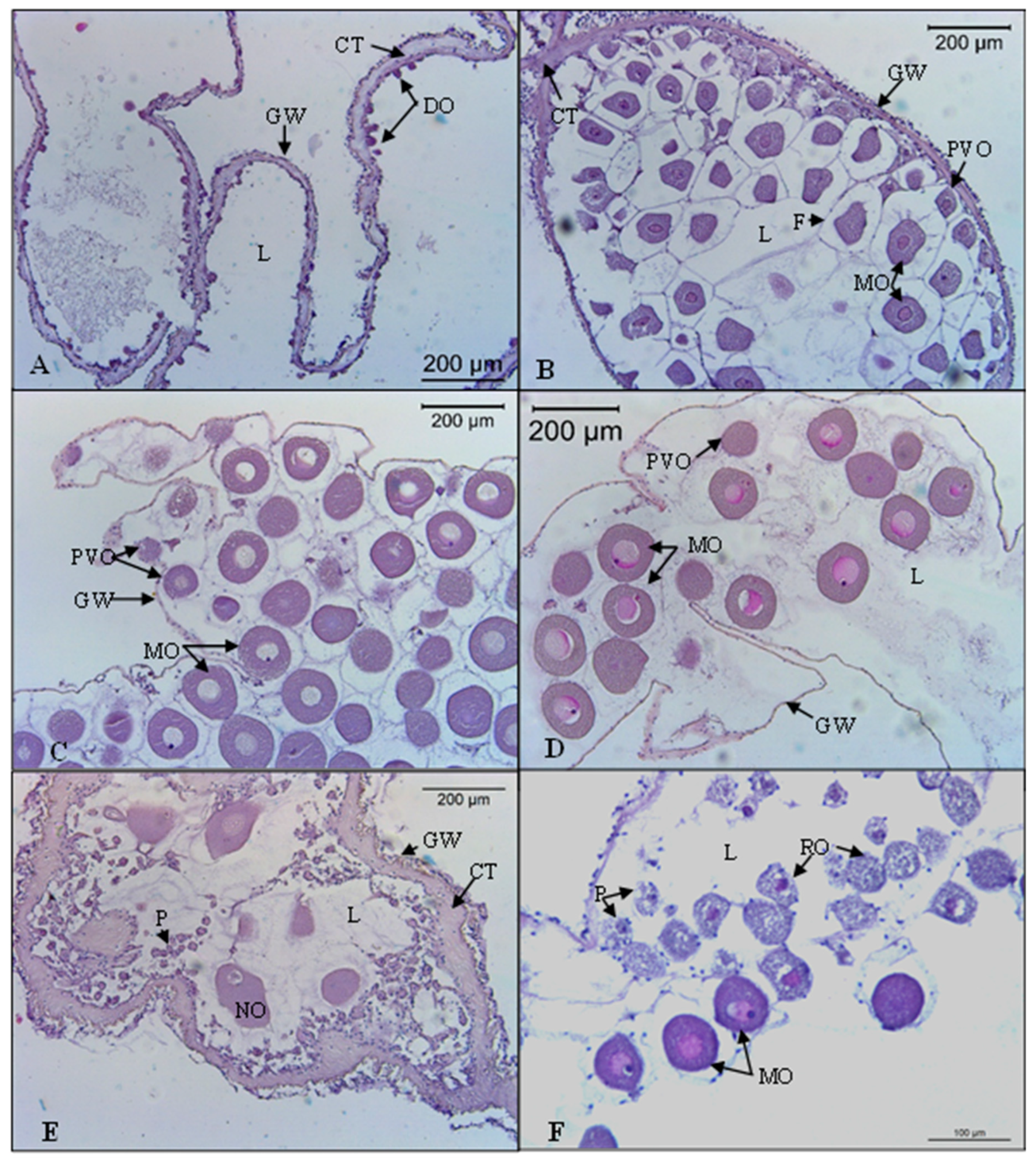
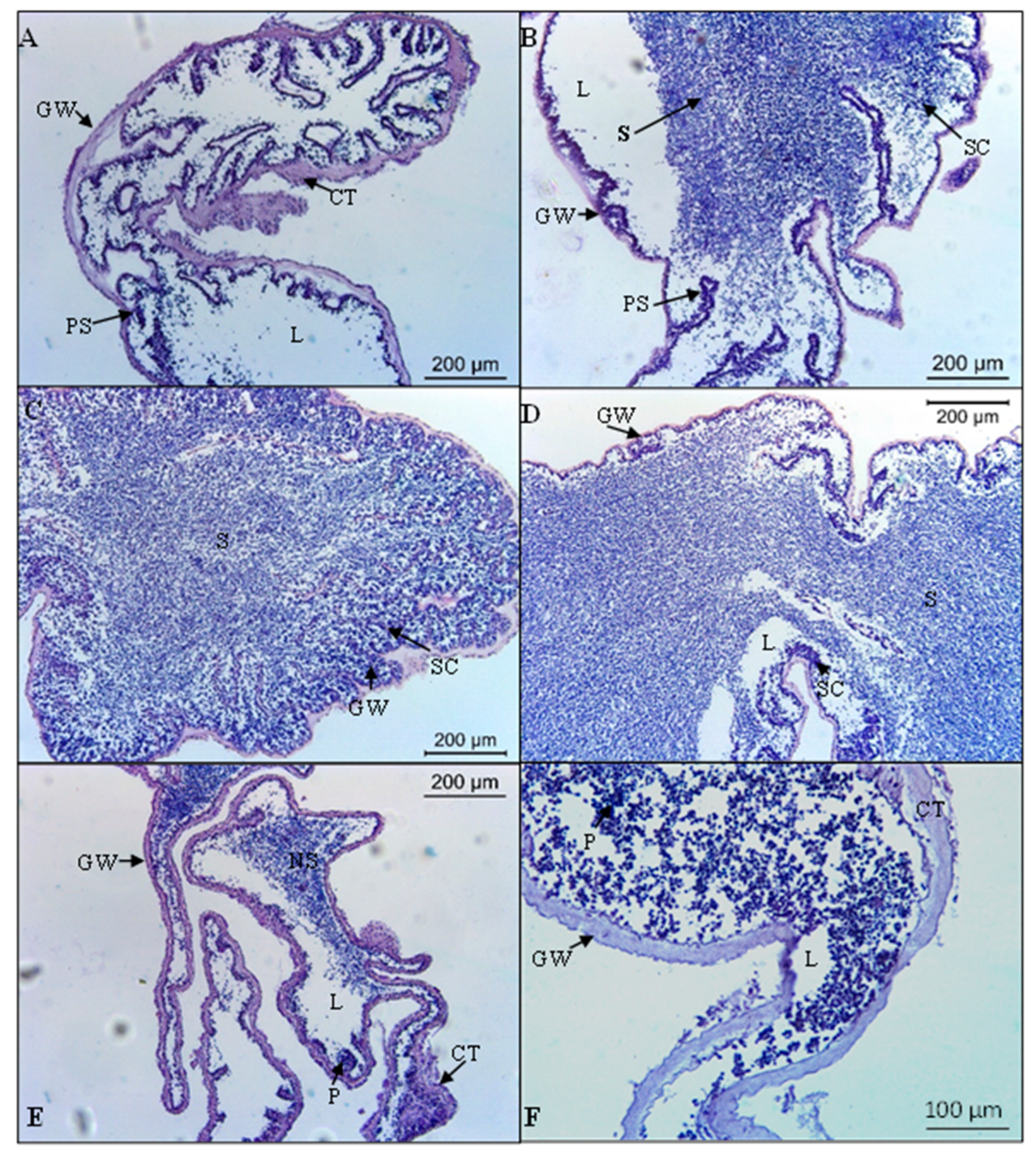
2.2. Histological Analysis
2.3. Data Analysis
3. Results
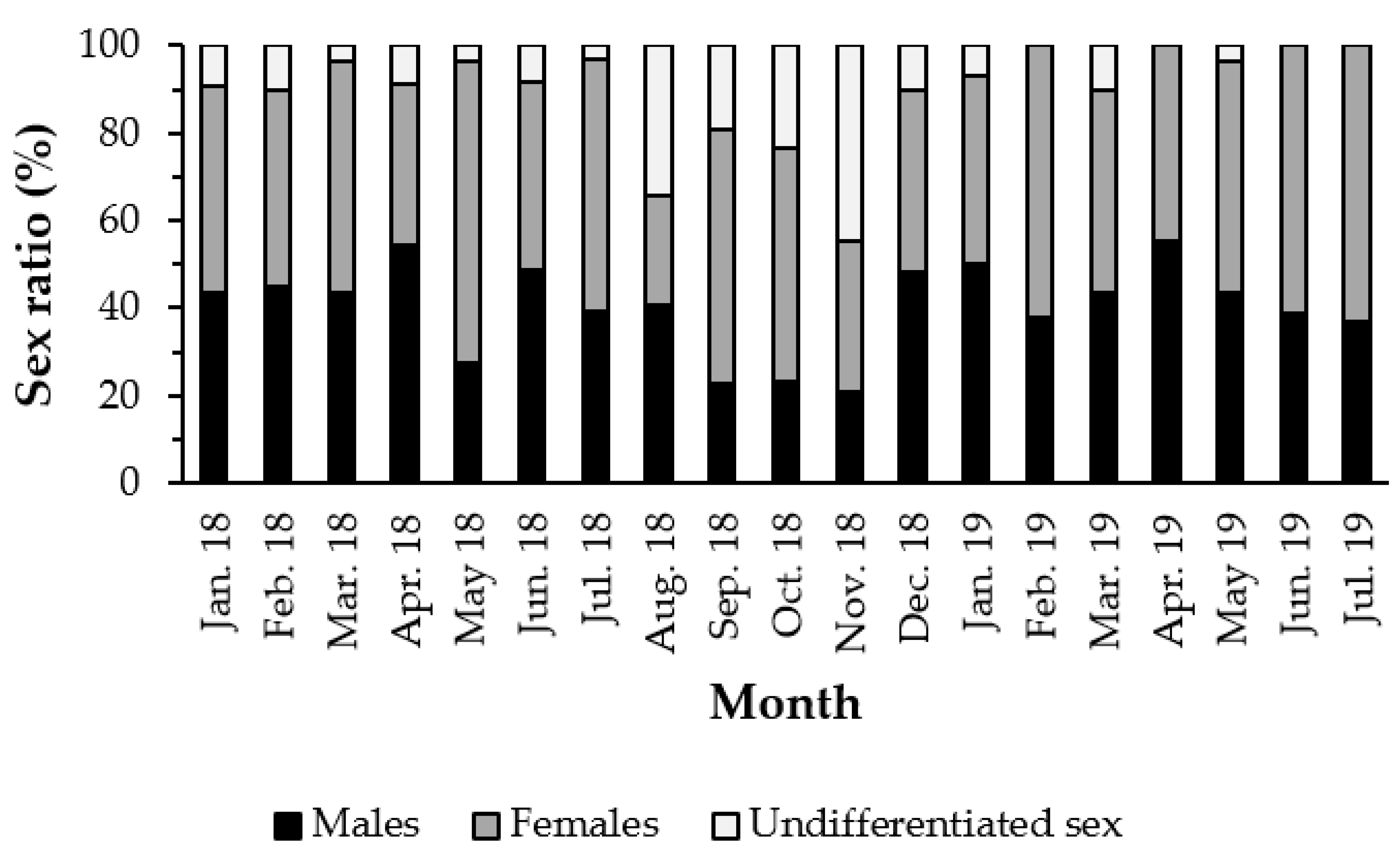
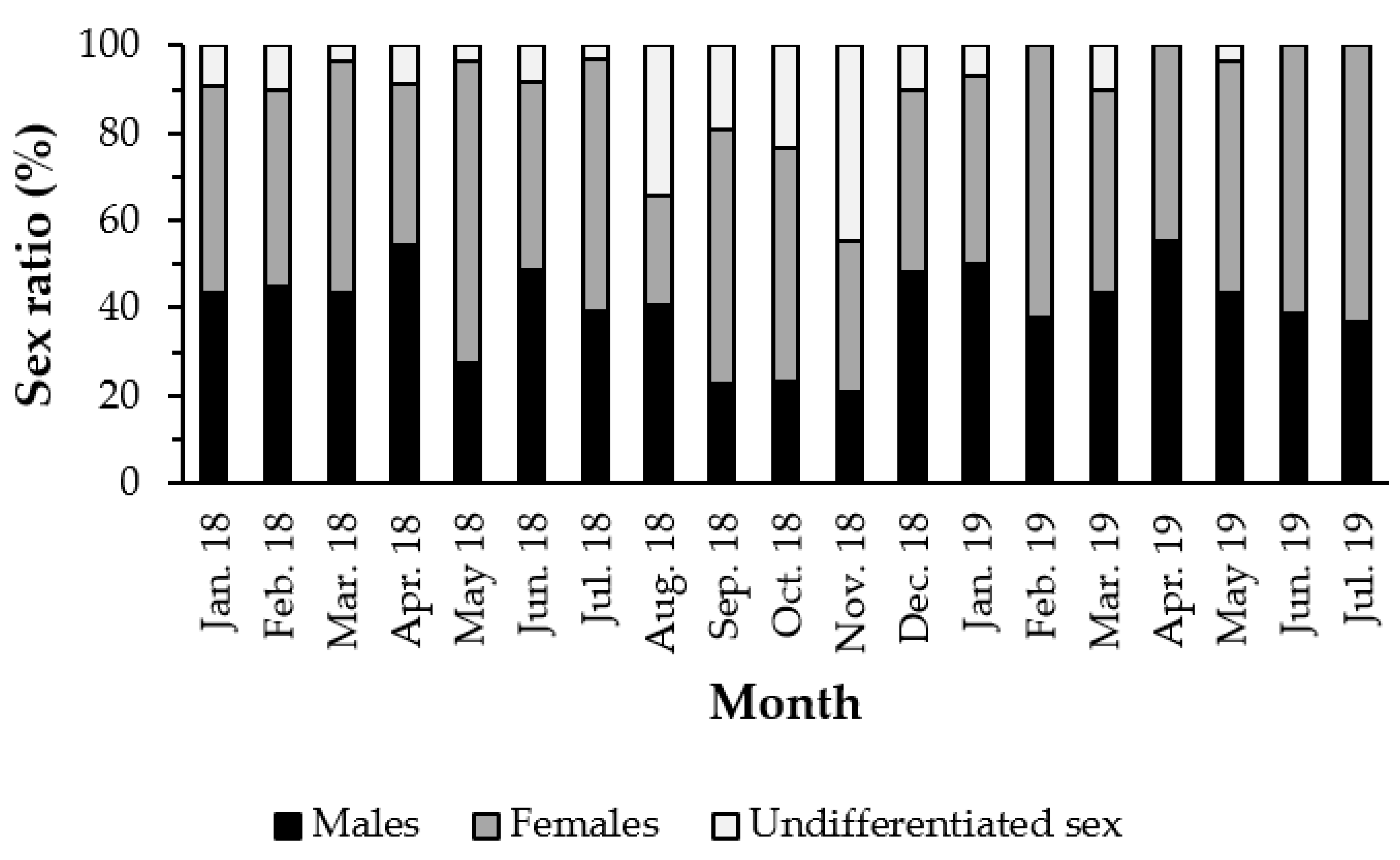
3.1. Sex Ratio
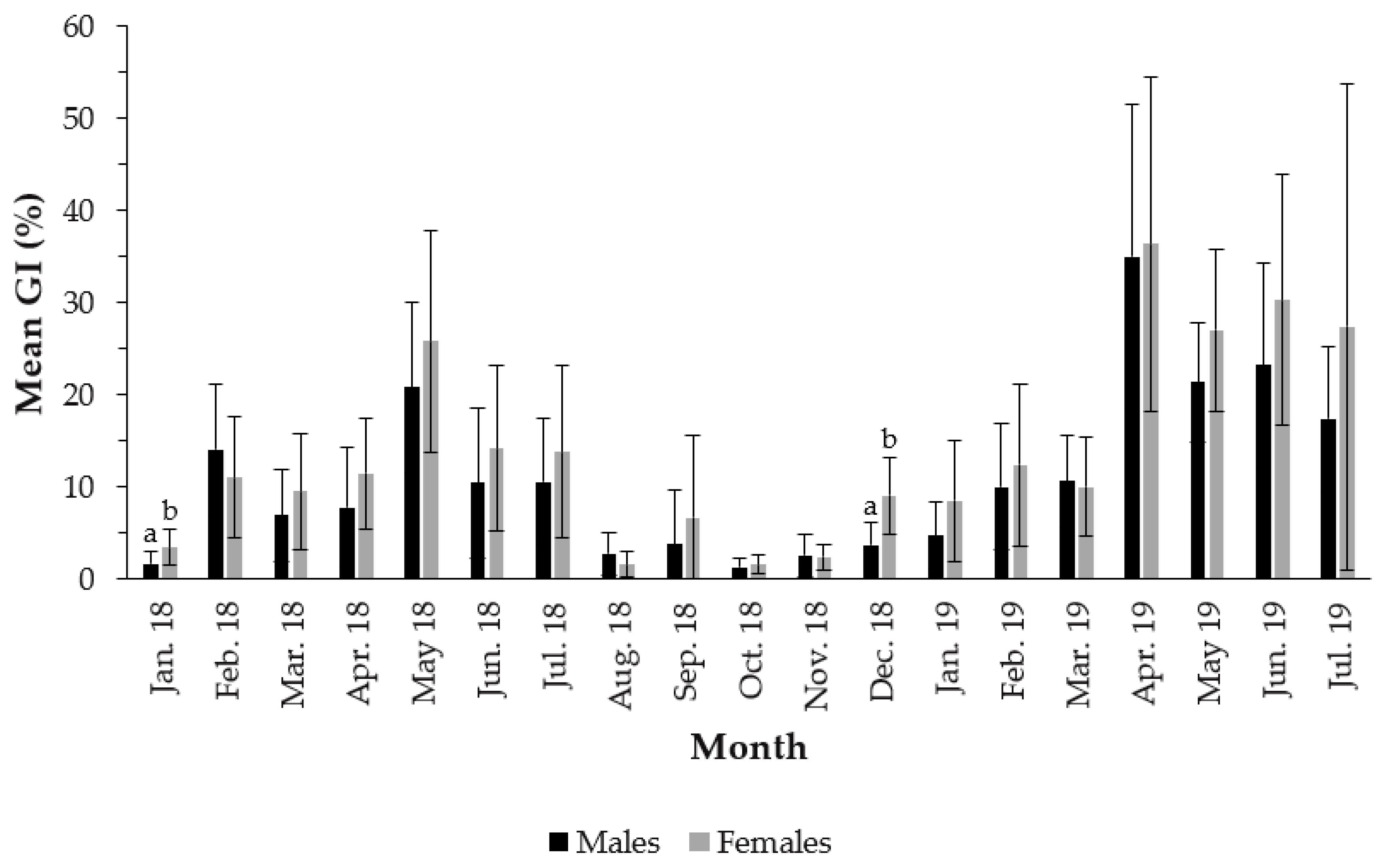
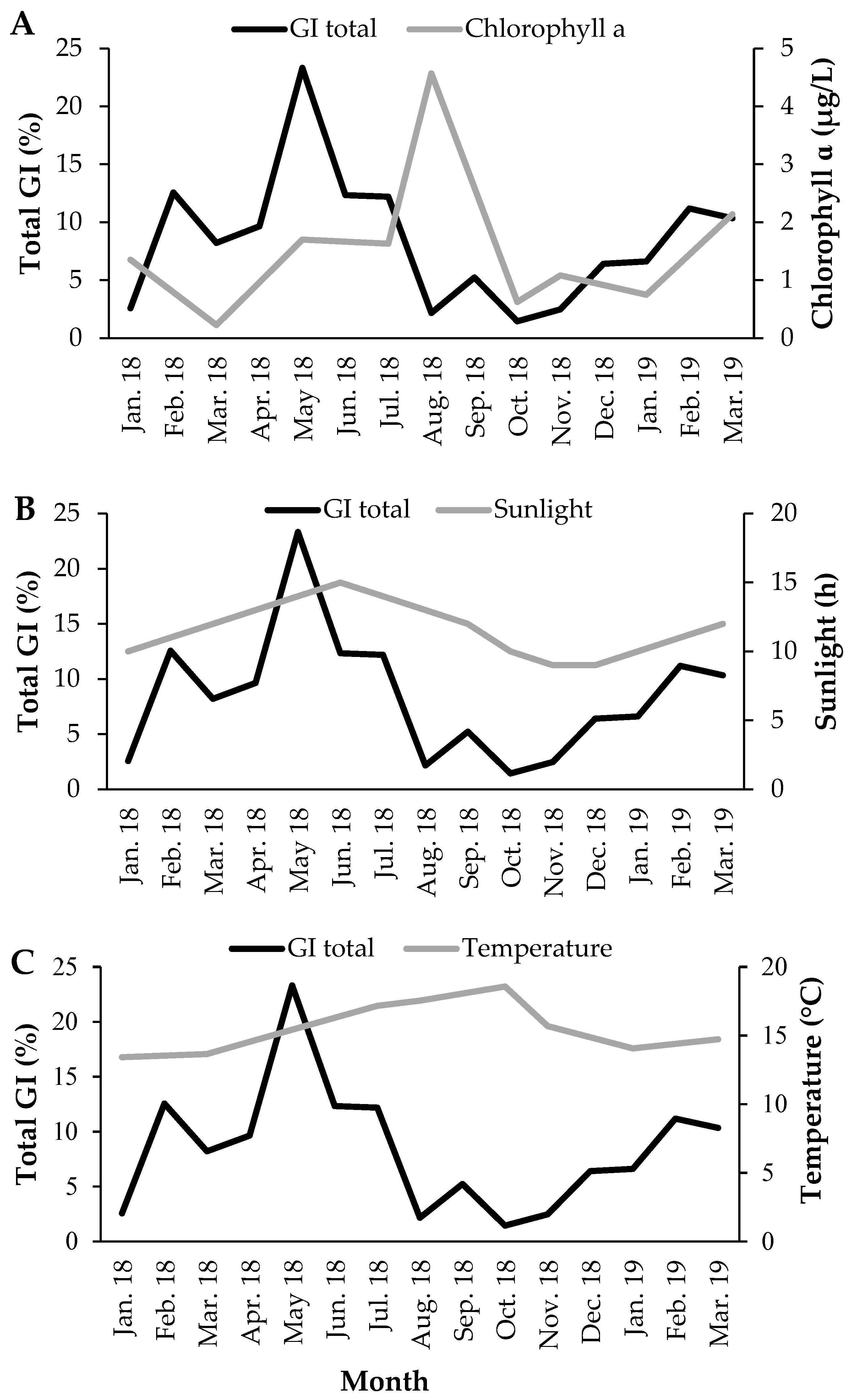
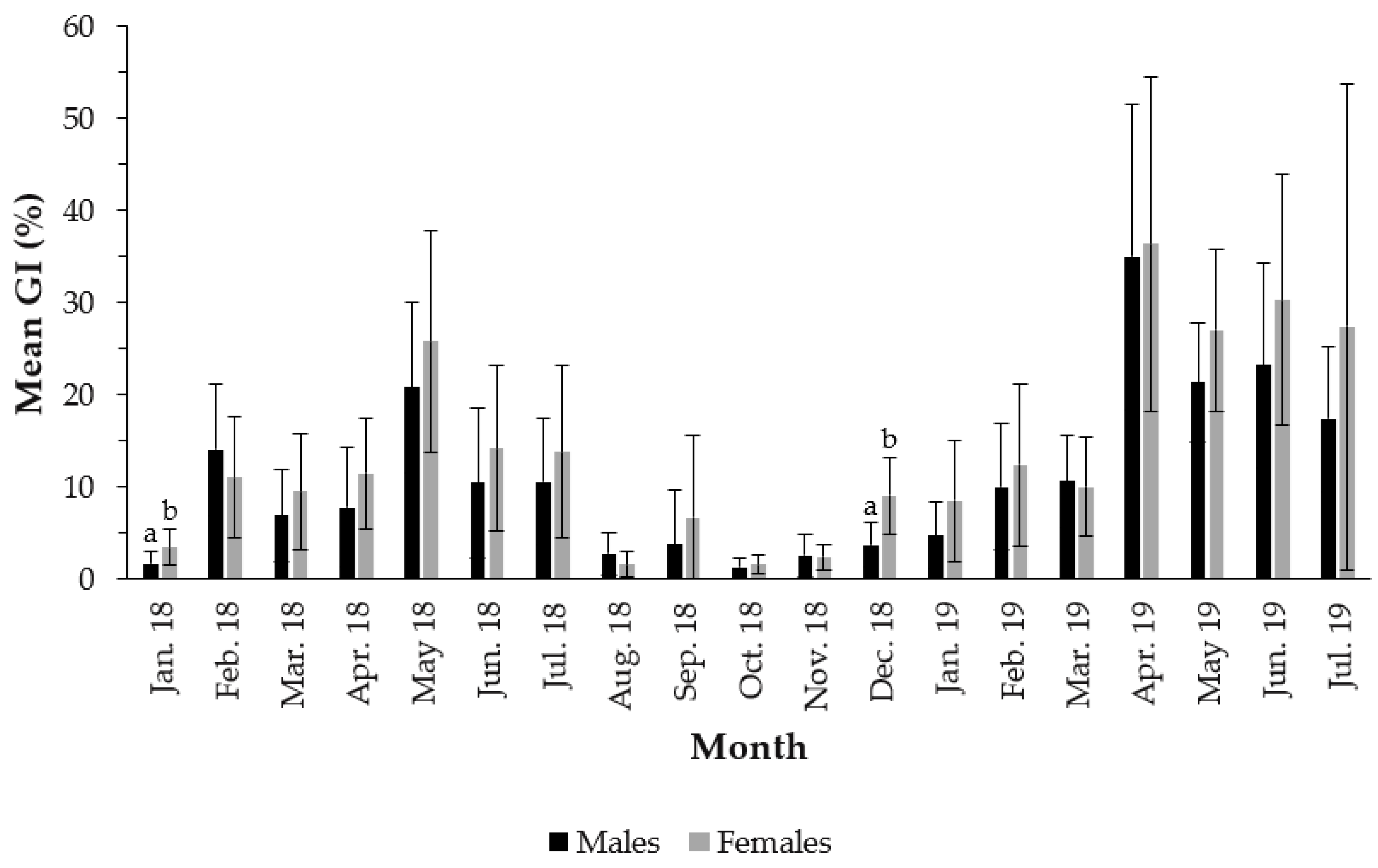
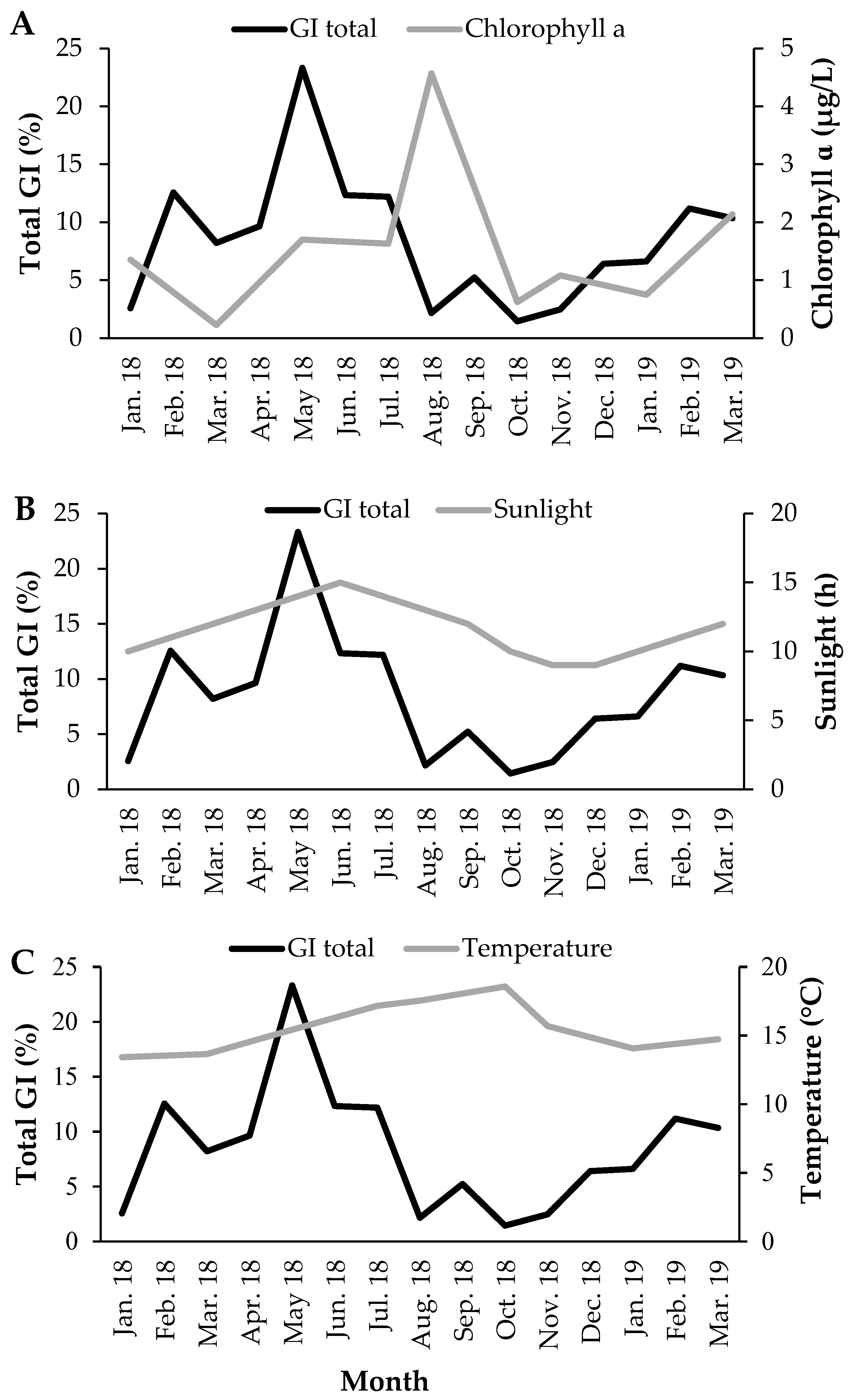
3.2. Gonadosomatic Index
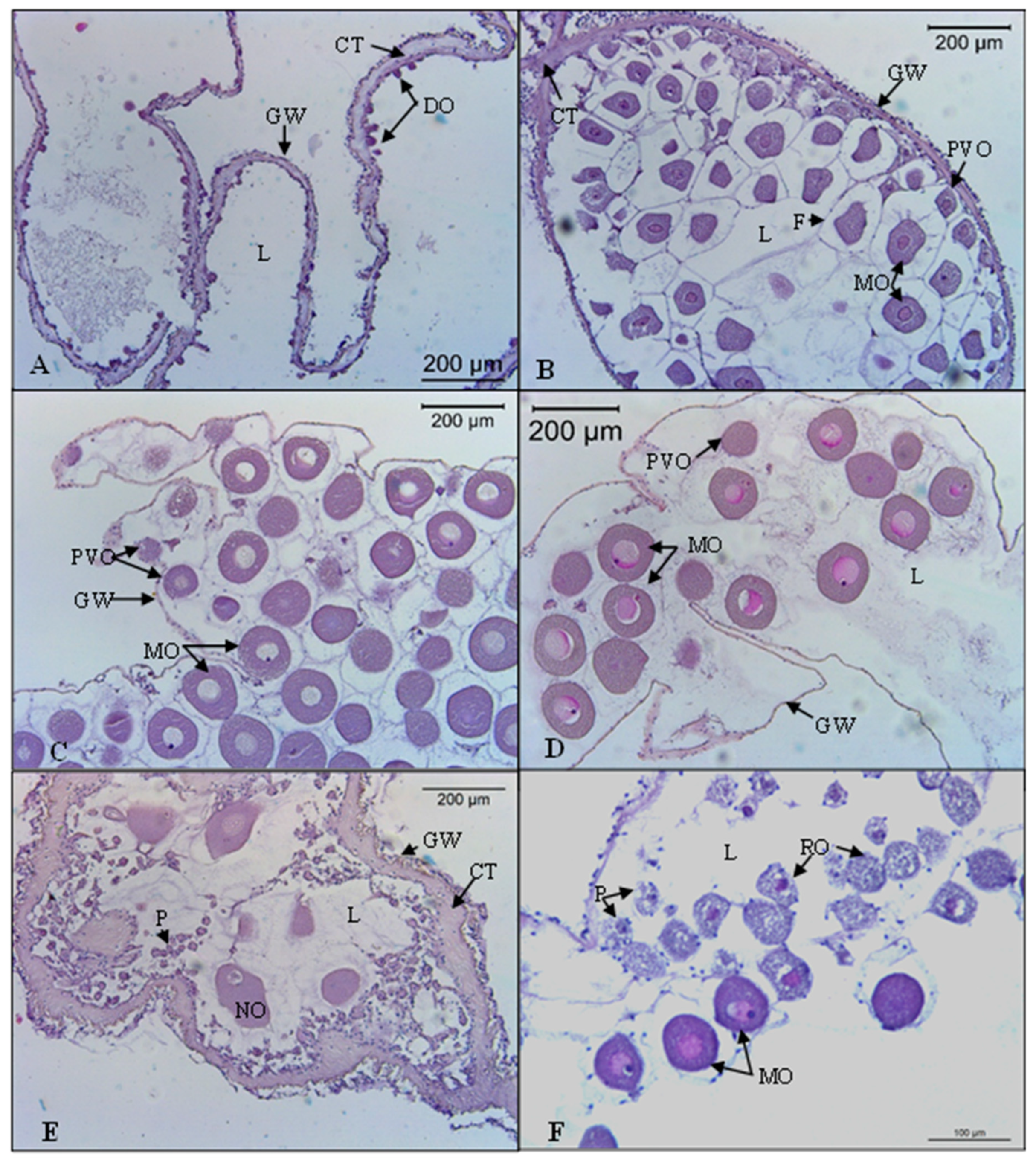
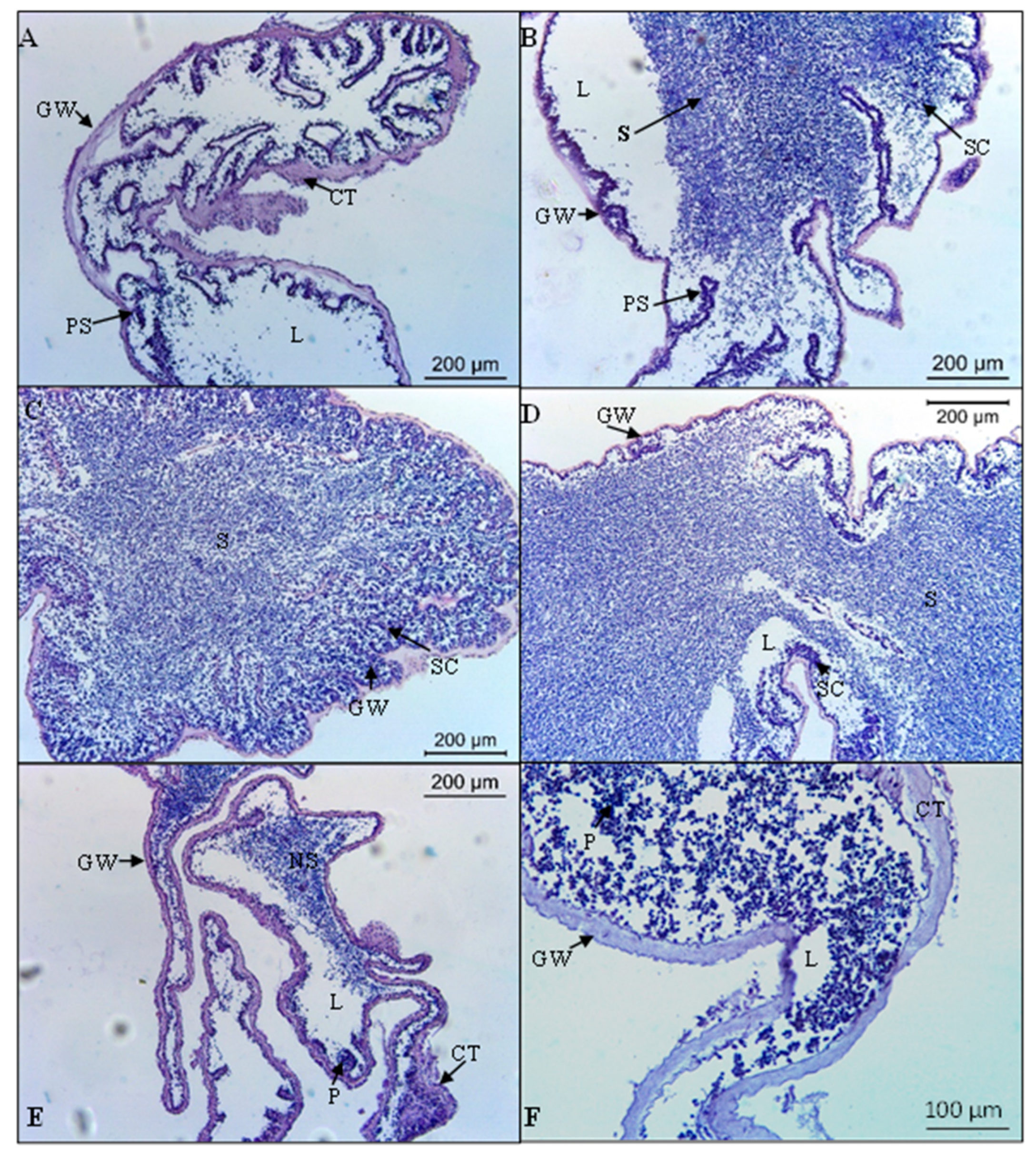
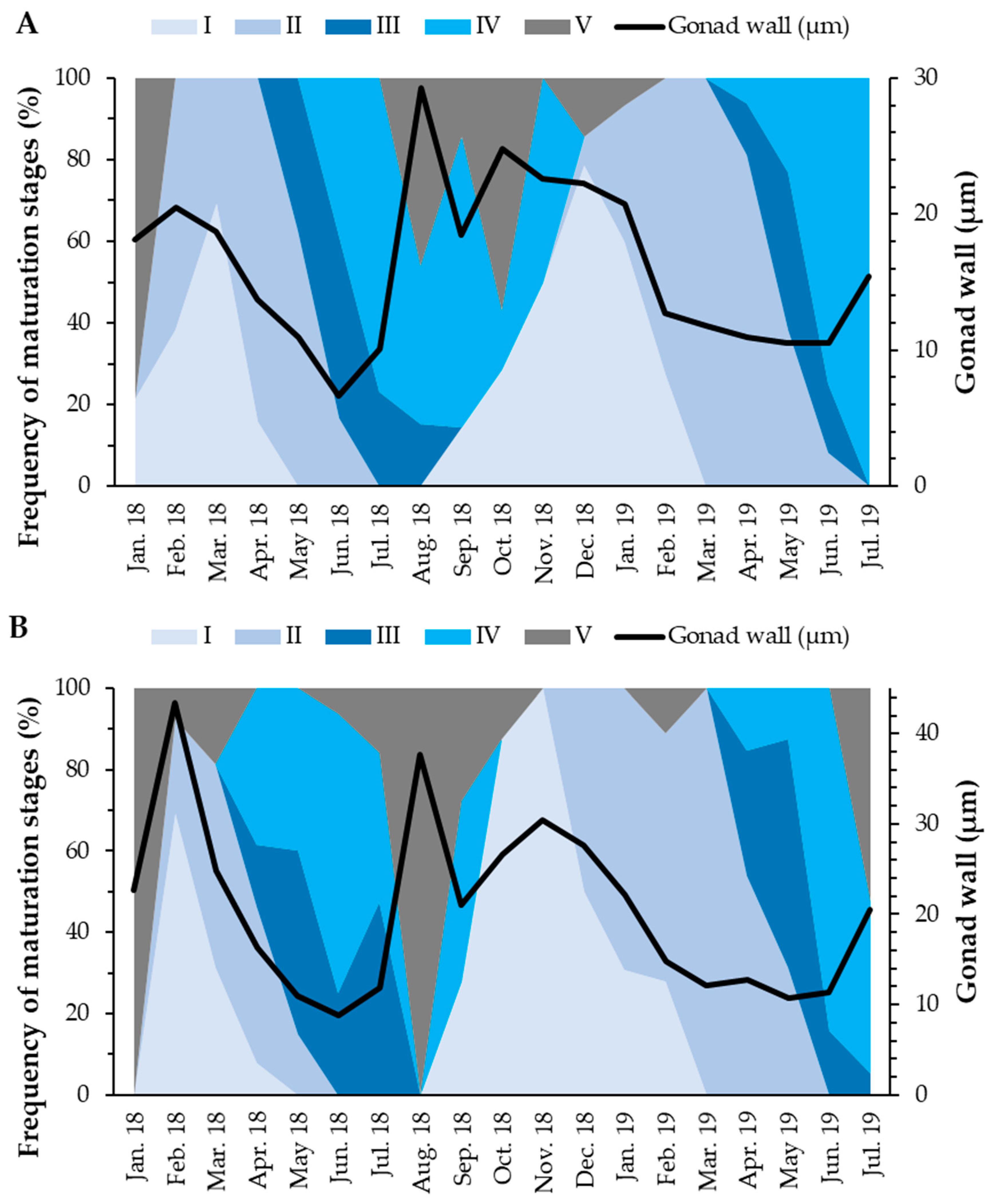
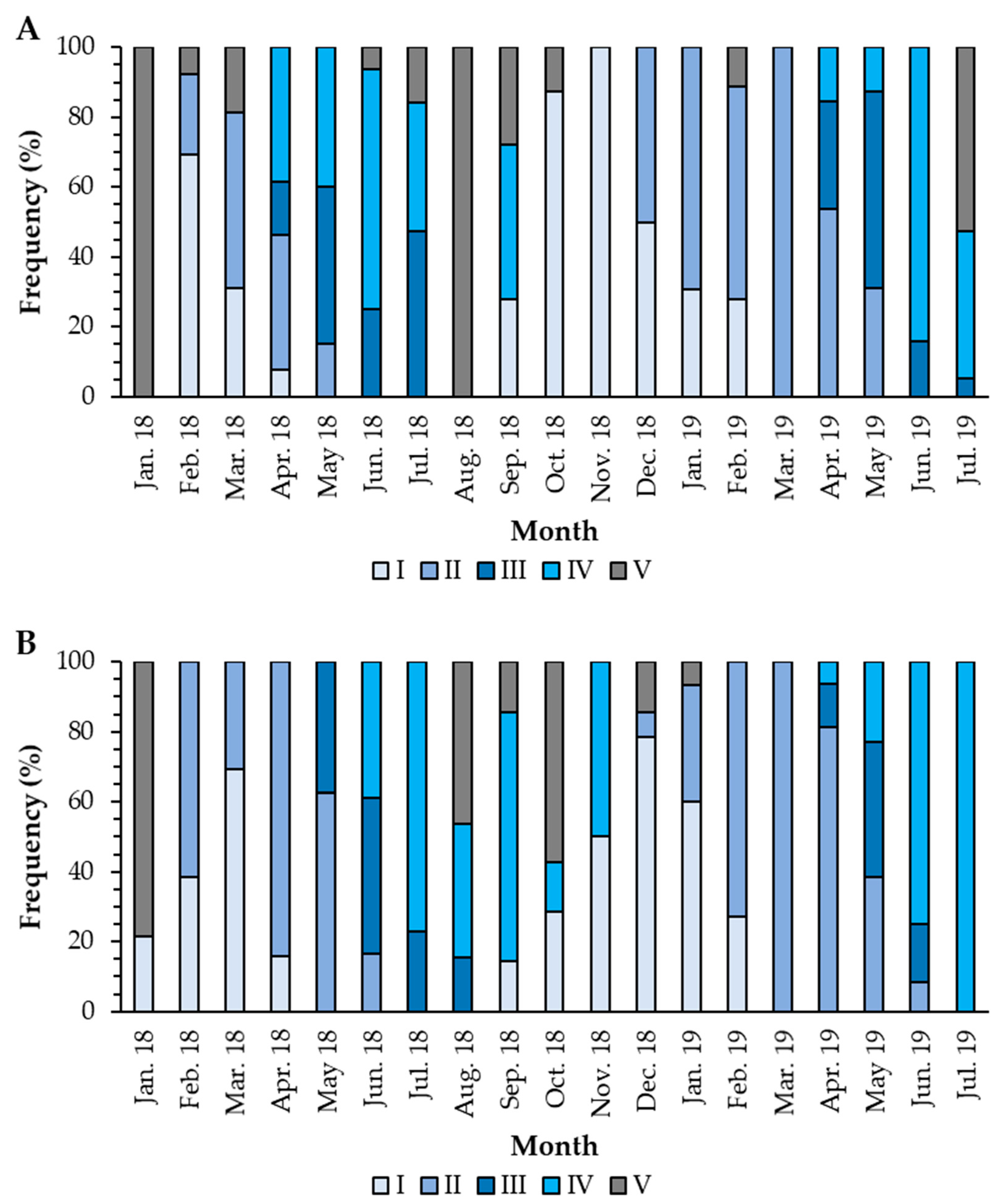
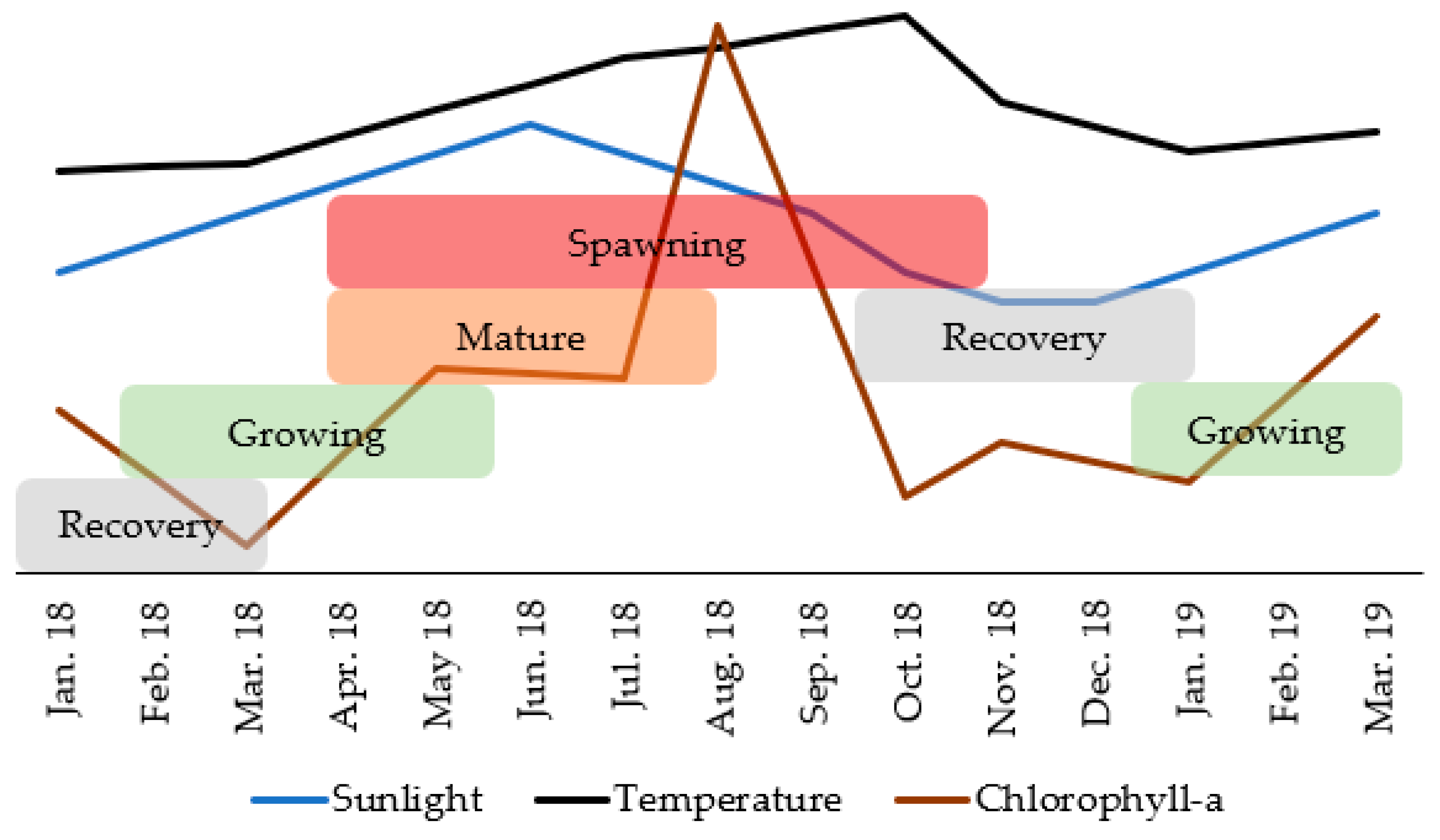
3.3. Reproductive Cycle
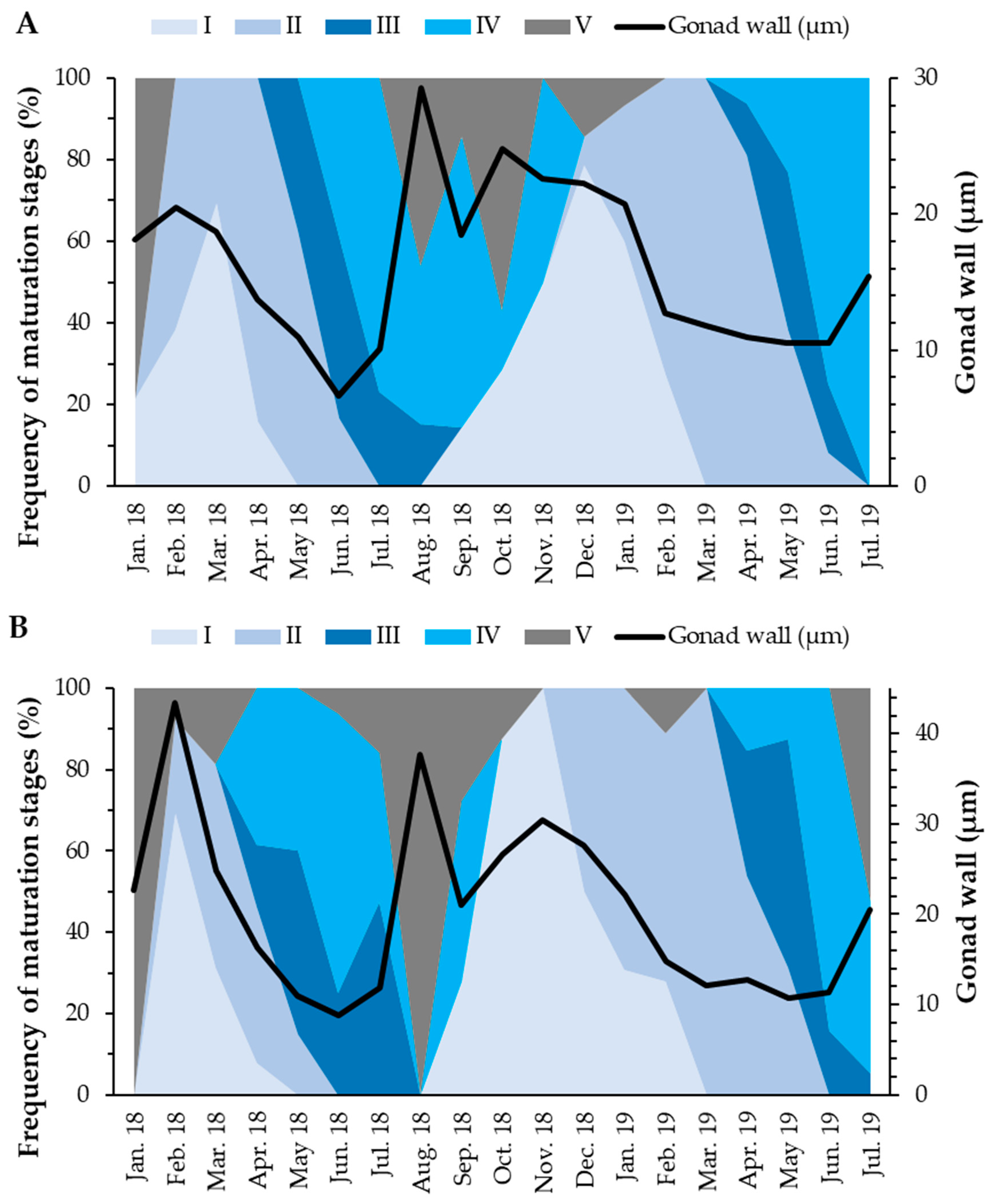
3.3.1. Gonadal Wall Thickness
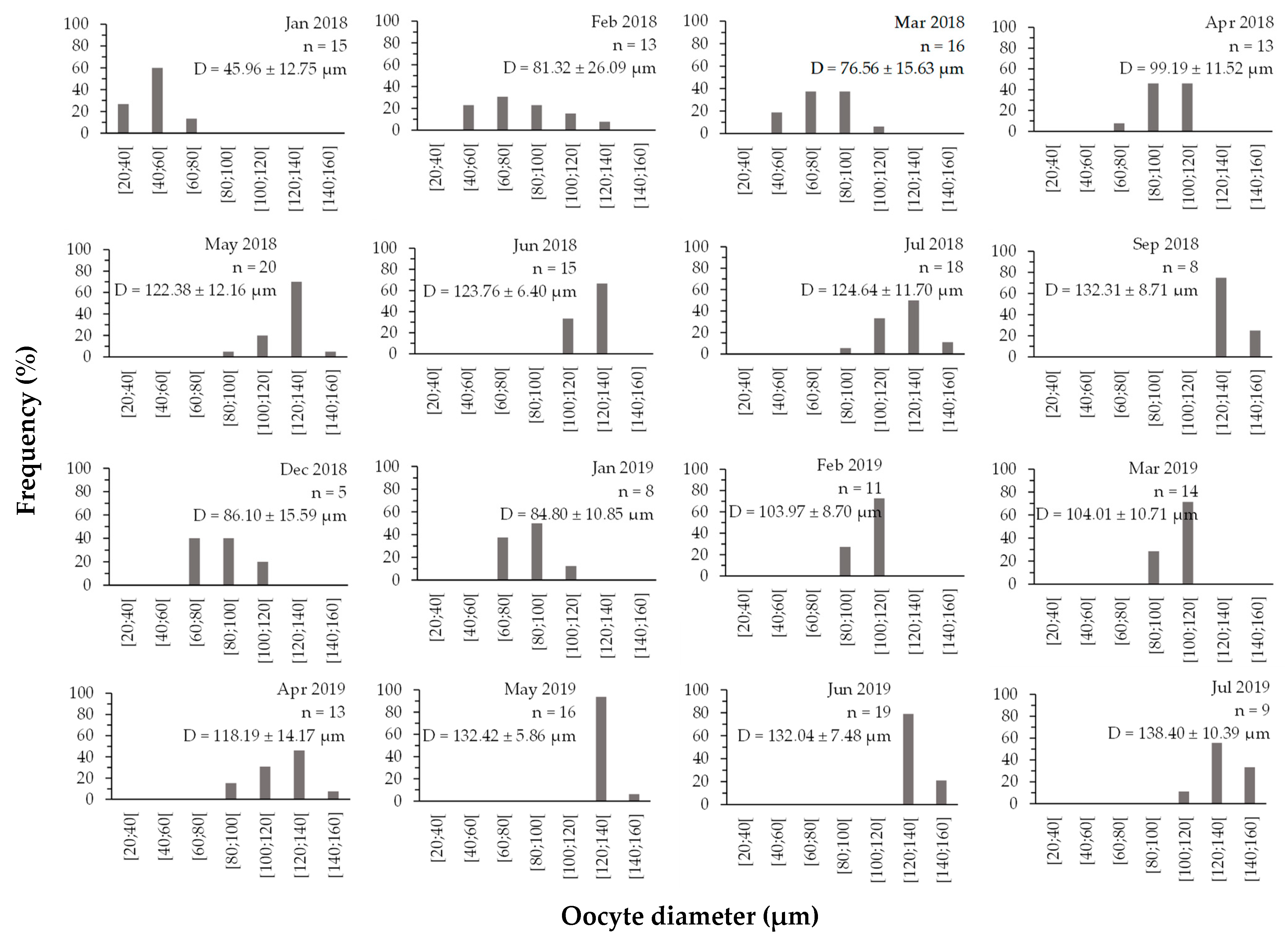
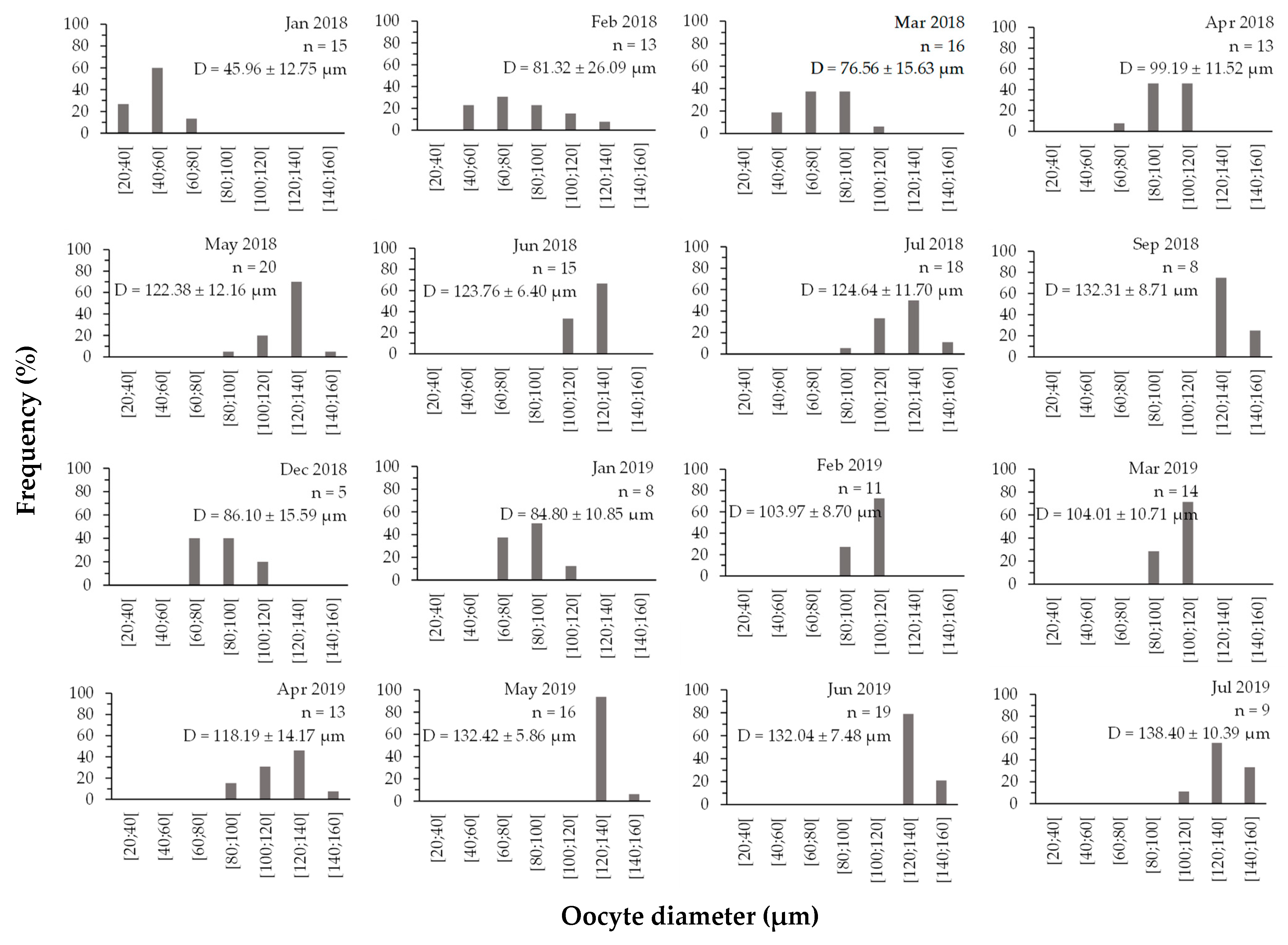
3.3.2. Oocyte Diameter
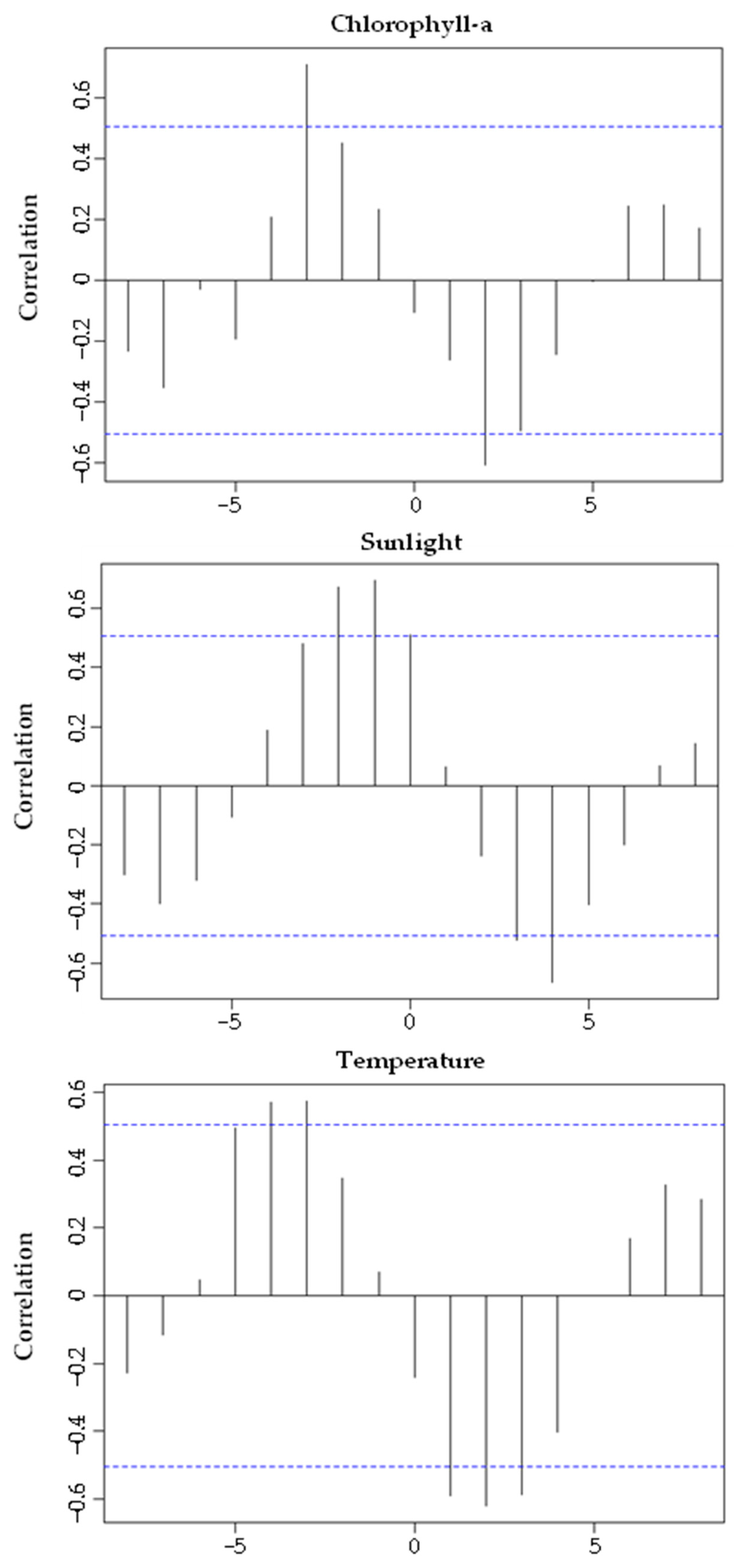
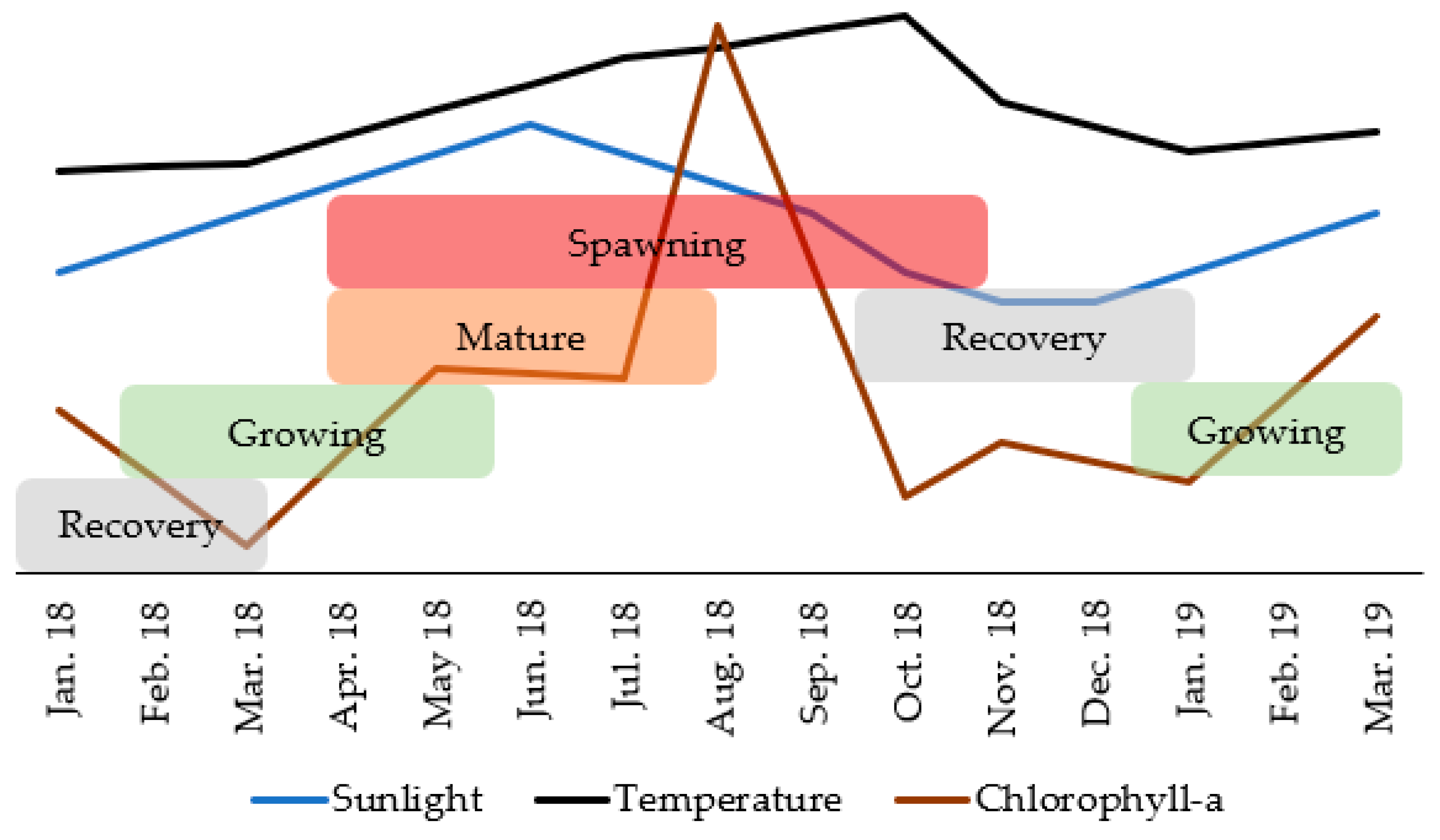
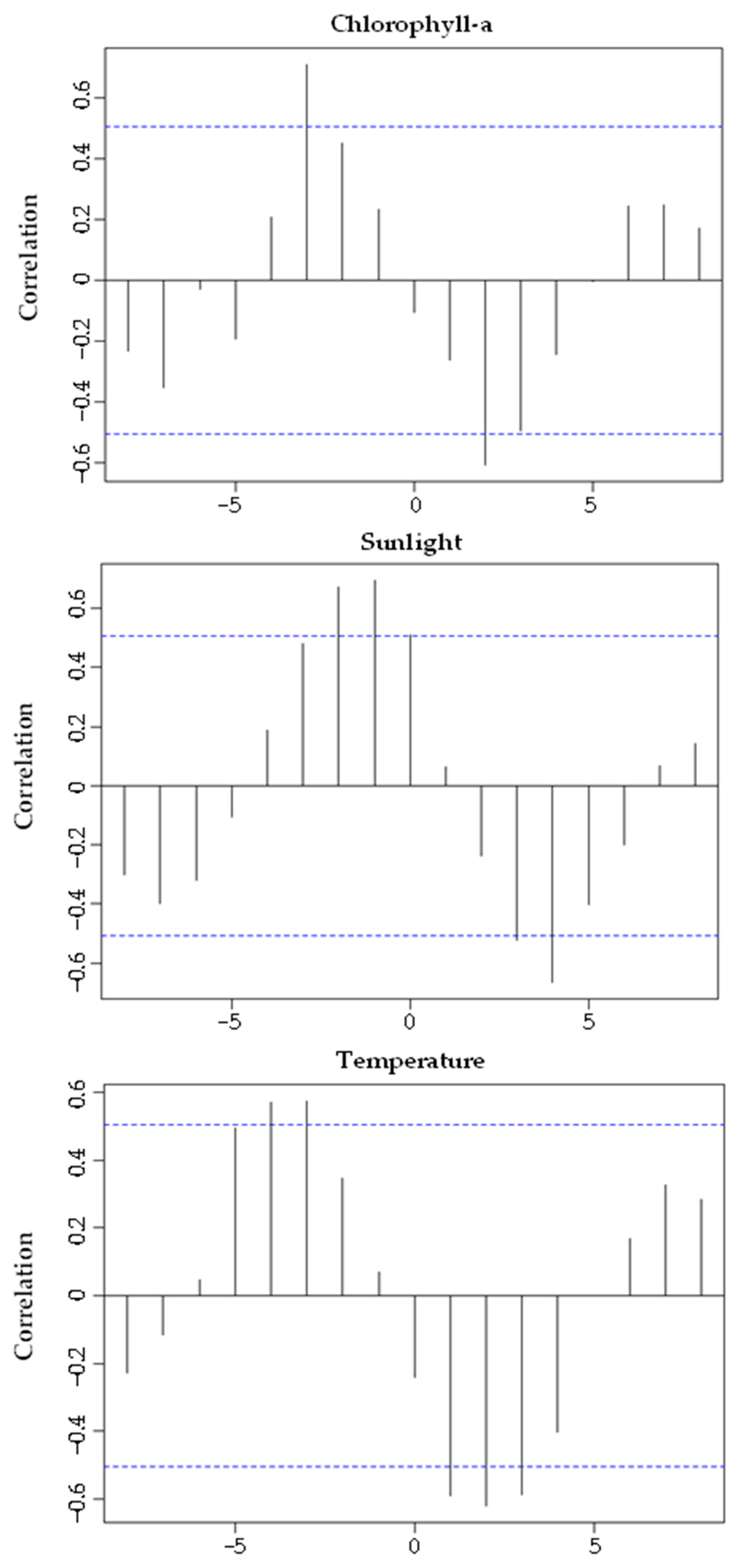
3.4. Reproductive Cycle vs. Environmental Factors
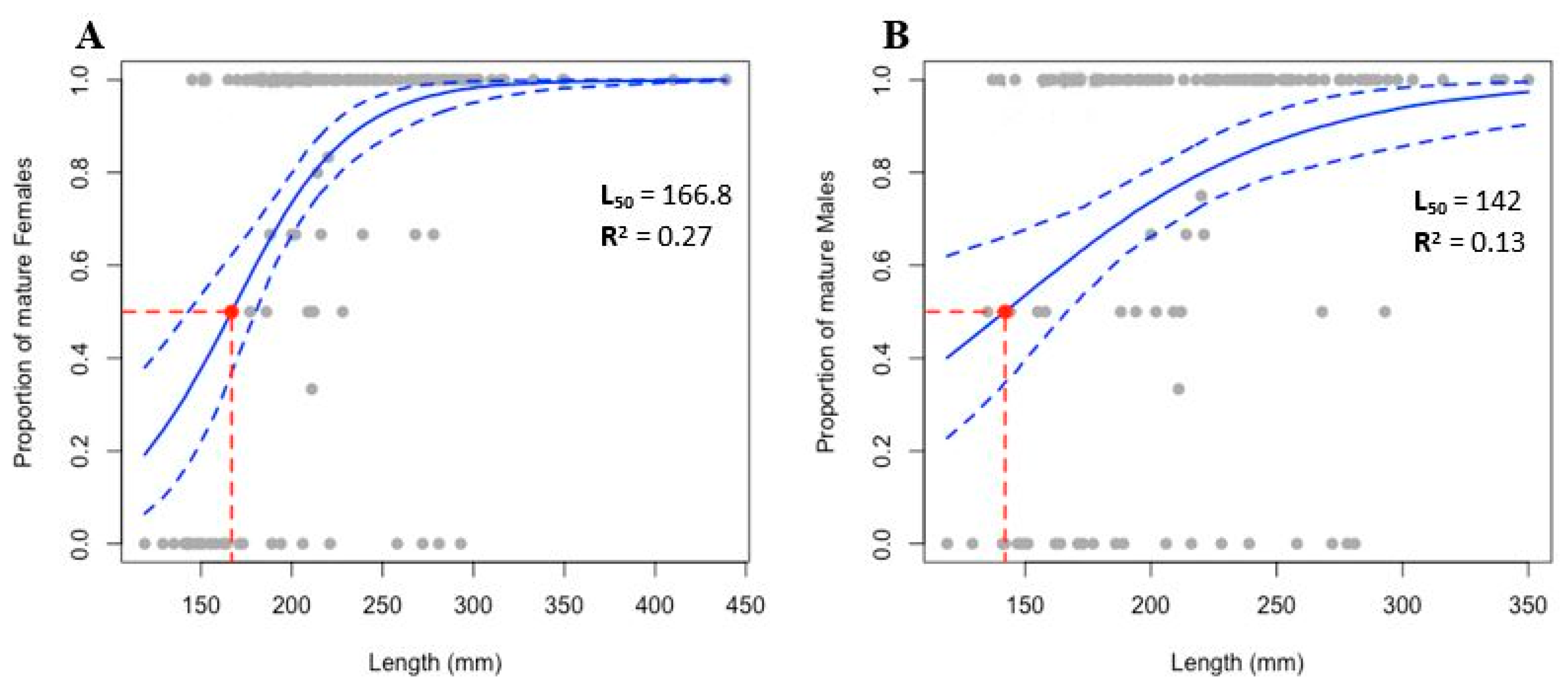
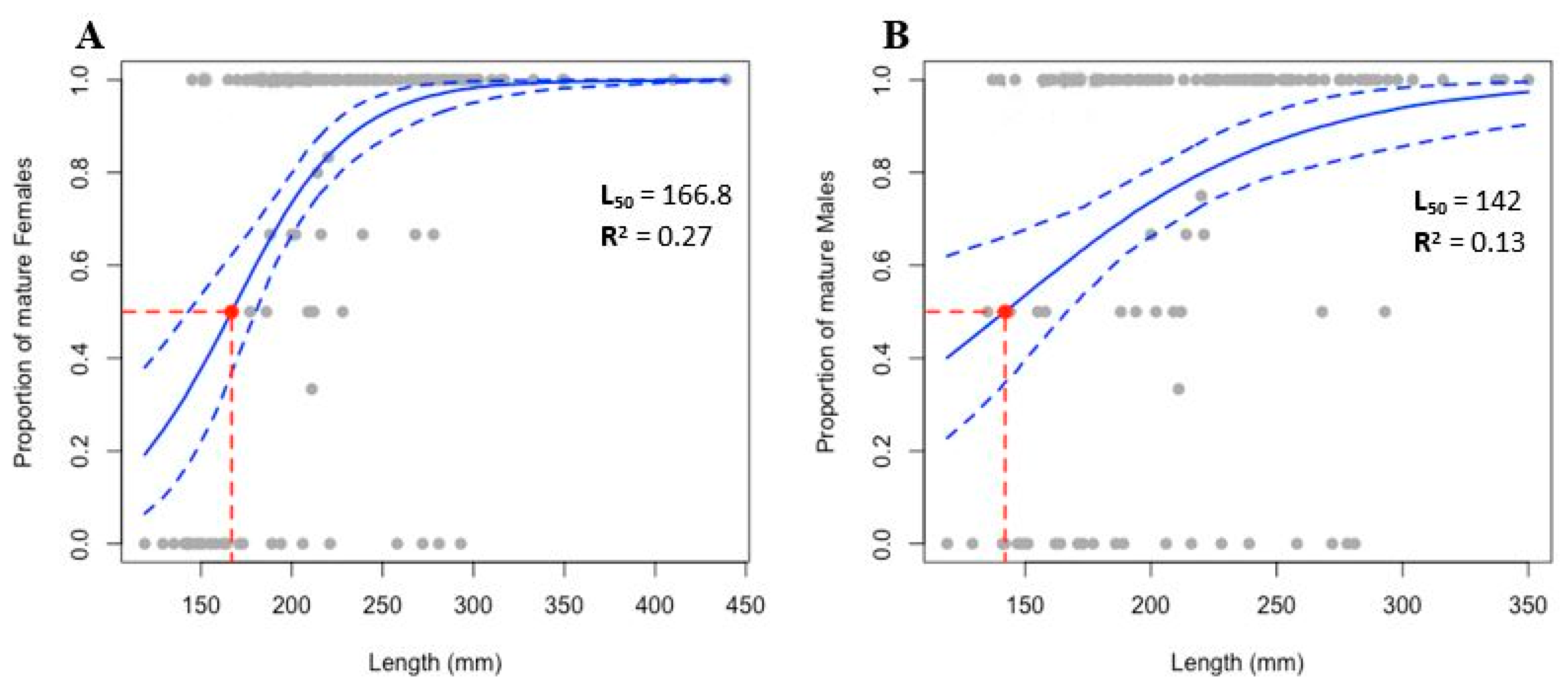
3.5. Size at First Sexual Maturity
4. Discussion
4.1. Sex Ratio
4.2. Reproductive Cycle
4.3. Reproductive Cycle vs. Environmental Factors
4.4. Size at First Sexual Maturity
4.5. Implications for Fisheries and Aquaculture Management
