
Identification of Key Candidate Genes in Runs of Homozygosity of the Genome of Two Chicken Breeds, Associated with Cold Adaptation



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
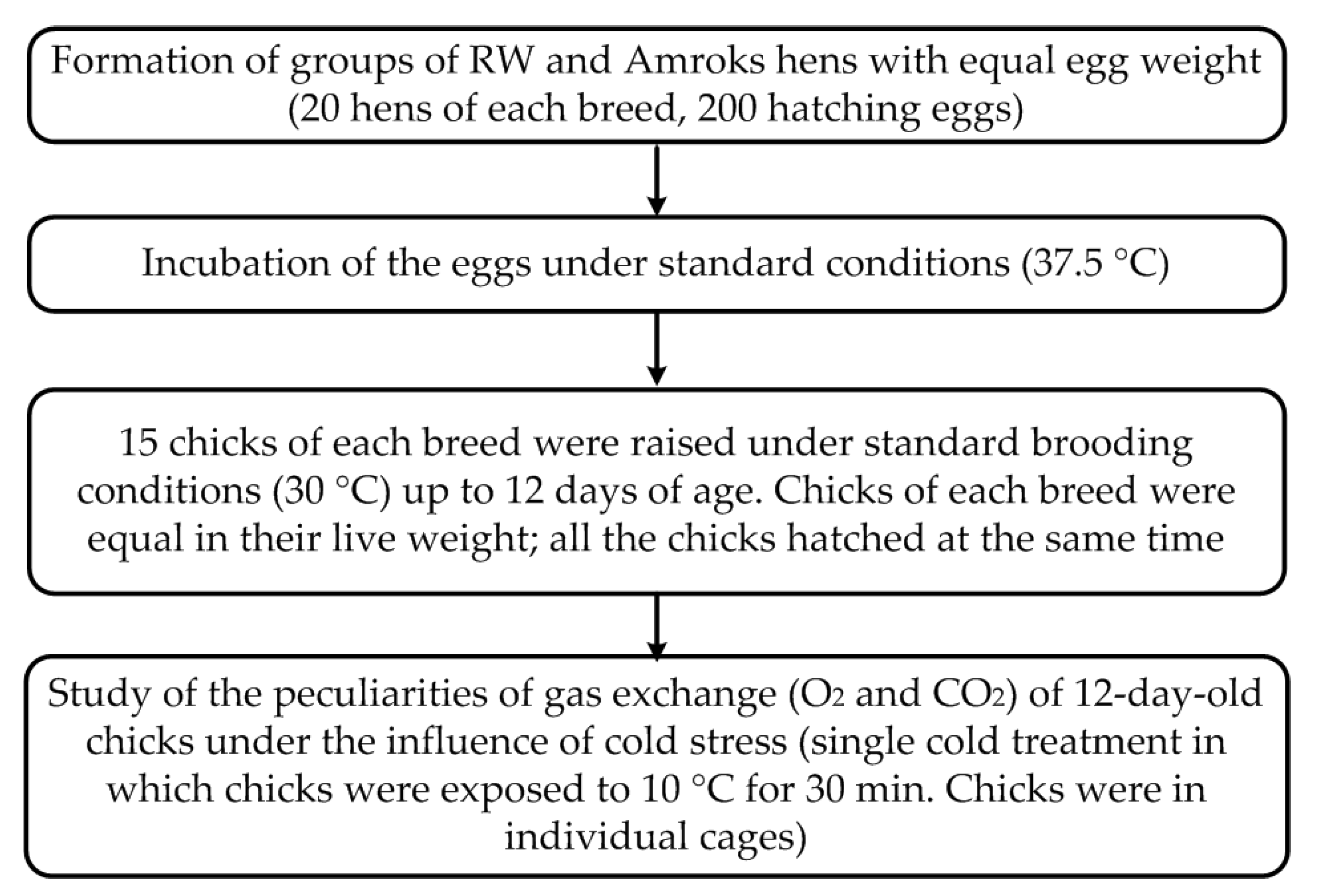
2.1. Experimental Chicken Breeds
2.2. Experimental Design of Thermoregulation Capabilities
2.3. Molecular Genetic Research
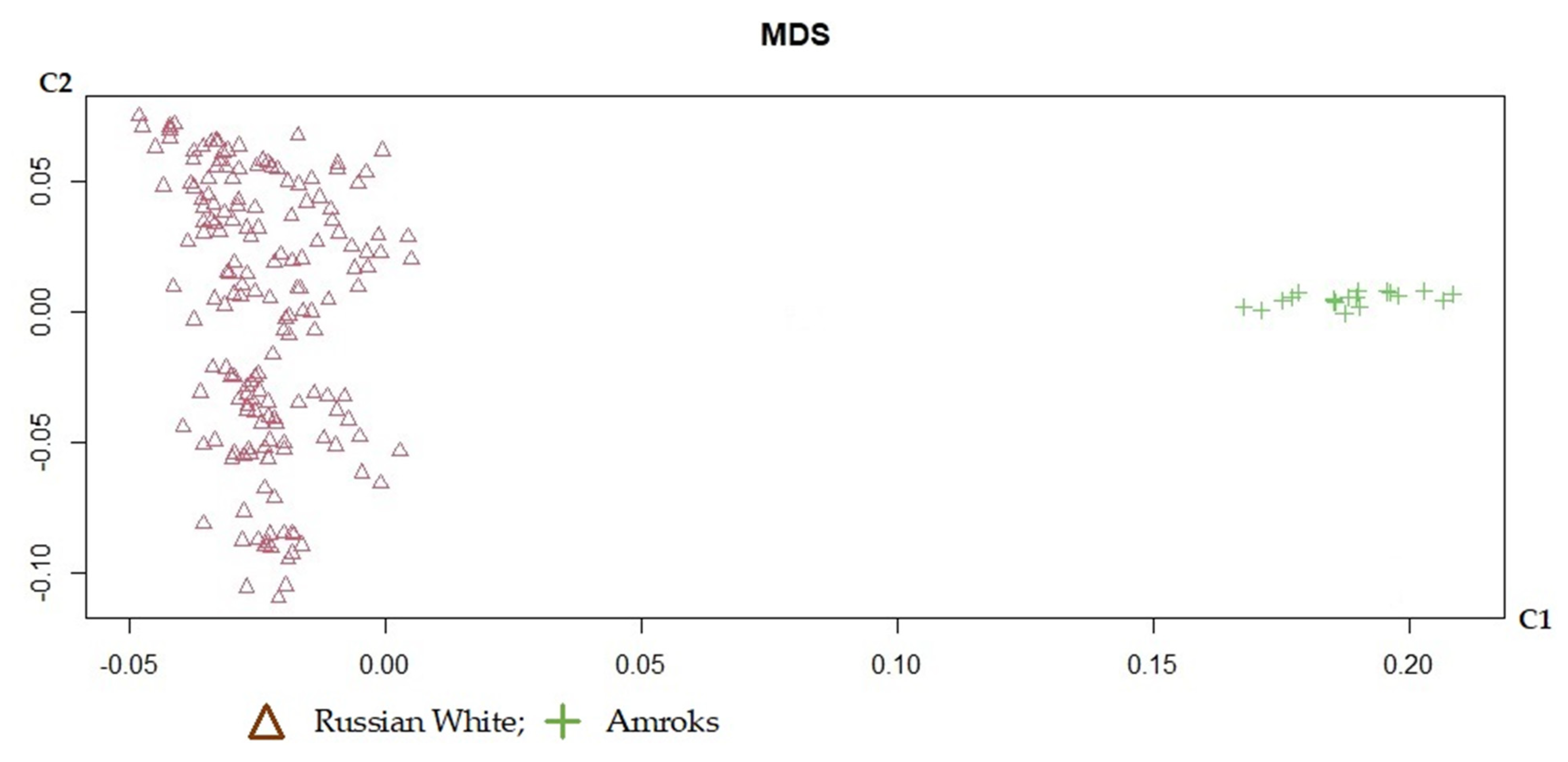
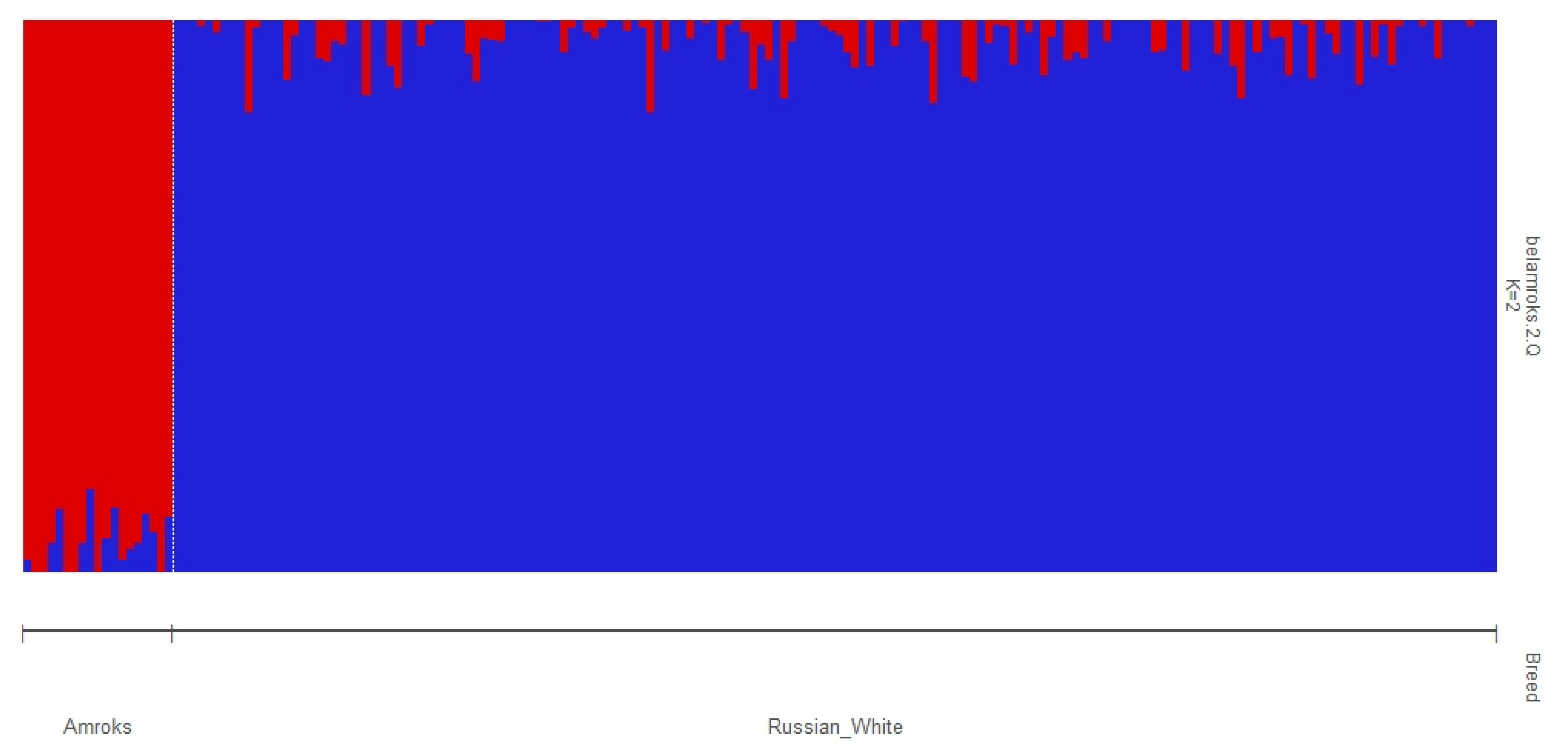
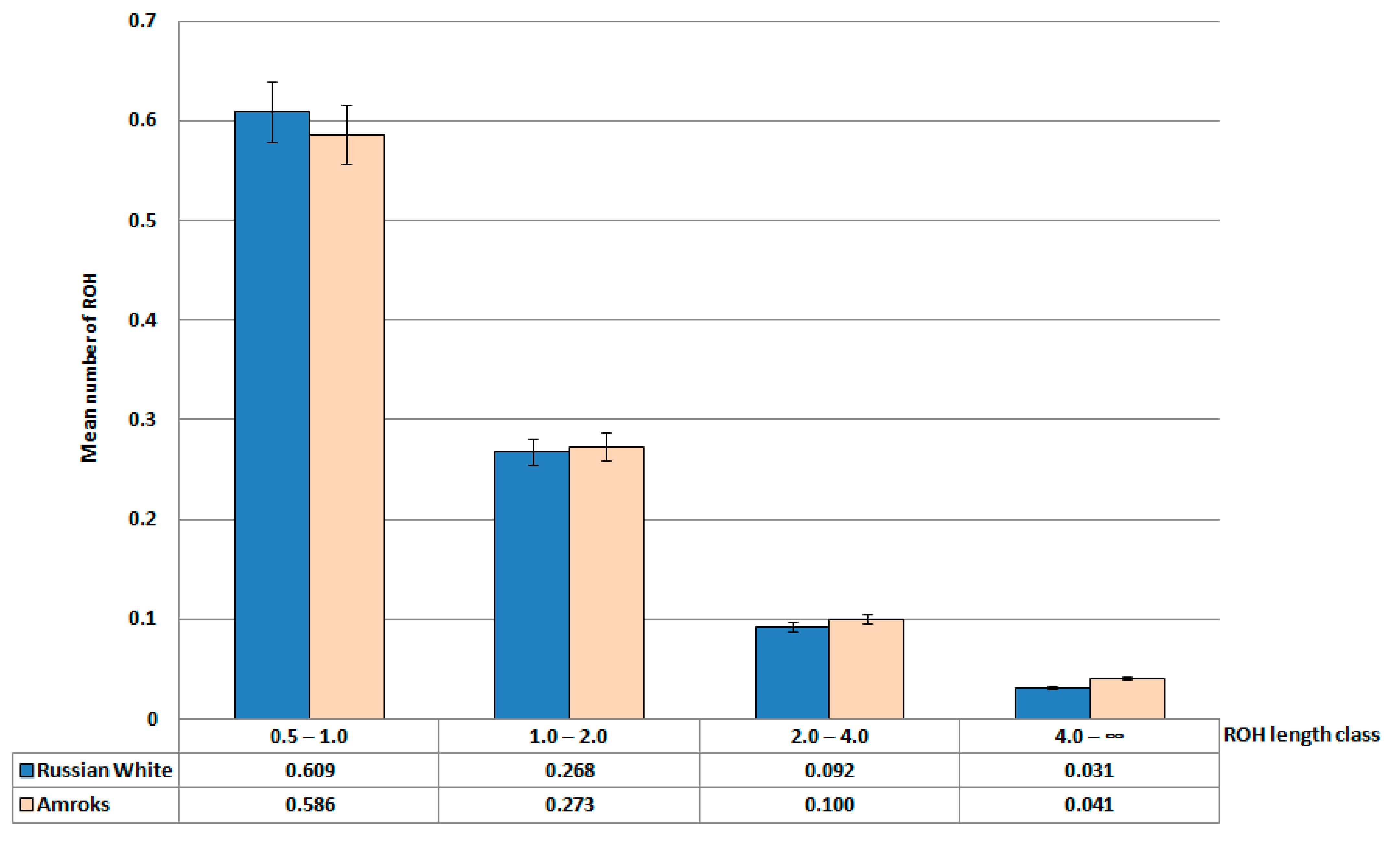
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Muir, W.M.; Wong, G.K.S.; Zhang, Y.; Wang, J.; Groenen, M.A.M.; Crooijmans, R.P.M.A.; Megens, H.J.; Zhang, H.; Okimoto, R.; Vereijken, A.; et al. Genome-wide assessment of worldwide chicken SNP genetic diversity indicates significant absence of rare alleles in commercial breeds. Proc. Natl. Acad. Sci. USA 2008, 105, 17312–17317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goel, A.; Ncho, C.M.; Choi, Y.H. Regulation of gene expression in chickens by heat stress. J. Anim. Sci. Biotechnol. 2021, 12, 11. [Google Scholar] [CrossRef] [PubMed]
- Cedraz de Oliveira, H.; Pinto Garcia, A.A., Jr.; Gonzaga Gromboni, J.G.; Vasconcelos Farias Filho, R.; Souza do Nascimento, C.; Arias Wenceslau, A. Influence of heat stress, sex and genetic groups on reference genes stability in muscle tissue of chicken. PLoS ONE 2017, 12, e0176402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radwan, L.M. Genetic improvement of egg laying traits in Fayoumi chickens bred under conditions of heat stress through selection and gene expression studies. J. Therm. Biol. 2020, 89, 102546. [Google Scholar] [CrossRef] [PubMed]
- Adu-Asiamah, P.; Zhang, Y.; Amoah, K.; Leng, Q.Y.; Zheng, J.H.; Yang, H.; Zhang, W.L.; Zhang, L. Evaluation of physiological and molecular responses to acute heat stress in two chicken breeds. Animal 2021, 15, 100106. [Google Scholar] [CrossRef] [PubMed]
