
Clostridium perfringens Associated with Foodborne Infections of Animal Origins: Insights into Prevalence, Antimicrobial Resistance, Toxin Genes Profiles, and Toxinotypes

 ,
,  ,
,  , , , and
, , , and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Isolation, Enumeration and Phenotypic Identification of C. perfringens
2.3. Genotypic Characterization of C. perfringens Isolates
2.4. Antimicrobial Susceptibility Testing
2.5. Typing of C. perfringens Toxins
2.6. Molecular Detection of C. perfringens Toxin and Antimicrobial Resistance Genes
2.7. Statistical Analyses
3. Results
3.1. Prevalence, Loads, and Characterization of C. perfringens Isolates
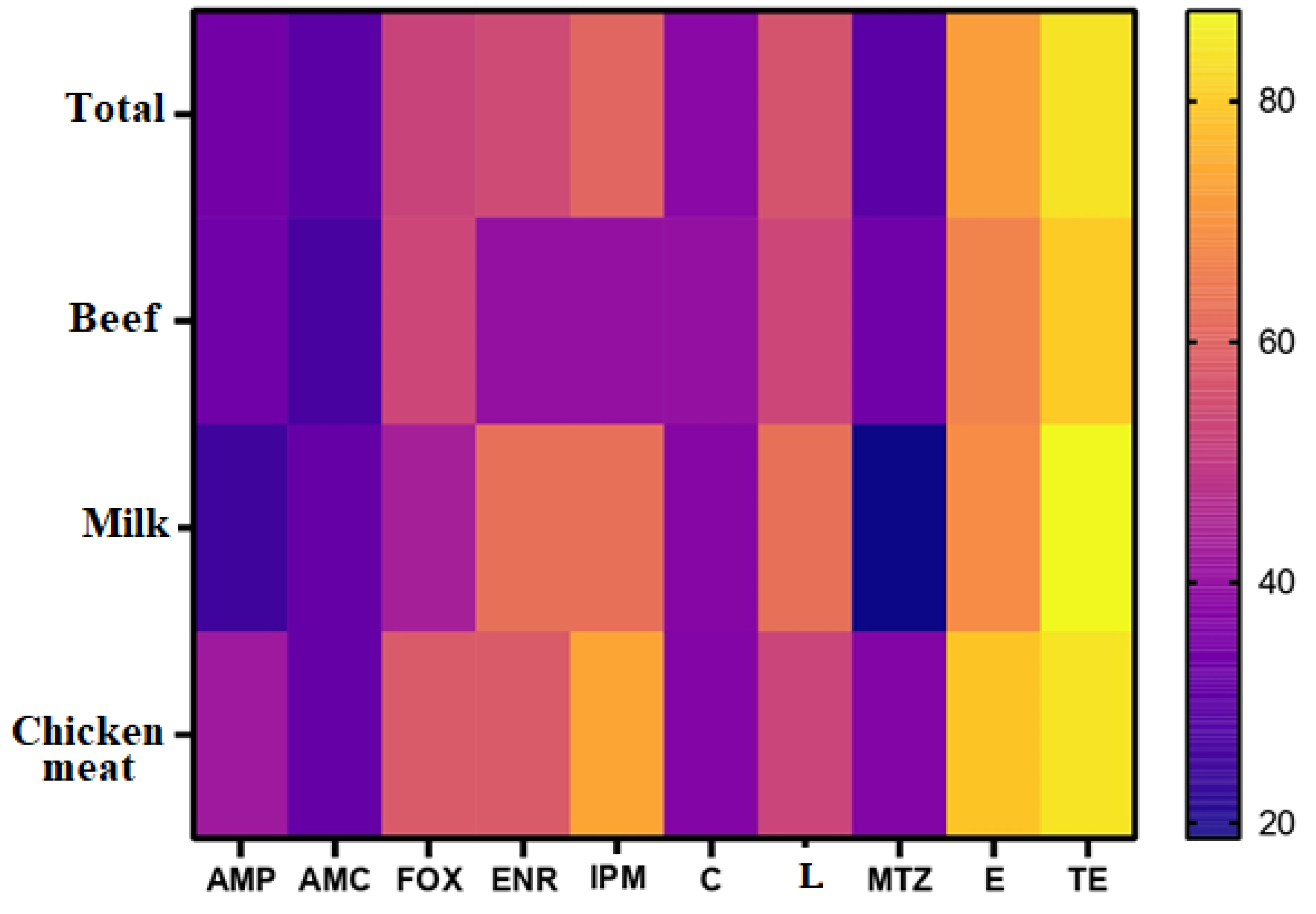
3.2. Antimicrobial Susceptibility Results
3.3. Molecular Basis of C. perfringens Resistance to Antimicrobials
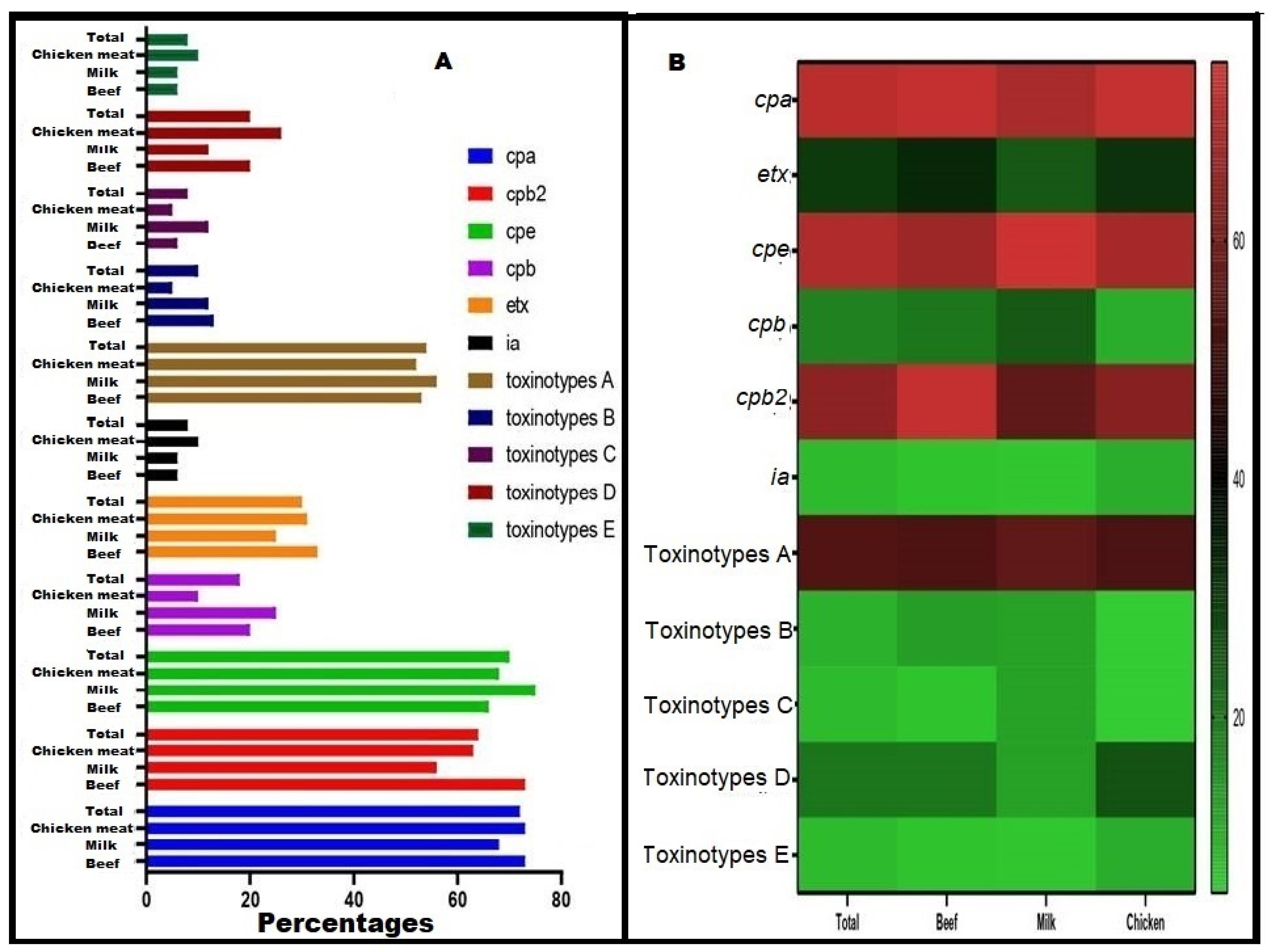
3.4. Toxinotyping and Toxin Gene Profiling of C. perfringens Isolates
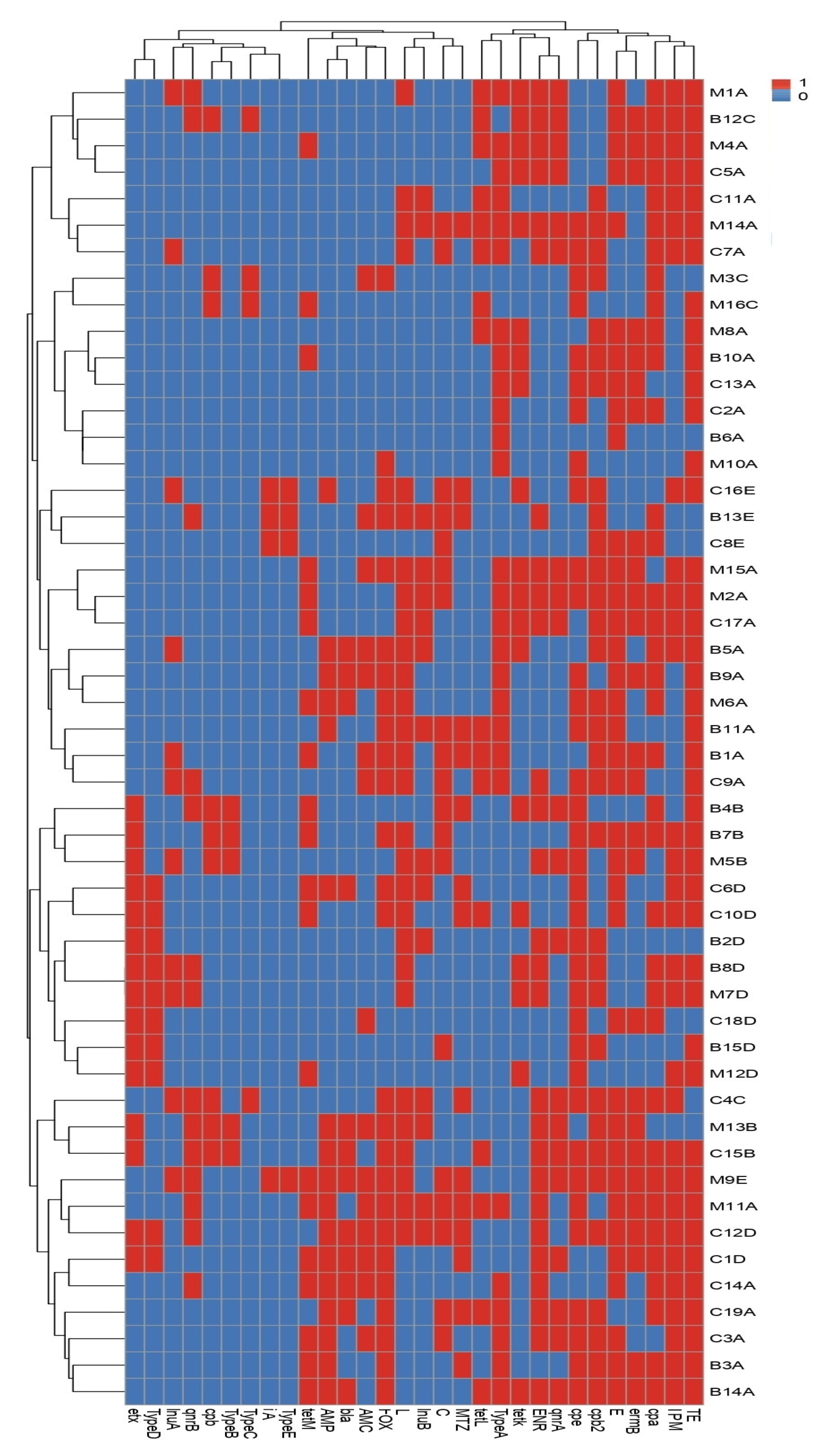
3.5. Phenotypic and Genotypic Diversity
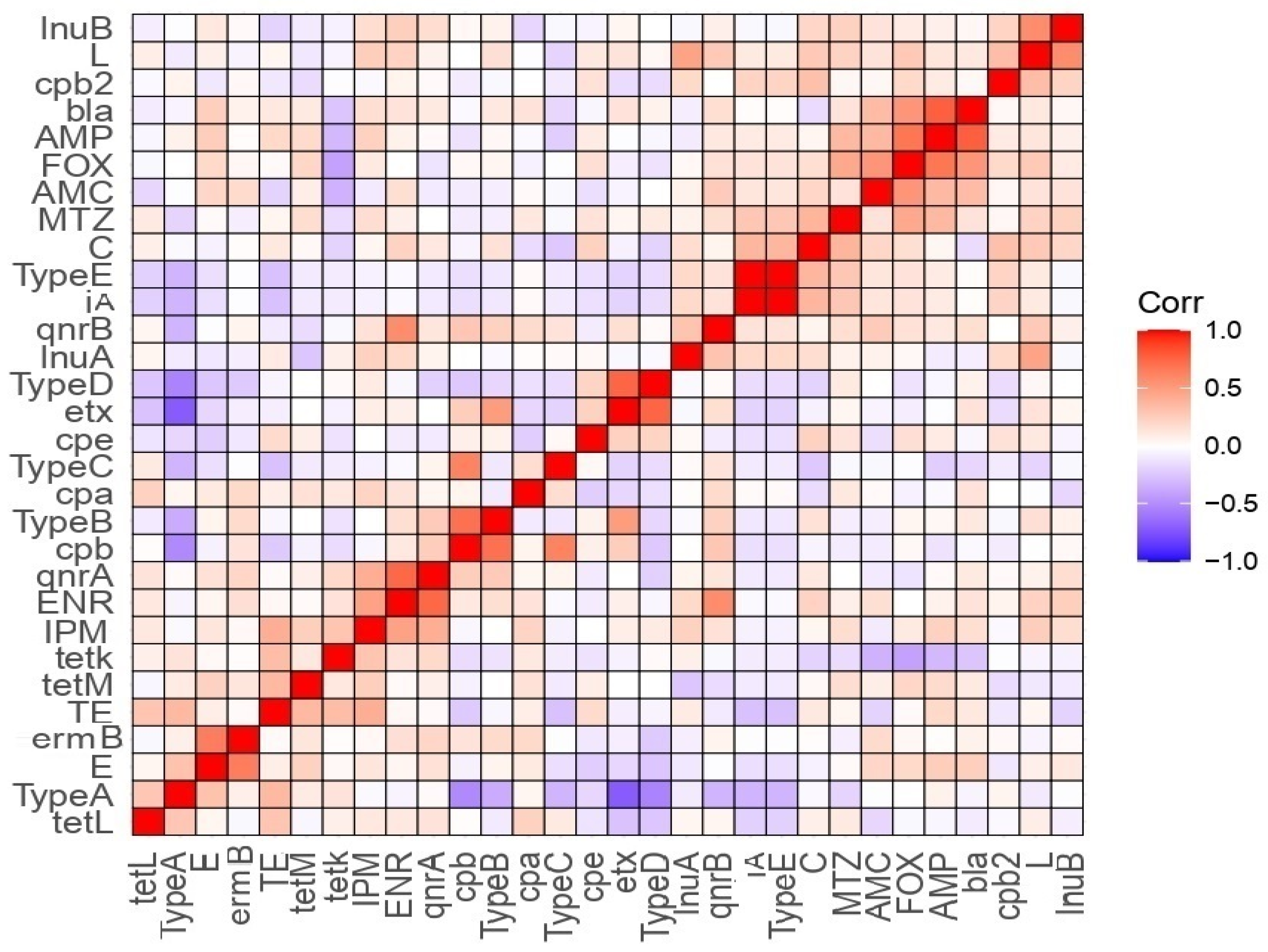
3.6. Correlation Analysis between Antimicrobial Susceptibility, Toxinotypes, and Toxin Gene Profiles
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ghaly, M.; Shaheen, A.; Bouhy, A.; Bendary, M. Alternative therapy to manage otitis media caused by multidrug-resistant fungi. Arch. Microbiol. 2020, 1, 1231–1240. [Google Scholar] [CrossRef] [PubMed]
- Okeke, I.N.; Laxminarayan, R.; Bhutta, Z.A.; Duse, A.G.; Jenkins, P.; O’Brien, T.F.; Pablos-Mendez, A.; Klugman, K.P. Antimicrobial resistance in developing countries. Part I: Recent trends and current status. Lancet Infect. Dis. 2005, 5, 481–493. [Google Scholar] [CrossRef]
- Abd El-Aziz, N.K.; Abd El-Hamid, M.I.; Bendary, M.M.; El-Azazy, A.A.; Ammar, A.M. Existence of vancomycin resistance among methicillin resistant S. aureues recovered from animal and human sources in Egypt. Slov. Vet. Res. 2018, 55, 221–230. [Google Scholar]
- Abd El-Hamid, M.I.; Abd El-Aziz, N.K.; Samir, M.; El-Naenaeey, E.Y.; Abo Remela, E.M.; Mosbah, R.A.; Bendary, M.M. Genetic Diversity of Campylobacter jejuni iolated from avian and human sources in Egypt. Front. Microbiol. 2019, 10, 2353. [Google Scholar] [CrossRef] [PubMed]
- Ghaly, M.F.; Nasr, Z.M.; Abousaty, A.I.; Seadawy, H.G.; Shaheen, M.A.A.; Albogami, S.; Al-Sanea, M.M.; Bendary, M.M. Alternative and complementary therapies against foodborne Salmonella infections. Antibiotics 2021, 10, 1453. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, D.; Abdelfattah-Hassan, A.; Badawi, M.; Ismail, T.A.; Bendary, M.M.; Abdelaziz, A.M.; Mosbah, R.A.; Mohamed, D.I.; Arisha, A.H.; Abd El-Hamid, M.I. Thymol nanoemulsion promoted broiler chicken’s growth, gastrointestinal barrier and bacterial community and conferred protection against Salmonella typhimurium. Sci. Rep. 2021, 11, 7742. [Google Scholar] [CrossRef] [PubMed]
- Gross, T.P.; Kamara, L.B.; Hatheway, C.L.; Powers, P.; Libonati, J.P.; Harmon, S.M.; Israel, E. Clostridium perfringens food poisoning: Use of serotyping in an outbreak setting. J. Clin. Microbiol. 1989, 27, 660–663. [Google Scholar] [CrossRef]