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Conand, C. The Fishery Resources of Pacific Island Countries. Part 2: Holothurians; FAO Fish Technical Paper No. 272.2; FAO: Rome, Italy, 1990; p. 143. [Google Scholar]
- Purcell, S.W. Managing Sea Cucumber Fisheries with an Ecosystem Approach; FAO Fisheries and Aquaculture Technical Paper No. 520; Lovatelli, A., Vasconcellos, M., Ye, Y., Eds.; FAO Fisheries and Aquaculture: Rome, Italy, 2010; p. 157. [Google Scholar]
- Santos, R.; Dias, S.; Pinteus, S.; Silva, J.; Alves, C.; Tecelão, C.; Pedrosa, R.; Pombo, A. Sea cucumber Holothuria forskali, a new resource for aquaculture? Reproductive biology and nutraceutical approach. Aquac. Res. 2015, 47, 2307–2323. [Google Scholar] [CrossRef]
- Purcell, S.W.; Williamson, D.H.; Ngaluafe, P. Chinese market prices of beche-de-mer: Implications for fisheries and aquaculture. Mar. Policy 2018, 91, 58–65. [Google Scholar] [CrossRef]
- Toral-Granda, V.; Lovatelli, A.; Vasconcellos, M. (Eds.) Sea Cucumbers—A Global Review of Fisheries and Trade; FAO Fisheries and Aquaculture Technical Paper No. 516; FAO: Rome, Italy, 2008; p. 317. [Google Scholar]
- Meloni, D.; Esposito, G. Hygienic and commercial issues related to the illegal fishing and processing of sea cucumbers in the Mediterranean: A case study on over-exploitation in Italy between 2015 and 2017. Reg. Stud. Mar. Sci. 2018, 19, 43–46. [Google Scholar] [CrossRef]