- Galal, A.; Radwan, L.M.; Rezik, H.H.; Ayoub, H. Expression levels of HSP70 and CPT-1 in three local breeds of chickens reared under normal or heat stress conditions after the introduction of the naked neck gene. J. Therm. Biol. 2019, 80, 113–118. [Google Scholar] [CrossRef]
- Al-Zghoul, M.B.; Saleh, K.M.; Ababneh, M.M.K. Effects of pre-hatch thermal manipulation and post-hatch acute heat stress on the mRNA expression of interleukin-6 and genes involved in its induction pathways in 2 broiler chicken breeds. Poult. Sci. 2019, 98, 1805–1819. [Google Scholar] [CrossRef]
- Irivboje, Y.I.; Sanni, M.T.; Fafiolu, A.O.; Olowofeso, O.; Ikeobi, C.O.N. Genetic polymorphisms in part of intron 7 and exon 8 of HSP90AA1 gene and its association with heat tolerance traits in two exotic layer chicken strains. Trop. Anim. Health Prod. 2020, 52, 969–977. [Google Scholar] [CrossRef]
- Xu, N.Y.; Si, W.; Li, M.; Gong, M.; Larivière, J.M.; Nanaei, H.A.; Bian, P.P.; Jiang, Y.; Zhao, X. Genome-wide scan for selective footprints and genes related to cold tolerance in Chantecler chickens. Zool. Res. 2021, 42, 710–720. [Google Scholar] [CrossRef]
- Xie, S.; Yang, X.; Wang, D.; Zhu, F.; Yang, N.; Hou, Z.; Ning, Z. Thyroid transcriptome analysis reveals different adaptive responses to cold environmental conditions between two chicken breeds. PLoS ONE 2018, 13, e0191096. [Google Scholar] [CrossRef] [Green Version]
- Xie, S.; Yang, X.; Gao, Y.; Jiao, W.; Li, X.; Li, Y.; Ning, Z. Performance differences of Rhode Island Red, Bashang Long-tail Chicken, and their reciprocal crossbreds under natural cold stress. Asian-Australas. J. Anim. Sci. 2017, 30, 1507–1514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dementieva, N.V.; Kudinov, A.A.; Larkina, T.A.; Mitrofanova, O.V.; Dysin, A.P.; Terletsky, V.P.; Tyshchenko, V.I.; Griffin, D.K.; Romanov, M.N. Genetic variability in local and imported germplasm chicken populations as revealed by analyzing runs of homozygosity. Animals 2020, 10, 1887. [Google Scholar] [CrossRef] [PubMed]