- Mellou, K.; Kyritsi, M.; Chrysostomou, A.; Sideroglou, T.; Georgakopoulou, T.; Hadjichristodoulou, C. Clostridium perfringens Foodborne Outbreak during an Athletic Event in Northern Greece, June 2019. Int. J. Environ. Res. Public Health 2019, 16, 3967. [Google Scholar] [CrossRef]
- Miller, R.W.; Skinner, J.; Sulakvelidze, A.; Mathis, G.F.; Hofacre, C.L. Bacteriophage therapy for control of necrotic enteritis of broiler chickens experimentally infected with Clostridium perfringens. Avian Dis. 2010, 54, 33–40. [Google Scholar] [CrossRef]
- Freedman, J.C.; Theoret, J.R.; Wisniewski, J.A.; Uzal, F.A.; Rood, J.I.; McClane, B.A. Clostridium perfringens type A-E toxin plasmids. Res. Microbiol. 2015, 166, 264–279. [Google Scholar] [CrossRef]
- Prescott, J.F. Brief Description of Animal Pathogenic Clostridia. In Clostridial Diseases of Animals, 1st ed.; Uzal, F.A., Songer, J.G., Prescott, J.F., Eds.; John Wiley and Sons: Ames, IA, USA, 2016; pp. 13–19. [Google Scholar]
- Adak, G.K.; Long, S.M.; O’Brien, S.J. Trends in indigenous foodborne disease and deaths, England and Wales: 1992 to 2000. Gut. 2002, 51, 832–841. [Google Scholar] [CrossRef] [PubMed]
- Leung, V.H.; Phan, Q.; Costa, C.E.; Nishimura, C.; Pung, K.; Horn, L.; Sosa, L. Notes from the Field: Clostridium perfringens outbreak at a catered lunch—Connecticut, September 2016. MMWR Morb. Morb. Mortal. Wkly. Rep. 2017, 66, 940–941. [Google Scholar] [CrossRef] [PubMed][Green Version]
- European Food Safety Authority (EFSA). European Centre for Disease Prevention and Control (ECDC) The European Union Summary Report on Trends and Sources of Zoonoses, Zoonotic Agents and Food-Borne Outbreaks in 2017. Available online: https://www.efsa.europa.eu/en/efsajournal/pub/5500 (accessed on 5 September 2019).
- Rhodehamel, J.; Harmon, S.M. Bacteriological Analytical Manual. In Clostridium Perfringens; U.S Food and Drug Adminstration, AOAC International: Gaithersburg, MD, USA, 1988; Chapter 16. Available online: https://www.fda.gov/food/laboratory-methods-food/bam-Clostridium-perfringens (accessed on 31 October 2017).
- Gizachew, H. A Review on Clostridium Perfringens Food Poisoning. Glob. Res. J. Public Health Epidemiol. 2017, 4, 104–109. [Google Scholar]
- Yoo, H.S.; Lee, S.U.; Park, K.Y.; Park, Y.H. Molecular typing and epidemiological survey of prevalence of Clostridium perfringens types by multiplex PCR. J. Clin. Microbiol. 1997, 35, 228–232. [Google Scholar] [CrossRef] [PubMed]
- Cooper, K.K.; Songer, J.G. Virulence of Clostridium perfringens in an experimental model of poultry necrotic enteritis. Vet. Microbiol. 2010, 142, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Rood, J.I.; McClane, B.A. Epsilon-toxin production by Clostridium perfringens type D strain CN3718 is dependent upon the agr operon but not the VirS/VirR two-component regulatory system. MBio 2011, 2, e00275-11. [Google Scholar] [CrossRef]
- Park, J.Y.; Kim, S.; Oh, J.Y.; Kim, H.R.; Jang, I.; Lee, H.S.; Kwon, Y.K. Characterization of Clostridium perfringens isolates obtained from 2010 to 2012 from chickens with necrotic enteritis in Korea. Poult. Sci. 2015, 94, 1158–1164. [Google Scholar] [CrossRef]