- Derelї, H.; Aydın, M. Sea cucumber fishery in Turkey: Management regulations and their efficiency. Reg. Stud. Mar. Sci. 2021, 41, 101551. [Google Scholar] [CrossRef]
- Sonnenholzner, J. Seasonal variation in the food composition of Holothuria theeli (Holothuroidea: Aspidochirotida) with ob-servations on density and distribution patterns at the central coast of Ecuador. Bull. Mar. Sci. 2003, 73, 527–543. [Google Scholar]
- MacTavish, T.; Stenton-Dozey, J.; Vopel, K.; Savage, C. Deposit-Feeding Sea Cucumbers Enhance Mineralization and Nutrient Cycling in Organically-Enriched Coastal Sediments. PLoS ONE 2012, 7, e50031. [Google Scholar] [CrossRef] [Green Version]
- Durden, J.M.; Schoening, T.; Althaus, F.; Friedman, A.; Garcia, R.; Glover, A.G.; Greinert, J.; Stout, N.J.; Jones, D.O.; Jordt, A.; et al. Perspectives in visual imaging for marine biology and ecology: From acquisition to understanding. In Oceanography and Marine Biology: An Annual Review; Hughes, R.N., Hughes, D.J., Smith, I.P., Dale, A.C., Eds.; CRC Press: Boca Raton, FL, USA, 2016; Volume 54, pp. 9–80. [Google Scholar] [CrossRef] [Green Version]
- Venâncio, E.; Félix, P.M.; Brito, A.C.; Sousa, J.; Azevedo e Silva, F.; Simões, T.; Narciso, L.; Amorim, A.; Dâmaso, L.; Pombo, A. Do broodstock diets influence viability and larval development of Holothuria mammata? Aquaculture 2021, 536, 736431. [Google Scholar] [CrossRef]