- Sokolova, A.N. Genetiko-Selekcionny‘e Metody‘ Sozdaniya Populyacii kur s Povy‘Shennoj Ustojchivost‘yu k Neoplazmam. Ph.D. Thesis, All-Russian Research Institute for Genetics and Farm Animal Breeding, St. Petersburg, Russia, January 1999. [Google Scholar]
- Sokolova, A.N. Selekciya kur po funkcii termoregulyacii i produktivnosti. In Proceedings of the 13th World’s Poultry Science Congress, USSR, Kiev, Ukraine, 15–21 August 1966. [Google Scholar]
- Dmitriev, V.B. Funkcional‘ny‘e e‘Ndokrinny‘e Rezervy‘ v Selekcii sel‘Skoxozyajstvenny‘x Zhivotny‘x; Savozh: St. Petersburg, Russia, 2009; p. 244. [Google Scholar]
- Dement’ev, G.P. Issledovaniya po okraske pozvonochnyh zhivotnyh. Zool. Zhurnal 1948, 23, 189–197. [Google Scholar]
- Rol’nik, V.V. Biologiya Embrional’nogo Razvitiya Ptic; Nauka: Leningrad, Russia, 1968; p. 107. [Google Scholar]
- Stanishevskaya, O.I.; Fedorova, E.S. Comparative evaluation of the peculiarities of stress reactivity of the Russian White breed chicken with sw+ mutation and Amrox in hypothermia conditions during embryonal and early postnatal periods of ontogenesis. Agric. Biol. 2019, 54, 1135–1143. [Google Scholar] [CrossRef]
- Stanishevskaya, O.I.; Fedorova, E.S. Dosed exposure to low temperature as a breeding background in the selection of gene pool breeds of chickens for viral vaccines production. Open Agric. J. 2020, 14, 345–351. [Google Scholar] [CrossRef]
- Abdelmanova, A.S.; Dotsev, A.V.; Romanov, M.N.; Stanishevskaya, O.I.; Gladyr, E.A.; Rodionov, A.N.; Vetokh, A.N.; Volkova, N.A.; Fedorova, E.S.; Gusev, I.V.; et al. Unveiling Comparative Genomic Trajectories of Selection and Key Candidate Genes in Egg-Type Russian White and Meat-Type White Cornish Chickens. Biology 2021, 10, 876. [Google Scholar] [CrossRef] [PubMed]
- Dementieva, N.V.; Mitrofanova, O.V.; Dysin, A.P.; Kudinov, A.A.; Stanishevskaya, O.I.; Larkina, T.A.; Plemyashov, K.V.; Griffin, D.K.; Romanov, M.N.; Smaragdov, M.G. Assessing the effects of rare alleles and linkage disequilibrium on estimates of genetic diversity in the chicken populations. Animal 2021, 15, 100171. [Google Scholar] [CrossRef]