- Uzal, F.A.; Vidal, J.E.; McClane, B.A.; Gurjar, A.A. Clostridium perfringens Toxins Involved in Mammalian Veterinary Diseases. Open Toxinol. J. 2010, 2, 24–42. [Google Scholar] [CrossRef]
- Hall, B.G. Predicting the evolution of antibiotic resistance genes. Nat. Rev. Microbiol. 2004, 2, 430–435. [Google Scholar] [CrossRef]
- Ngamwongsatit, B.; Tanomsridachchai, W.; Suthienkul, O.; Urairong, S.; Navasakuljinda, W.; Janvilisri, T. Multidrug resistance in Clostridium perfringens isolated from diarrheal neonatal piglets in Thailand. Anaerobe 2016, 38, 88–93. [Google Scholar] [CrossRef]
- Ma, Y.H.; Ye, G.S. Determination of multidrug resistance mechanisms in Clostridium perfringens type A isolates using RNA sequencing and 2D-electrophoresis. Braz. J. Med. Biol. Res. 2018, 51, 8. [Google Scholar] [CrossRef] [PubMed]
- Bannam, T.L.; Johanesen, P.A.; Salvado, C.L.; Pidot, S.J.; Farrow, K.A.; Rood, J.I. The Clostridium perfringens TetA(P) efflux protein contains a functional variant of the Motif A region found in major facilitator superfamily transport proteins. Microbiology 2004, 150, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Martel, A.; Devriese, L.A.; Cauwerts, K.; De Gussem, K.; Decostere, A.; Haesebrouck, F. Susceptibility of Clostridium perfringens strains from broiler chickens to antibiotics and anticoccidials. Avian Pathol. 2004, 33, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.Z.; Islam, M.M. Characterization of β-lactamase and quinolone resistant Clostridium perfringens recovered from broiler chickens with necrotic enteritis in Bangladesh. Iran. J. Vet. Res. 2021, 22, 48–54. [Google Scholar]
- Soge, O.O.; Tivoli, L.D.; Meschke, J.S.; Roberts, M.C. A conjugativemacrolide resistance gene, mef(A), in environmental Clostridium perfringens carrying multiple macrolide and/or tetracyclineresistance genes. J. Appl. Microbiol. 2009, 106, 34–40. [Google Scholar] [CrossRef]
- ICMSF. Microorganisms in Foods. 1. Sampling Plans for Soft Drinks, Fruit Juices, Concentrates and Fruit Preserves, 2nd ed.; University of Toronto Press: Toronto, ON, Canada, 1978. [Google Scholar]
- Abudabos, A.M.; Alyemni, A.H.; Al-Marshad, M.B. Bacillus subtilis PB6 based-probiotic (CloSTAT TM) improves intestinal morphological and microbiological status of broiler chickens under Clostridium perfringens challenge. Int. J. Agric. Biol. 2013, 15, 978–982. [Google Scholar]
- Bergey, D.H. Clostridium. In Bergey’s Manual of Systematic Bacteriology, the Firmicutes, 2nd ed; Williams & Wilkins: Athens, GA, USA, 2009; Volume 3, p. 738. [Google Scholar]
- Kikuchi, E.; Miyamoto, Y.; Narushima, S.; Itoh, K. Design of species-specific primers to identify 13 species of Clostridium harbored in human intestinal tracts. Microbiol. Immunol. 2002, 46, 353–358. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute (CLSI). Methods for Antimicrobial Susceptibility Testing of Anaerobic Bacteria, 9th ed.; CLSI standard M11; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing, 30th ed.; CLSI supplement M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2020. [Google Scholar]
- Bullen, J.J. Clostridium welchii type D in the alimentary tract of normal sheep. J. Path Bact. 1952, 64, 201–210. [Google Scholar] [CrossRef]