- Uthicke, S. Interactions between sediment-feeders and microalgae on coral reefs: Grazing losses versus production enhancement. Mar. Ecol. Prog. Ser. 2001, 210, 125–138. [Google Scholar] [CrossRef] [Green Version]
- Uthicke, S. Nutrient regeneration by abundant coral reef holothurians. J. Exp. Mar. Biol. Ecol. 2001, 265, 153–170. [Google Scholar] [CrossRef]
- Purcell, S.W.; Polidoro, B.A.; Hamel, J.-F.; Gamboa, R.U.; Mercier, A. The cost of being valuable: Predictors of extinction risk in marine invertebrates exploited as luxury seafood. Proc. R. Soc. B 2014, 281, 20133296. [Google Scholar] [CrossRef] [Green Version]
- Aydin, M. The commercial sea cucumber fishery in Turkey. SPC Beche-De-Mer Inf. Bull. 2008, 28, 40–41. [Google Scholar]
- González-Wangüemert, M.; Aydin, M.; Conand, C. Assessment of sea cucumber populations from the Aegean Sea (Turkey): First insights to sustainable management of new fisheries. Ocean Coast. Manag. 2014, 92, 87–94. [Google Scholar] [CrossRef]
- González-Wangüemert, M.; Valente, S.; Aydin, M. Effects of fishery protection on biometry and genetic structure of two target sea cucumber species from the Mediterranean Sea. Hydrobiologia 2014, 743, 65–74. [Google Scholar] [CrossRef] [Green Version]