- Kudinov, A.A.; Dementieva, N.V.; Mitrofanova, O.V.; Stanishevskaya, O.I.; Fedorova, E.S.; Larkina, T.A.; Mishina, A.I.; Plemyashov, K.V.; Griffin, D.K.; Romanov, M.N. Genome-wide association studies targeting the yield of extraembryonic fluid and production traits in Russian White chickens. BMC Genom. 2019, 20, 270. [Google Scholar] [CrossRef]
- Schmidt, H.; Proll, R. Taschenatlas Hühner und Zwerghühner: 182 Rassen für Garten, Haus, Hof und Ausstellung, 2nd ed.; Ulmer: Schduegerd, Germany, 2008; 191 S. [Google Scholar]
- Walsberg, G.; Wolf, B. Variation in the respiratory quotient of birds and implications for indirect calorimetry using measurements of carbon dioxide production. J. Exp. Biol. 1995, 198 Pt 1, 213–219. [Google Scholar] [CrossRef]
- Chang, C.C.; Chow, C.C.; Tellier, L.C.; Vattikuti, S.; Purcell, S.M.; Lee, J.J. Second-generation PLINK: Rising to the challenge of larger and richer datasets. Gigascience 2015, 4, 7. [Google Scholar] [CrossRef]
- Thomas, L.; Ferreira, M.A.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.; Daly, M.J.; Sham, P.C. PLINK: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [Green Version]
- Alexander, D.H.; Novembre, J.; Lange, K. Fast model-based estimation of ancestry in unrelated individuals. Genome Res. 2009, 19, 1655–1664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, S. Variability within and among Natural Populations. In Evolution and the Genetics of Populations; University of Chicago Press: Chicago, IL, USA, 1978; p. 590. [Google Scholar]
- Patterson, N.; Price, A.L.; Reich, D. Population structure and eigenanalysis. PLoS Genet. 2006, 2, e190. [Google Scholar] [CrossRef] [PubMed]
- Biscarini, F.; Paolo Cozzi, P.; Gaspa, G.; Marras, G. detectRUNS: Detect Runs of Homozygosity and Runs of Heterozygosity in Diploid Genomes. R Package Version 0.9.5. Available online: https://cran.r-project.org (accessed on 7 February 2022).