- Sterne, M.; Batty, I. Pathogenic Clostridia; Butter Worth: London, UK; Boston, MA, USA, 1975. [Google Scholar]
- Smith, L.D.S.; Holdeman, L. The Pathogenic Anaerobic Bacteria, 1st ed.; Charles Thomas Publisher: Athens, GA, USA, 1968; pp. 201–255. [Google Scholar]
- Trzcinski, K.; Cooper, B.S.; Hryniewicz, W.; Dowson, C.G. Expression of resistance to tetracyclines in strains of methicillin-resistant Staphylococcus aureus. J. Antimicrob. Chemother. 2000, 45, 763–770. [Google Scholar] [CrossRef]
- Aminov, R.I.; Garrigues-Jeanjean, N.; Mackie, R.I. Molecular ecology of tetracycline resistance: Development and validation of primers for detection of tetracycline resistance genes encoding ribosomal protection proteins. Appl. Environ. Microbiol. 2001, 67, 22–32. [Google Scholar] [CrossRef]
- Masco, L.; Van-Hoorde, K.; De-Brandt, E.; Swings, J.; Huys, G. Antimicrobial susceptibility of Bifidobacterium strains from humans, animals and probiotic products. J. Antimicrob. Chemother. 2006, 58, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Cattoir, V.; Poirel, L.; Rotimi, V.; Soussy, C.J.; Nordmann, P. Multiplex PCR for detection of plasmid-mediated quinolone resistance qnr genes in ESBLproducing enterobacterial isolates. J. Antimicrob. Chemother. 2007, 60, 394–397. [Google Scholar] [CrossRef] [PubMed]
- Catalán, A.; Espoz, M.; Cortés, W.; Sagua, H.; González, J.; Araya, J. Tetracycline and penicillin resistant Clostridium perfringens isolated from the fangs and venom glands of Loxosceles laeta: Its implications in loxoscelism treatment. Toxicon 2010, 56, 890–896. [Google Scholar] [CrossRef] [PubMed]
- Meer, R.R.; Songer, J.G. Multiplex polymerase chain reaction assay for genotyping Clostridium perfringens. Am. J. Vet. Res. 1997, 58, 702–705. [Google Scholar]
- Bueschel, D.M.; Helen-Jost, B.; Billington, S.J.; Trinh, H.T.; Glenn-Songer, J. Prevalence of cpb2, encoding beta2 toxin, in Clostridium perfringens field isolates: Correlation of genotype with phenotype. Vet. Microbiol. 2003, 94, 121–129. [Google Scholar] [CrossRef]
- Ammar, A.M.; Attia, A.M.; Abd El-Aziz, N.K.; Abd El Hamid, M.I.; El-Demerdash, A.S. Class 1 integron and associated gene cassettes mediating multiple-drug resistance in some foodborne pathogens. Int. Food Res. J. 2016, 23, 332–339. [Google Scholar]
- Ammar, A.M.; Attia, A.M.; Abd El-Hamid, M.I.; El-Shorbagy, I.M.; Abd El-Kader, S.A. Genetic basis of resistance waves among methicillin resistant Staphylococcus aureus isolates recovered from milk and meat products in Egypt. Cell. Mol. Biol. 2016, 62, 7–15. [Google Scholar]
- Ahmed, H.A.; Tahoun, A.B.; Abou Elez, R.M.; Abd El-Hamid, M.I.; Abd Ellatif, S.S. Prevalence of Yersinia enterocolitica in milk and dairy products and the effects of storage temperatures on survival and virulence gene expression. Int. Dairy J. 2019, 94, 16–21. [Google Scholar] [CrossRef]
- Abd El-Hamid, M.I.; Bendary, M.M. Comparative phenotypic and genotypic discrimination of methicillin resistant and susceptible Staphylococcus aureus in Egypt. Cell. Mol. Biol. 2015, 61, 106–117. [Google Scholar]