- Gaudron, S.M.; Kohler, S.A.; Conand, C. Reproduction of the sea cucumber Holothuria leucospilota in the Western Indian Ocean: Biological and ecological aspects. Invertebr. Reprod. Dev. 2008, 51, 19–31. [Google Scholar] [CrossRef]
- Dissanayake, D.C.T.; Stefansson, G. Reproductive biology of the commercial sea cucumber Holothuria atra (Holothuroidea: Aspidochirotida) in the northwestern coastal waters of Sri Lanka. Invertebr. Reprod. Dev. 2010, 54, 65–76. [Google Scholar] [CrossRef]
- Omar, H.A.; Abdel Razek, F.A.; Abdel Rahman, S.H. Reproductive periodicity of sea cucumber Bohadschia vitiensis (Echinodermata: Holothuroidea) in Hurghada area, Red Sea, Egypt. Egypt. J. Aquat. Res. 2013, 39, 115–123. [Google Scholar] [CrossRef] [Green Version]
- Kubota, T.; Tomari, M. Reproduction in the Apodid Sea Cucumber Polycheira Rufescens: Semilunar Spawning Rhythm and Sex Change. J. Mar. Biol. Assoc. United Kingd. 1998, 78, 249–267. [Google Scholar] [CrossRef]
- Mercier, A.; Ycaza, R.; Hamel, J. Long-term study of gamete release in a broadcast-spawning holothurian: Predictable lunar and diel periodicities. Mar. Ecol. Prog. Ser. 2007, 329, 179–189. [Google Scholar] [CrossRef]
- Hamel, J.-F.; Himmelman, J.H.; Dufresne, L. Gametogenesis and Spawning of the Sea Cucumber Psolus fabricii (Duben and Koren). Biol. Bull. 1993, 184, 125–143. [Google Scholar] [CrossRef]