- Jin, C.F.; Chen, Y.J.; Yang, Z.Q.; Shi, K.; Chen, C.K. A genome-wide association study of growth trait-related single nucleotide polymorphisms in Chinese Yancheng chickens. Genet. Mol. Res. 2015, 14, 15783–15792. [Google Scholar] [CrossRef]
- Shen, M.; Qu, L.; Ma, M.; Dou, T.; Lu, J.; Guo, J.; Hu, Y.; Wang, X.; Li, Y.; Wang, K.; et al. A genome-wide study to identify genes responsible for oviduct development in chickens. PLoS ONE 2017, 12, e0189955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, S.; Chen, Z.T.; Zheng, R.; Diao, S.; Teng, J.; Yuan, X.; Zhang, H.; Chen, Z.; Zhang, X.; Li, J.; et al. New Insights From Imputed Whole-Genome Sequence-Based Genome-Wide Association Analysis and Transcriptome Analysis: The Genetic Mechanisms Underlying Residual Feed Intake in Chickens. Front. Genet. 2020, 11, 243. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Liu, R.; Zhao, G.; Zheng, M.; Li, P.; Liu, L.; Wen, J. Genome-Wide Linkage Analysis Identifies Loci for Testicle and Ovary Traits in Chickens. Anim. Biotechnol. 2018, 29, 309–315. [Google Scholar] [CrossRef]
- Van Goor, A.; Bolek, K.J.; Ashwell, C.M.; Persia, M.E.; Rothschild, M.F.; Schmidt, C.J.; Lamont, S.J. Identification of quantitative trait loci for body temperature, body weight, breast yield, and digestibility in an advanced intercross line of chickens under heat stress. Genet. Sel. Evol. 2015, 47, 96. [Google Scholar] [CrossRef] [Green Version]
- Romé, H.; Varenne, A.; Hérault, F.; Chapuis, H.; Alleno, C.; Dehais, P.; Vignal, A.; Burlot, T.; Le Roy, P. GWAS analyses reveal QTL in egg layers that differ in response to diet differences. Genet. Sel. Evol. 2015, 47, 83. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Li, D.; Liu, J.; Chen, S.; Qu, L.; Zheng, J.; Xu, G.; Yang, N. A genome-wide SNP scan reveals novel loci for egg production and quality traits in white leghorn and brown-egg dwarf layers. PLoS ONE 2011, 6, e28600. [Google Scholar] [CrossRef] [Green Version]
- Mignon-Grasteau, S.; Rideau, N.; Gabriel, I.; Chantry-Darmon, C.; Boscher, M.Y.; Sellier, N.; Chabault, M.; Le Bihan-Duval, E.; Narcy, A. Detection of QTL controlling feed efficiency and excretion in chickens fed a wheat-based diet. Genet. Sel. Evol. 2015, 47, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, Q.C.; Wu, P.F.; Dai, G.J.; Zhang, G.X.; Zhang, T.; Xue, Q.; Shi, H.Q.; Wang, J.Y. Identification of 19 loci for reproductive traits in a local Chinese chicken by genome-wide study. Genet. Mol. Res. 2017, 16, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.S.; Ouyang, H.J.; Liu, J.; Chen, B.; Nie, Q.H.; Zhang, X.Q. Expression of variant transcripts of the potassium channel tetramerization domain-containing 15 (KCTD15) gene and their association with fatness traits in chickens. Domest. Anim. Endocrinol. 2015, 50, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zheng, M.Q.; Liu, R.R.; Wen, J.; Wu, D.; Hu, Y.D.; Sun, Y.F.; Li, P.; Liu, L.; Zhao, G.P. Genome-Wide Association of Thymus and Spleen Mass in Chicken. Sci. Agric. Sin. 2012, 45, 3165–3175. [Google Scholar]
- Van Goor, A.; Ashwell, C.M.; Persia, M.E.; Rothschild, M.F.; Schmidt, C.J.; Lamont, S.J. Quantitative trait loci identified for blood chemistry components of an advanced intercross line of chickens under heat stress. BMC Genom. 2016, 17, 287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, Y.; Lin, S.; Mu, H.; Tang, X.; Ou, Y.; Chen, J.; Ma, Y.; Li, Y. Analysis of differentially expressed genes and signaling pathways related to intramuscular fat deposition in skeletal muscle of sex-linked dwarf chickens. Biomed. Res. Int. 2014, 2014, 724274. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, A.; Ito, M.; Chikuma, S.; Akanuma, T.; Nakatsukasa, H. Negative regulation of cytokine signaling in immunity. Cold Spring Harb. Perspect. Biol. 2018, 10, a028571. [Google Scholar] [CrossRef]
- Reynés, B.; Klein Hazebroek, M.; García-Ruiz, E.; Keijer, J.; Oliver, P.; Palou, A. Specific features of the hypothalamic leptin signaling response to cold exposure are reflected in peripheral blood mononuclear cells in rats and ferrets. Front. Physiol. 2017, 8, 581. [Google Scholar] [CrossRef] [Green Version]
- Javanrouh-Aliabad, A.; Vaez Torshizi, R.; Masoudi, A.A.; Ehsani, A. Identification of candidate genes for blood metabolites in Iranian chickens using a genome-wide association study. Br. Poult. Sci. 2018, 59, 381–388. [Google Scholar] [CrossRef]
- Sun, Y.; Li, Q.; Hu, Y.; Sun, Y.; Liu, R.; Zheng, M.; Wen, J.; Li, P.; Liu, L.; Zhao, G. Genomewide association study of immune traits in chicken F2 resource population. J. Anim. Breed. Genet. 2016, 133, 197–206. [Google Scholar] [CrossRef]
- Dumonteil, E.; Barré, H.; Meissner, G. Sarcoplasmic reticulum Ca(2+)-ATPase and ryanodine receptor in cold-acclimated ducklings and thermogenesis. Am. J. Physiol. 1993, 265, C507–C513. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zhao, G.; Liu, R.; Zheng, M.; Hu, Y.; Wu, D.; Zhang, L.; Li, P.; Wen, J. The identification of 14 new genes for meat quality traits in chicken using a genome-wide association study. BMC Genom. 2013, 14, 458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bihan-Duval, E.L.; Hennequet-Antier, C.; Berri, C.; Beauclercq, S.A.; Bourin, M.C.; Boulay, M.; Demeure, O.; Boitard, S. Identification of genomic regions and candidate genes for chicken meat ultimate pH by combined detection of selection signatures and QTL. BMC Genom. 2018, 19, 294. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Lee, S.H.; Lee, D.H.; Manjula, P.; Lee, S.H.; Lee, J.H. Genetic association of DEGS1, ELOVL6, FABP3, FABP4, FASN and SCD genes with fatty acid composition in breast and thigh muscles of Korean native chicken. Anim. Genet. 2020, 51, 344–345. [Google Scholar] [CrossRef] [PubMed]