- Abd El-Hamid, M.I.; Bendary, M.M.; Merwad, A.M.; Elsohaby, I.; Ghaith, D.M.; Alshareef, W.A. What is behind phylogenetic analysis of hospital-, communityand livestock-associated methicillin-resistant Staphylococcus aureus? Transbound Emerg. Dis. 2019, 66, 1506–1517. [Google Scholar]
- Ammar, A.M.; Abd El-Hamid, M.I.; El-Malt, R.S.; Azab, D.S.; Albogami, S.; Al-Sanea, M.M.; Soliman, W.E.; Ghoneim, M.M.; Bendary, M.M. Molecular detection of fluoroquinolone resistance among multidrug-, extensively drug-, and pan-drug-resistant Campylobacter species in Egypt. Antibiotics 2021, 10, 1342. [Google Scholar] [CrossRef] [PubMed]
- Mosallam, F.M.; Helmy, E.A.; Bendary, M.M.; El-Batal, I.A. Potency of a novel synthesized Ag- eugenol nanoemulsion for treating some bacterial and fungal pathogens. J. Mater. Res. 2021, 36, 1524–1537. [Google Scholar] [CrossRef]
- Elfaky, M.A.; Abdel-Hamid, M.I.; Khalifa, E.; Alshareef, W.A.; Mosbah, R.A.; Elazab, S.T.; Ghoneim, M.M.; Al-Sanea, M.M.; Bendary, M.M. Innovative next-generation therapies in combating multi-drug-resistant and multi-virulent Escherichia coli isolates: Insights from in vitro, in vivo, and molecular docking studies. Appl. Microbiol. Biotechnol. 2022, 106, 1691–1703. [Google Scholar] [CrossRef] [PubMed]
- McClane, B.A. Clostridium perfringens. In Food Microbiology: Fundamentals and Frontiers, 2nd ed; Beuchat, L.R., Doyle, M.P., Montville, T.J., Eds.; ASM Press: Washington, DC, USA, 2001; pp. 351–372. [Google Scholar]
- Kamber, U.; Gokce, H.I.; Elmali, M. Clostridium perfringens and its toxins in minced meat from Kars, Turkey. Food Addit. Contam. 2007, 24, 673–678. [Google Scholar] [CrossRef] [PubMed]
- Mead, P.S.; Slutsker, L.; Dietz, V.; McCaig, L.F.; Bresee, J.S.; Shapiro, C.; Griffen, P.M.; Tauxe, R.V. Food-related illness and death in the United States. Emerg. Infect. Dis. 1999, 5, 607–625. [Google Scholar] [CrossRef]
- Chalmers, G.; Bruce, H.L.; Hunter, D.B.; Parreira, V.R.; Kulkarni, R.R.; Jiang, Y.F.; Prescott, J.F.; Boerlin, P. Multilocus sequence typing analysis of Clostridium perfringens isolates from necrotic enteritis outbreaks in broiler chicken populations. J. Clin. Microbiol. 2008, 46, 3957–3964. [Google Scholar] [CrossRef]
- Gurmu, E.B.; Hazarika, R.A.; Borah, P.; Barua, A.G. Prevalence of enterotoxigenic Clostridium perfringens in foods of animal origin, Guwahati, India. J. Environ. Occup. Sci. 2013, 2, 45–50. [Google Scholar] [CrossRef]
- Emara, M.S. Anaerobic and Aerobic Microorganisms in Human Food. Master’s Thesis, Cairo University, Cairo, Egypt, 2014. [Google Scholar]
- Zhang, T.; Zhang, W.; Ai, D.; Zhang, R.; Lu, Q.; Luo, Q.; Shao, H. Prevalence and characterization of Clostridium perfringens in broiler chickens and retail chicken meat in central China. Anaerobe 2018, 54, 100–103. [Google Scholar] [CrossRef]
- Lin, Y.T.; Labbe, R. Enterotoxigenicity and genetic relatedness of Clostridium perfringens isolates from retail foods in the United States. Appl Environ. Microbiol. 2003, 69, 1642–1646. [Google Scholar] [CrossRef]
- Wen, Q.; McClane, B.A. Detection of enterotoxigenic Clostridium perfringens type A isolates in American retail foods. Appl. Environ. Microbiol. 2004, 70, 2685–2691. [Google Scholar] [CrossRef]