- Wigham, B.; Hudson, I.R.; Billett, D.S.M.; Wolff, G.A. Is long-term change in the abyssal Northeast Atlantic driven by qualitative changes in export flux? Evidence from selective feeding in deep-sea holothurians. Prog. Oceanogr. 2003, 59, 409–441. [Google Scholar] [CrossRef]
- Hamel, J.-F.; Mercier, A. Early development, settlement, growth, and spatial distribution of the sea cucumber Cucumaria frondosa (Echinodermata: Holothuroidea). Can. J. Fish. Aquat. Sci. 1996, 53, 253–271. [Google Scholar] [CrossRef]
- Marquet, N.; Conand, C.; Power, D.M.; Canário, A.V.M.; González-Wangüemert, M. Sea cucumbers, Holothuria arguinensis and H. mammata, from the southern Iberian Peninsula: Variation in reproductive activity between populations from different habitats. Fish. Res. 2017, 191, 120–130. [Google Scholar] [CrossRef]
- Santos, R.; Dias, S.; Tecelão, C.; Pedrosa, R.; Pombo, A. Reproductive biological characteristics and fatty acid profile of Holo-thuria mammata (Grube, 1840). SPC Beche-De-Mer Inf. Bull. 2017, 37, 58–64. [Google Scholar]
- Azevedo e Silva, F.; Brito, A.C.; Simões, T.; Pombo, A.; Marques, T.A.; Rocha, C.; Sousa, J.; Venâncio, E.; Félix, P.M. Allometric relationships to assess ontogenetic adaptative changes in three NE Atlantic commercial sea cucumbers (Echinodermata, Holothuroidea). Aquat. Ecol. 2021, 55, 711–720. [Google Scholar] [CrossRef]