- Nowack, J.; Giroud, S.; Arnold, W.; Ruf, T. Muscle non-shivering thermogenesis and its role in the evolution of endothermy. Front. Physiol. 2017, 8, 889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balsa, E.; Marco, R.; Perales-Clemente, E.; Szklarczyk, R.; Calvo, E.; Landázuri, M.O.; Enríquez, J.A. NDUFA4 is a subunit of complex IV of the mammalian electron transport chain. Cell Metab. 2012, 16, 378–386. [Google Scholar] [CrossRef] [Green Version]
- Ijiri, D.; Miura, M.; Kanai, Y.; Hirabayashi, M. Increased mass of slow-type skeletal muscles and depressed myostatin gene expression in cold-tolerant chicks. Zool. Sci. 2009, 26, 277–283. [Google Scholar] [CrossRef]
- Bal, N.C.; Muthu, P. Uncoupling of sarcoendoplasmic reticulum calcium ATPase pump activity by sarcolipin as the basis for muscle non-shivering thermogenesis. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2020, 375, 20190135. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.; Lu, H.; Wang, L.; Zhao, J.; Zeng, W.; Zhang, T. Genome-wide identification and transcriptional expression of the METTL21C gene family in chicken. Genes 2019, 10, 628. [Google Scholar] [CrossRef] [Green Version]
- Arnér, E.S.; Holmgren, A. Physiological functions of thioredoxin and thioredoxin reductase. Eur. J. Biochem. 2000, 267, 6102–6109. [Google Scholar] [CrossRef]
- Chen, J.; Ren, X.; Li, L.; Lu, S.; Chen, T.; Tan, L.; Liu, M.; Luo, Q.; Liang, S.; Nie, Q.; et al. Integrative Analyses of mRNA expression profile reveal the Involvement of IGF2BP1 in chicken adipogenesis. Int. J. Mol. Sci. 2019, 20, 2923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, F.; Gao, P.; Wu, W.; Wang, Z.; Yang, J.; Di, J.; Jiang, B.; Su, X. STK25-induced inhibition of aerobic glycolysis via GOLPH3-mTOR pathway suppresses cell proliferation in colorectal cancer. J. Exp. Clin. Cancer Res. 2018, 37, 144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nerstedt, A.; Cansby, E.; Andersson, C.X.; Laakso, M.; Stančáková, A.; Blüher, M.; Smith, U.; Mahlapuu, M. Serine/threonine protein kinase 25 (STK25): A novel negative regulator of lipid and glucose metabolism in rodent and human skeletal muscle. Diabetologia 2012, 55, 1797–1807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, J.; Liu, W.; Liu, Z.L.; Li, N. cDNA cloning, genomic structure, chromosomal mapping and expression analysis of ADIPOQ (adiponectin) in chicken. Cytogenet. Genome Res. 2006, 112, 148–151. [Google Scholar] [CrossRef]
- Mellouk, N.; Ramé, C.; Barbe, A.; Grandhaye, J.; Froment, P.; Dupont, J. Chicken is a useful model to investigate the role of adipokines in metabolic and reproductive diseases. Int. J. Endocrinol. 2018, 2018, 4579734. [Google Scholar] [CrossRef] [Green Version]
- Wei, Q.; Lee, J.H.; Wang, H.; Bongmba, O.Y.N.; Wu, C.S.; Pradhan, G.; Sun, Z.; Chew, L.; Bajaj, M.; Chan, L.; et al. Adiponectin is required for maintaining normal body temperature in a cold environment. BMC Physiol. 2017, 17, 8. [Google Scholar] [CrossRef] [Green Version]
- Ren, Y.; Li, Y.; Yan, J.; Ma, M.; Zhou, D.; Xue, Z.; Zhang, Z.; Liu, H.; Yang, H.; Jia, L.; et al. Adiponectin modulates oxidative stress-induced mitophagy and protects C2C12 myoblasts against apoptosis. Sci. Rep. 2017, 7, 3209. [Google Scholar] [CrossRef] [Green Version]
- Huang, G.; He, C.; Meng, F.; Li, J.; Zhang, J.; Wang, Y. Glucagon-like peptide (GCGL) is a novel potential TSH-releasing factor (TRF) in chickens: I) Evidence for its potent and specific action on stimulating TSH mRNA expression and secretion in the pituitary. Endocrinology 2014, 155, 4568–4580. [Google Scholar] [CrossRef] [Green Version]
- Baez, D.; Raddatz, N.; Ferreira, G.; Gonzalez, C.; Latorre, R. Gating of thermally activated channels. Curr. Top. Membr. 2014, 74, 51–87. [Google Scholar] [CrossRef]
- Tan, C.H.; McNaughton, P.A. The TRPM2 ion channel is required for sensitivity to warmth. Nature 2016, 536, 460–463. [Google Scholar] [CrossRef] [Green Version]
- Kraft, R.; Grimm, C.; Grosse, K.; Hoffmann, A.; Sauerbruch, S.; Kettenmann, H.; Schultz, G.; Harteneck, C. Hydrogen peroxide and ADP-ribose induce TRPM2-mediated calcium influx and cation currents in microglia. Am. J. Physiol. Cell Physiol. 2004, 286, 129–137. [Google Scholar] [CrossRef] [PubMed]
- González-Ramírez, R.; Chen, Y.; Liedtke, W.B.; Morales-Lázaro, S.L. Chapter 8. TRP channels and pain. In Neurobiology of TRP Channels; Emir, T.L.R., Ed.; CRC Press: Boca Raton, FL, USA; Taylor & Francis: Abingdon, UK, 2017. Available online: https://www.ncbi.nlm.nih.gov/books/NBK476120/ (accessed on 7 February 2022). [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population | n | Ho ± SD | He± SD | MAF ± SD | FHOM ± SD |
|---|---|---|---|---|---|
| Russian White | 177 | 0.285 a ± 0.001 | 0.296 a ± 0.001 | 0.270 a ± 0.001 | 0.020 ± 0.016 |
| Amroks | 20 | 0.357 b ± 0.004 | 0.353 b ± 0.001 | 0.223 b ± 0.001 | 0.017 ± 0.005 |
| Population | n | FROH ± SD | MNROH ± SD |
|---|---|---|---|
| Russian White | 177 | 0.083 a ± 0.006 | 25.9 a ± 1.1 |
| Amroks | 20 | 0.105 b ± 0.009 | 18.9 c ± 1.5 |
| GGA | Region (Mb) | Breed | Genes |
|---|---|---|---|
| 1 | 18.07…20.85 | RW | CRELD2, ALG12, MLC1, IL17REL, TRABD, MAPK11, MAPK12, PLXNB2 |
| 143.35…145.21 | RW | SLC10A2, METTL21C, FGF14, NALCN | |
| 2 | 26.01…27.01 | RW | NDUFA4 |
| 47.88…48.93 | RW | FKBP9, PDE1C, PPP1R17, NEUROD6 | |
| Amroks | |||
| 54.86…56.83 | RW | IGFBP1, IGFBP3, NFX1, LRRC14B, RECK, RBFA, HSBP1L1, NFATC1, HNF4G, IL7 | |
| 122.18…126.07 | RW | WWP1, ATP6V0D2, DECR1, RIPK2, OSGIN2, NECAB1, RUNX1T1 | |
| 3 | 36.71…39.95 | RW | RYR2, MIR, GPR137B, ERO1B, EDARADD, LGALS8, GGPS1,COA6, DISC1 |
| 81.27…84.12 | RW | CD109, CGAS, MTO1, OGFRL1, B3GAT2, FAM135A | |
| 4 | 17.82…19.76 | RW | RNF128, HMGB3, MTM1 |
| 5 | 2.14…4.51 | RW, Amroks | PRMT3, NELL1, ANO5, SLC17A6, GAS2, SLC5A12, FIBIN, LGR4, BDNF |