- Khan, M.; Nazir, J.; Anjum, A.A.; Ahmad, M.U.; Nawaz, M.; Shabbir, M.Z. Toxinotyping and antimicrobial susceptibility of enterotoxigenic Clostridium perfringens isolates from mutton, beef and chicken meat. J. Food Sci. Technol. 2015, 52, 5323–5328. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Miki, Y.; Miyamoto, K.; Kaneko-Hirano, I.; Fujiuchi, K.; Akimoto, S. Prevalence and characterization of enterotoxin gene-carrying Clostridium perfringens isolates from retail meat products in Japan. Appl. Environ. Microbiol. 2008, 74, 5366–5372. [Google Scholar] [CrossRef] [PubMed]
- Hashem, H.M. Bacteriological criteria of dressed poultry with special reference to some microbial decontaminators. Master’s Thesis, Benha University, Cairo, Egypt, 2015. [Google Scholar]
- Hemmat, M.I.; Amin, R.A.; El-Shater, M.A.; Hafez, M.S. Bacteriological evaluation of freshly slaughtered chicken carcasses. BVMJ 2015, 28, 74–82. [Google Scholar]
- Miwa, N.; Nishina, T.; Kubo, S.; Atsumi, M.; Honda, H. Amount of enterotoxigenic Clostridium perfringens in meat detected by nested PCR. Int. J. Food Microbiol. 1998, 42, 195–200. [Google Scholar] [CrossRef]
- Singh, R.V.; Bhilegaonkar, K.N.; Agarwal, R.K. Studies on occurrence and characterization of Clostridium perfringens from select meats. J. Food Saf. 2005, 25, 146–156. [Google Scholar] [CrossRef]
- Kamal, A. Clostridium perfringens in Meat and Chicken Received in University Hostel. Master’s Thesis, Benha University, Cairo, Egypt, 2017. [Google Scholar]
- Arnold, S.; Gassner, B.; Giger, T.; Zwahlen, R. Banning antimicrobial growth promoters in feedstuffs does not result in increased therapeutic use of antibiotics in medicated feed in pig farming. Pharmacoepidemiol. Drug Saf. 2004, 13, 323–331. [Google Scholar] [CrossRef]
- Marks, S.L.; Kather, E.J. Antimicrobial susceptibilities of canine Clostridium difficile and Clostridium perfringens isolates to commonly utilized antimicrobial drugs. Vet. Microbiol. 2003, 94, 39–45. [Google Scholar] [CrossRef]
- Chon, J.W.; Seo, K.H.; Bae, D.; Park, J.H.; Khan, S.; Sung, K. Prevalence, toxin gene profile, antibiotic resistance, and molecular characterization of Clostridium perfringens from diarrheic and non-diarrheic dogs in Korea. J. Vet. Sci. 2018, 19, 368–374. [Google Scholar] [CrossRef]
- Slavić, D.; Boerlin, P.; Fabri, M.; Klotins, K.C.; Zoethout, J.K.; Weir, P.E.; Bateman, D. Antimicrobial susceptibility of Clostridium perfringens isolates of bovine, chicken, porcine, and turkey origin from Ontario. Can. J. Vet. Res. Rev. Can. Rech. Vet. 2011, 75, 89–97. [Google Scholar]
- Ghoneim, N.H.; Hamza, D.A. Epidemiological studies on Clostridium perfringens food poisoning in retail foods. Rev. Sci. Tech. Int. Off. Epizoot. 2017, 36, 1025–1032. [Google Scholar] [CrossRef]
- Nauerby, B.; Pedersen, K.; Madsen, M. Analysis by pulsed-field gel electrophoresis of the genetic diversity among Clostridium perfringens isolates from chickens. Vet. Microbiol. 2003, 94, 257–266. [Google Scholar] [CrossRef]