- Benítez-Villalobos, F.; Avila-Poveda, O.H.; Gutiérrez-Méndez, I.S. Reproductive biology of Holothuria fuscocinerea (Echinodermata: Holothuroidea) from Oaxaca, Mexico. Sex. Early Dev. Aquat. Org. 2013, 1, 13–24. [Google Scholar] [CrossRef] [Green Version]
- Despalatović, M.; Grubelić, I.; Šimunović, A.; Antolić, B.; Žuljević, A. Reproductive biology of the holothurian Holothuria tubulosa (Echinodermata) in the Adriatic Sea. J. Mar. Biol. Assoc. United Kingd. 2004, 84, 409–414. [Google Scholar] [CrossRef]
- Kazanidis, G.; Lolas, A.; Vafidis, D. Reproductive cycle of the traditionally exploited sea cucumber Holothuria tubulosa (Holothuroidea: Aspidochirotida) in Pagasitikos Gulf, western Aegean Sea, Greece. Turk. J. Zoӧl. 2014, 38, 306–315. [Google Scholar] [CrossRef]
- Sousa, J.; Félix, P.M.; Brito, A.C.; Venâncio, E.; Azevedo e Silva, F.; Simões, T.; Raposo, A.; Neves, M.; Narciso, L.; Melo, R.; et al. The effects of stocking density on physiological traits in Holothuria forskali broodstock. Aquac. Res. 2021, 52, 1548–1560. [Google Scholar] [CrossRef]
- Chernick, M.R.; Liu, C.Y. The Saw-Toothed Behavior of Power Versus Sample Size and Software Solutions. Am. Stat. 2002, 56, 149–155. [Google Scholar] [CrossRef]
- Engels, W.R. Exact Tests for Hardy-Weinberg Proportions. Genetics 2009, 183, 1431–1441. [Google Scholar] [CrossRef] [Green Version]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: http://www.R-project.org/ (accessed on 15 January 2022).