| 6 | 16.65…17.20 | RW | CAMC2G, MYOZ1, SORBS1 |
| 7 | 13.18…14.51 | RW, Amroks | CTLA4, ICOS, CD28, NBEAL1, IDH1, PIKFYVE, PPP1R1C |
| 9 | 5.13…5.88 | RW | HDLBP, STK25, ETV5, DGKG, TRPM2, ADIPOQ, GPR55, CAB39, HES6, PIK3CB |
| 11 | 2.35…2.62 10.20…10.82 | RW, Amroks | HSF4, MBTPS1, NECAB2 CEBPG, KCTD15 |
| 13 | 4.43…5.43 | RW, Amroks | KCNIP1, KCNMB1, LCP2, DOCK2 |
| 15 | 1.30…2.61 | RW | TXNRD2, COMT, ZDHHC8, DGCR8, PGAM5 |
| 18 | 9.13…10.86 | RW | TSPAN10, SLC38A10, NPTX1, USP36, ENGASE, SOCS3, TK1, TMC6, AFMID, GCGR, SIRT7, RYCR1, SPAG9, NPB, CACNA1G, WFIKKN2, ACSF2, CD300LG, LRRC59, FADDS6, MIF4CD, KCTD2 |
| Environmental Conditions | Russian White | Amroks |
|---|---|---|
| Number of chicks | 15 | 15 |
| Standard rearing conditions (30 °C) before cold treatment | 0.858 a ± 0.01 | 0.913 a ± 0.01 |
| Immidiatelly after 30 min of cold treatment (at 10 °C) | 0.884 b ± 0.009 | 0.884 b ± 0.01 |
| Parameters | Russian White | Amroks |
|---|---|---|
| Number of chicks | 15 | 15 |
| Huddling, muscle shivering | no | yes |
| Torpor (in individual cages) | no | yes |
| Changing the oxygen consumption level, % | +10.9 | +15.6 |
| Changing the carbon dioxide output level, % | +13.5 | +13.5 |
| Changing the respiratory coefficient level, % | +3.0 | −3.2 |
| Gene | GGA | Function |
|---|---|---|
| SOCS3 | 18 | In the adipocytokine signaling pathway, SOCS3 affects adipogenesis by inhibiting the expression of LERP (lipoforin receptor) and IRS1 (insulin receptor), participating in the adipocytokine signaling pathway. IGF2BP3 affects the insulin signaling pathway by regulating intramuscular fat deposition [43]. In mammals, the immune system takes the most active part in the transformation of one adipose tissue into another. White adipose tissue macrophages force fat cells to turn into brown adipose tissue cells when the temperature drops. During physical exercise and when the ambient temperature decreases, meteorin-like hormone is released from the muscles, which, through the immune signaling proteins interleukins, acts on macrophages located in adipose tissue. Then, under the action of special signaling proteins, macrophages compel adipose tissue to burn its reserves. SOCS proteins also regulate innate immunity cells [44]. Exposure to cold causes hyperphagia to counteract fat loss associated with lipid mobilization and thermogenic activation. In experiments, chronic cold stress increased the expression of SOCS3 gene mRNA in the hypothalamus and peripheral mononuclear blood cells in rats and ferrets [45]. The selected ROH island overlapped with QTL for metabolic traits: the level of total protein in the blood (QTL:165737) [46], as well as with QTL for immune traits: bursa mass (QTL: 101247) [47]. |
| RYR2 | 3 | High-conductivity Ca2+ release channels, known as ryanodine receptors (RyR), mediate the release of Ca2+ from the endo/sarcoplasmic reticulum. Prolonged exposure of birds to cold causes the development of non-shivering thermogenesis (NST) of muscular origin. NST is characterized by increased heat release, which can be achieved by increasing the ATP-dependent Ca2+ cycle between the sarcoplasmic reticulum (SR) and cytosolic compartments in the muscles. Studies on the effect of prolonged cold exposure on SR function in ducklings have established that the activity of SR Ca2+-ATPase, and the proportion of vesicles containing a Ca2+ release channel sensitive to ryanodine in the muscle gastrocnemius of ducklings, increased by 30–50% in response to prolonged exposure to cold. These results show that the content of two components directly involved in the Ca2+ cycle by SR increases with acclimatization to cold, and this is due to NST [48]. The region of RYR2’s location found as affected by selection in WC chickens overlaps with several QTL associated with the percentage of intramuscular fat (QTL:24481) [49], as well as with the pH of the pectoral muscle (QTL:157164), which reflects the glycogen content in chicken muscles and is associated with diseases of glycogen accumulation in humans [50]. |
| CAMK2G | 6 | This gene participates in the transport of Ca2+ in the sarcoplasmic reticulum in skeletal muscles. In slow-twitch muscles, it participates in the regulation of Ca2+ transport in the sarcoplasmic reticulum (SR), and in fast-twitch muscles, it participates in the control of Ca2+ release from SR by phosphorylation of the ryanodine receptor. The studied region of the genome overlaps with several QTL associated with the fatty acid composition of meat: QTL for the content of stearic acid (QTL:193105), oleic acid (QTL:193106) and linoleic acid (QTL:193113) [51]. The skeletal muscles of birds have a very high ability to absorb circulating fatty acids. Endurance muscle work in birds can actually be provided with up to 95% of the energy derived from lipids. The use of this high-calorie fuel allows birds to successfully activate the mechanisms of NST under the impact of cold stress [52]. |
| NDUFA4 | 2 | This gene is a component of cytochrome-c-oxidase, the last enzyme in the mitochondrial electron transport chain that controls oxidative phosphorylation. Cytochrome-c-oxidase is a component of the respiratory chain that catalyzes the reduction of oxygen to water [53]. According to literature data, in skeletal muscles of cold-adapted chickens, there is an increase in the content of slow-twitch muscle fibers that contain many mitochondria and use oxidative phosphorylation to produce ATP [54,55]. |