- Gholamiandekhordi, A.R.; Ducatelle, R.; Heyndrickx, M.; Haesebrouck, F.; Van-Immerseel, F. Molecular and phenotypical characterization of Clostridium perfringens isolates from poultry flocks with different disease status. Vet. Microbiol. 2006, 113, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, H.; Ogasawara, J.; Monma, C.; Hase, A.; Suzuki, H.; Kai, A.; Haruki, K.; Nishikawa, Y. Usefulness of a combination of pulsed-field gel electrophoresis and enrichment culture in labora-tory investigation of a foodborne outbreak due to Clostridium perfringens. Diagn. Microbiol. Infect. Dis. 2003, 47, 471–475. [Google Scholar] [CrossRef]
- Fisher, D.J.; Miyamoto, K.; Harrison, B.; Akimoto, S.; Sarker, M.R.; McClane, B.A. association of beta2 toxin production with Clostridium perfringens type A human gastrointestinal disease isolates carrying a plasmid enterotoxin gene. Mol. Microbiol. 2005, 56, 747–762. [Google Scholar] [CrossRef]
- Xiao, Y.; Wagendorp, A.; Moezelaar, R.; Abee, T.; Wells-Bennik, M.H. A wide variety of Clostridium perfringens type A food-borne isolates that carry a chromosomal cpe gene belong to one multilocus sequence typing cluster. Appl. Environ. Microbiol. 2012, 78, 7060–7068. [Google Scholar] [CrossRef]
- Lacey, J.A.; Keyburn, A.L.; Ford, M.E.; Portela, R.W.; Johanesen, P.A.; Lyras, D.; Moore, R.J. Conjugation-mediated horizontal gene transfer of Clostridium perfringens plasmids in the chicken gastrointestinal tract results in the formation of new virulent strains. Appl. Environ. Microbiol. 2017, 83, e01814-17. [Google Scholar] [CrossRef] [PubMed]
- Petit, L.; Gibert, M.; Popoff, M.R. Clostridium perfringens: Toxinotype and genotype. Trends Microbiol. 1999, 7, 104–110. [Google Scholar] [CrossRef]
- Rood, J.I.; Adams, V.; Lacey, J.; Lyras, D.; McClane, B.A.; Melville, S.B.; Moore, R.J.; Popoff, M.R.; Sarker, M.R.; Songer, J.G. Expansion of the Clostridium perfringens toxin-based typing scheme. Anaerobes 2018, 53, 5–10. [Google Scholar] [CrossRef]
- Doyle, M.E. Multidrug-resistant pathogens in the food supply. Foodborne Pathog. Dis. 2015, 12, 261–279. [Google Scholar] [CrossRef]
- Fayez, M.; El-Ghareeb, W.R.; Elmoslemany, A.; Alsunaini, S.J.; Alkafafy, M.; Alzahrani, O.M.; Mahmoud, S.F.; Elsohaby, I. genotyping and antimicrobial susceptibility of Clostridium perfringens and clostridioides difficile in camel minced meat. Pathogens 2021, 10, 1640. [Google Scholar] [CrossRef]
- Beceiro, A.; Tomás, M.; Bou, G. Antimicrobial resistance and virulence: A successful or deleterious association in the bacterial world? Clin. Microbiol. Rev. 2013, 26, 185–230. [Google Scholar] [CrossRef] [PubMed]
- Vilei, E.M.; Schlatter, Y.; Perreten, V.; Straub, R.; Popoff, M.R.; Gibert, M.; Grone, A.; Frey, J. Antibiotic-induced expression of a cryptic cpb2 gene in equine β2-toxigenic Clostridium perfringens. Mol. Microbiol. 2005, 57, 1570–1581. [Google Scholar] [CrossRef] [PubMed]
- Ahn, D.; Prince, A. Host-pathogen interface: Progress in understanding the pathogenesis of infection due to multidrug-resistant bacteria in the intensive care unit. J. Infect. Dis. 2017, 215, S1–S8. [Google Scholar] [CrossRef] [PubMed]