- Torrejon-Magallanes, J. Package “sizeMat”, Version 1.1.2; Estimate Size at Sexual Maturity. Repository CRAN 2020; Available online: http://CRAN.R-project.org/package=sizeMat (accessed on 15 January 2022).
- Shiell, G.R.; Uthicke, S. Reproduction of the commercial sea cucumber Holothuria whitmaei [Holothuroidea: Aspidochirotida] in the Indian and Pacific Ocean regions of Australia. Mar. Biol. 2006, 148, 973–986. [Google Scholar] [CrossRef]
- Muthiga, N.A. The reproductive biology of a new species of sea cucumber, Holothuria (Mertensiothuria) arenacava in a Kenyan marine protected area: The possible role of light and temperature on gametogenesis and spawning. Mar. Biol. 2006, 149, 585–593. [Google Scholar] [CrossRef]
- Muthiga, N.A.; Kawaka, J.A.; Ndirangu, S. The timing and reproductive output of the commercial sea cucumber Holothuria scabra on the Kenyan coast. Estuar. Coast. Shelf Sci. 2009, 84, 353–360. [Google Scholar] [CrossRef]
- Ramos-Miranda, J.; del Rio Rodriguez, R.E.; Flores-Hernández, D.; Rojas-González, R.I.; Gomez-Solano, M.I.; Cu-Escamilla, A.D.; Gómez-Criollo, F.; Sosa-Lopez, A.; Torres-Rojas, Y.E.; Juárez-Camargo, P. Reproductive cycle of the sea cucumber Holothuria floridana in the littorals of Campeche, Mexico. Fish. Sci. 2017, 83, 699–714. [Google Scholar] [CrossRef]
- Félix, P.M.; Pombo, A.; Azevedo e Silva, F.; Simões, T.; Marques, T.A.; Melo, R.; Rocha, C.; Sousa, J.; Venâncio, E.; Costa, J.L.; et al. Modelling the Distribution of a Commercial NE-Atlantic Sea Cucumber, Holothuria mammata: Demographic and Abundance Spatio-Temporal Patterns. Front. Mar. Sci. 2021, 8, 675330. [Google Scholar] [CrossRef]
- Tuwo, A.; Conand, C. Reproductive biology of the holothurian Holothuria forskali (Echinodermata). J. Mar. Biol. Assoc. United Kingd. 1992, 72, 745–758. [Google Scholar] [CrossRef]
- Conand, C. Reproductive biology of the holothurians from the major communities of the New Caledonian Lagoon. Mar. Biol. 1993, 116, 439–450. [Google Scholar] [CrossRef]
- Navarro, P.G.; García-Sanz, S.; Tuya, F. Reproductive biology of the sea cucumber Holothuria sanctori (Echinodermata: Holothuroidea). Sci. Mar. 2011, 76, 741–752. [Google Scholar] [CrossRef] [Green Version]
- Ramofafia, C.; Byrne, M.; Battaglene, C. Reproduction of the commercial sea cucumber Holothuria scabra (Echinodermata: Holothuroidea) in the Solomon Islands. Mar. Biol. 2003, 142, 281–288. [Google Scholar] [CrossRef]
- Ramofafia, C.; Battaglene, S.C.; Bell, J.D.; Byrne, M. Reproductive biology of the commercial sea cucumber Holothuria fuscogilva in the Solomon Islands. Mar. Biol. 2000, 136, 1045–1056. [Google Scholar] [CrossRef]
- Foglietta, L.M.; Camejo, M.I.; Gallardo, L.; Herrera, F.C. A maturity index for holothurians exhibiting asynchronous development of gonad tubules. J. Exp. Mar. Biol. Ecol. 2004, 303, 19–30. [Google Scholar] [CrossRef]
- Ramofafia, C.; Byrne, M.; Battaglene, S. Reproductive biology of the intertidal sea cucumber Actinopyga mauritiana in the Solomon Islands. J. Mar. Biol. Assoc. United Kingd. 2001, 81, 523–531. [Google Scholar] [CrossRef]
- Leite-Castro, L.V.; Junior, J.D.S.; Salmito-Vanderley, C.S.B.; Nunes, J.F.; Hamel, J.-F.; Mercier, A. Reproductive biology of the sea cucumber Holothuria grisea in Brazil: Importance of social and environmental factors in breeding coordination. Mar. Biol. 2016, 163, 1–13. [Google Scholar] [CrossRef]
- Boidron-Métairon, I.F. Larval Nutrition. In Ecology of Marine Invertebrate Larvae; McEdward, L., Ed.; CRC Press: Boca Raton, FL, USA, 1995; pp. 223–248. Available online: http://www.taylorfrancis.com/chapters/edit/10.1201/9780138758950-7/larval-nutrition-isabelle-boidron-métairon (accessed on 3 November 2021).
- Starr, M.; Himmelman, J.H.; Therriault, J.-C. Direct Coupling of Marine Invertebrate Spawning with Phytoplankton Blooms. Science 1990, 247, 1071–1074. [Google Scholar] [CrossRef] [PubMed]
- Watson, G.J.; Bentley, M.G.; Gaudron, S.M.; Hardege, J.D. The role of chemical signals in the spawning induction of polychaete worms and other marine invertebrates. J. Exp. Mar. Biol. Ecol. 2003, 294, 169–187. [Google Scholar] [CrossRef]
- Domínguez-Godino, J.A.; González-Wangüemert, M. Breeding and larval development of Holothuria mammata, a new target species for aquaculture. Aquac. Res. 2018, 49, 1430–1440. [Google Scholar] [CrossRef]
- Conand, C. Sexual cycle of three commercially important Holothurian species (Echinodermata) from the Lagoon of New Caledonia. Bull. Mar. Sci. 1981, 31, 523–543. [Google Scholar]
- Abdel-Razek, F.A.; Abdel-Rahmen, S.H.; El-Shimy, N.A.; Omar, H.A. Reproductive biology of the tropical sea cucumber Holothuria atra (Echinodermata:Holothuroidea) in the Red Sea coast of Egypt. Egypt. J. Aquat. Res. 2005, 31, 383–402. [Google Scholar]
- Hasan, M.H. Destruction of a Holothuria scabra population by overfishing at Abu Rhamada Island in the Red Sea. Mar. Environ. Res. 2005, 60, 489–511. [Google Scholar] [CrossRef]
- Hashemi, A.; Taghavimotlagh, S.A.; Aminrad, T.; Gaudron, S. Length at First Maturity and Spawning Time of Holothuria Leu-cospilota (Brandt, 1835) in the Northern Waters of the Oman Sea (Sistan and Baluchestan Province). J. Mar. Biol. 2020, 12, 93–105. Available online: http://jmb.iauahvaz.ac.ir/article-1-862-en.html (accessed on 17 January 2022).
- Acosta, E.J.; Rodríguez-Forero, A.; Werding, B.; Kunzmann, A. Ecological and reproductive characteristics of holothuroids Isostichopus badionotus and Isostichopus sp. in Colombia. PLoS ONE 2021, 16, e0247158. [Google Scholar] [CrossRef] [PubMed]
- Liu, O.R.; Thomas, L.R.; Clemence, M.; Fujita, R.; Kritzer, J.P.; McDonald, G.; Szuwalski, C. An Evaluation of Harvest Control Methods for Fishery Management. Rev. Fish. Sci. Aquac. 2016, 24, 244–263. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Venâncio, E.; Félix, P.M.; Brito, A.C.; Azevedo e Silva, F.; Simões, T.; Sousa, J.; Mendes, S.; Pombo, A. Reproductive Biology of the Sea Cucumber Holothuria mammata (Echinodermata: Holothuroidea). Biology 2022, 11, 622. https://doi.org/10.3390/biology11050622
Venâncio E, Félix PM, Brito AC, Azevedo e Silva F, Simões T, Sousa J, Mendes S, Pombo A. Reproductive Biology of the Sea Cucumber Holothuria mammata (Echinodermata: Holothuroidea). Biology. 2022; 11(5):622. https://doi.org/10.3390/biology11050622
Chicago/Turabian StyleVenâncio, Eliana, Pedro M. Félix, Ana C. Brito, Francisco Azevedo e Silva, Tomás Simões, João Sousa, Susana Mendes, and Ana Pombo. 2022. "Reproductive Biology of the Sea Cucumber Holothuria mammata (Echinodermata: Holothuroidea)" Biology 11, no. 5: 622. https://doi.org/10.3390/biology11050622
APA StyleVenâncio, E., Félix, P. M., Brito, A. C., Azevedo e Silva, F., Simões, T., Sousa, J., Mendes, S., & Pombo, A. (2022). Reproductive Biology of the Sea Cucumber Holothuria mammata (Echinodermata: Holothuroidea). Biology, 11(5), 622. https://doi.org/10.3390/biology11050622