- Wei, B.; Cha, S.Y.; Zhang, J.F.; Shang, K.; Park, H.C.; Kang, J.; Lee, K.J.; Kang, M.; Jang, H.K. Antimicrobial susceptibility and association with toxin determinants in Clostridium perfringens isolates from chickens. Microorganisms 2020, 8, 1825. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primer Sequence (5′-3′) | Amplicon Size (bp) | Annealing Temperature (°C) | Reference |
|---|---|---|---|---|
| tet(K) | F: TTATGGTGGTTGTAGCTAGAAA R: AAAGGGTTAGAAACTCTTGAAA | 382 | 50 | [40] |
| tet(L) | F: ATAAATTGTTTCGGGTCGGTAAT R: AACCAGCCAACTAATGACAATGAT | 1077 | 50 | [38] |
| tet(M) | F: ACAGAAAGCTTATTATATAAC R: TGGCGTGTCTATGATGTTCAC | 171 | 55 | [39] |
| lnu(A) | F: GGTGGCTGGGGGGTAGATGTATTAACTGG R: GCTTCTTTTGAAATACATGGTATTTTTCGATC | 323 | 54 | [39] |
| lnu(B) | F: CCTACCTATTGTTTGTGGAA R: ATAACGTTACTCTCCTATTC | 906 | 45 | [39] |
| erm(B) | F: GAAAAGGTACTCAACCAAATA R: AGTAACGGTACTTAAATTGTTTAC | 638 | 57 | [28] |
| bla | F: ATGAAAGAAGTTCAAAAATATTTAGAG R: TTAGTGCCAATTGTTCATGATGG | 780 | 50 | [42] |
| qnrA | F: AGAGGATTTCTCACGCCAGG R: TGCCAGGCACAGATCTTGAC | 580 | 54 | [41] |
| qnrB | F: GGMATHGAAATTCGCCACTG R: TTTGCYGYYCGCCAGTCGAA | 264 | 54 | [41] |
| cpa | F: GCTAATGTTACTGCCGTTGA R: CCTCTGATACATCGTGTAAG | 324 | 54 | [41] |
| cpb | F: GCGAATATGCTGAATCATCTA R: GCAGGAACATTAGTATATCTTC | 196 | 54 | [43] |
| etx | F: GCGGTGATATCCATCTATTC R: CCACTTACTTGTCCTACTAAC | 655 | 54 | [43] |
| iA | F: ACTACTCTCAGACAAGACAG R: CTTTCCTTCTATTACTATACG | 446 | 54 | [43] |
| cpe | F: GGAGATGGTTGGATATTAGG R: GGACCAGCAGTTGTAGATA | 233 | 54 | [43] |
| cpb2 | F: AGATTTTAAATATGATCCTAACC R: CAATACCCTTCACCAAATACTC | 567 | 54 | [44] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bendary, M.M.; Abd El-Hamid, M.I.; El-Tarabili, R.M.; Hefny, A.A.; Algendy, R.M.; Elzohairy, N.A.; Ghoneim, M.M.; Al-Sanea, M.M.; Nahari, M.H.; Moustafa, W.H. Clostridium perfringens Associated with Foodborne Infections of Animal Origins: Insights into Prevalence, Antimicrobial Resistance, Toxin Genes Profiles, and Toxinotypes. Biology 2022, 11, 551. https://doi.org/10.3390/biology11040551
Bendary MM, Abd El-Hamid MI, El-Tarabili RM, Hefny AA, Algendy RM, Elzohairy NA, Ghoneim MM, Al-Sanea MM, Nahari MH, Moustafa WH. Clostridium perfringens Associated with Foodborne Infections of Animal Origins: Insights into Prevalence, Antimicrobial Resistance, Toxin Genes Profiles, and Toxinotypes. Biology. 2022; 11(4):551. https://doi.org/10.3390/biology11040551
Chicago/Turabian StyleBendary, Mahmoud M., Marwa I. Abd El-Hamid, Reham M. El-Tarabili, Ahmed A. Hefny, Reem M. Algendy, Nahla A. Elzohairy, Mohammed M. Ghoneim, Mohammad M. Al-Sanea, Mohammed H. Nahari, and Walaa H. Moustafa. 2022. "Clostridium perfringens Associated with Foodborne Infections of Animal Origins: Insights into Prevalence, Antimicrobial Resistance, Toxin Genes Profiles, and Toxinotypes" Biology 11, no. 4: 551. https://doi.org/10.3390/biology11040551
APA StyleBendary, M. M., Abd El-Hamid, M. I., El-Tarabili, R. M., Hefny, A. A., Algendy, R. M., Elzohairy, N. A., Ghoneim, M. M., Al-Sanea, M. M., Nahari, M. H., & Moustafa, W. H. (2022). Clostridium perfringens Associated with Foodborne Infections of Animal Origins: Insights into Prevalence, Antimicrobial Resistance, Toxin Genes Profiles, and Toxinotypes. Biology, 11(4), 551. https://doi.org/10.3390/biology11040551