| METTL21C | 1 | This gene is closely related to the development of chicken muscles. METTL21C performs regular processing of the protein structure and is also a heat shock protein that acts as an ATP-dependent molecular chaperone. METTL21C can regulate calcium homeostasis in bones and muscles and promote differentiation of myoblasts into muscle tubes, as well as reducing osteocyte apoptosis and regulating intracellular homeostasis [56]. The studied RW-specific ROH island with the METTL21C gene located is also overlapped by the well-known QTL for the pH of the pectoral muscle (QTL:157161), reflecting the glycogen content in the muscles, which, in turn, affects the processes of shivering thermogenesis. |
| TXNRD2 | 15 | Thioredoxins are crucial for the redox regulation of protein function and signal transmission through the redox control of thiols. Mammalian cytosolic thioredoxin performs many functions to protect against oxidative stress and controls growth and apoptosis, but it is also secreted and has co-cytokine and chemokine activities [57]. |
| IGFBP1,3 | 2 | This gene can regulate the expression of genes related to the metabolism of fatty acids and promote the proliferation and differentiation of adipocytes [58]. |
| STK 25 | 9 | This gene is crucial for regulating glucose and insulin homeostasis in the body and accumulation of ectopic lipids. STK25 interacts with GOLPH3 and mediates glycolysis through GOLPH3-regulated mTOR signaling [59]. Partial depletion of STK25 increases the expression of uncoupling protein 3 (Ucp3), which is accompanied by increased lipid oxidation in myoblasts. Additionally, a reduced level of STK25 indirectly improves insulin-stimulated glucose uptake by muscle cells [60]. |
| ADIPOQ | 9 | This gene is involved in the differentiation of adipocytes in mammals and may play a similar role in chickens. The structures of the adiponectin protein domain among the homologs of chickens and mammals are highly conserved [61]. Adipose tissue can secrete hormones called adipokines, which play an important role in regulating metabolic and reproductive processes at both the central and peripheral levels. The metabolic system of chickens is closely related to that of mammals. Glucose is stored in tissues as glycogen and is used for energy production through glycolysis. However, chickens are constantly hyperglycemic and insulin-resistant, which mimics the condition of type 2 diabetes in mammals. Glycemic levels depend on the line, age and sex of birds, and increased obesity in chickens is associated with lower fasting plasma glucose levels, which contrasts with the situation in mammals [62]. Adiponectin is an important regulator of thermogenesis, and it is necessary to maintain body temperature when exposed to cold [63]. Adiponectin is a circulating hormone that is predominantly produced by adipose tissue. Several pharmacological studies have shown that this protein has powerful antidiabetic, antiatherogenic and anti-inflammatory properties. ADIPOQ plays a moderate regulatory role in mitophagy caused by oxidative stress and suppresses apoptosis. These data demonstrate the antioxidant potential of ADIPOQ in skeletal muscle diseases associated with oxidative stress [64]. |
| GCGR | 18 | A glucagon-like peptide (GCGL) encoded by a glucagon-like gene identified in chickens and other lower vertebrates is probably a hypophysiotropic factor in non-mammalian vertebrates. Hypothalamic GCGL is a thyroid-stimulating hormone release factor active in chickens and participates in the control of the pituitary–thyroid axis [65]. |
| TRPM2 | 9 | This class of ion channels, which belongs to the superfamily of transient receptor potentials (TRP) and is present in specialized neurons that are projected onto the skin, evolved as temperature sensors. TRP thermal channels are polymodal receptors. That is, they can be activated by temperature, voltage, pH and chemical agents [66]. TRPM2 initiates a “warm” signal that controls the coolness search behavior. In this regard, it should be noted that the behavioral reactions of neonatal RW chicks under standard conditions of rearing (+30–32 °C) differ markedly from the reaction of chicks of other breeds: they tend to look for a zone with a lower temperature. In the absence of such an opportunity, they try to cool off by bathing in a drinking bowl. TRPM2 is involved in stress-related inflammatory, vascular and neurodegenerative conditions [67]. TRPM2 is a member of the melastatin-related transient potential ion channel receptor subfamily associated with inflammatory and neuropathic pain perception. The main endogenous activator of the TRPM2 channel is ADP-ribose, which has been identified in TRPM2 as a high-temperature sensor, which plays a central role in preventing overheating when the temperature rises. In addition, some reactive oxygen species are positive regulators of TRPM2 activation; thus, this ion channel plays an important role as a sensor of the oxidative response in some cellular systems [68]. Additionally, TRPM2 plays a key role in the development of pathological pain. Thus, in mice deprived of TRF2 expression, the pain response decreased [69]. This gene is supposed to have an effect on reducing the sensitivity of central thermoreceptive structures to low temperature in RW chickens, which contributes to the adaptation of the body in cold environments. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fedorova, E.S.; Dementieva, N.V.; Shcherbakov, Y.S.; Stanishevskaya, O.I. Identification of Key Candidate Genes in Runs of Homozygosity of the Genome of Two Chicken Breeds, Associated with Cold Adaptation. Biology 2022, 11, 547. https://doi.org/10.3390/biology11040547
Fedorova ES, Dementieva NV, Shcherbakov YS, Stanishevskaya OI. Identification of Key Candidate Genes in Runs of Homozygosity of the Genome of Two Chicken Breeds, Associated with Cold Adaptation. Biology. 2022; 11(4):547. https://doi.org/10.3390/biology11040547
Chicago/Turabian StyleFedorova, Elena S., Natalia V. Dementieva, Yuri S. Shcherbakov, and Olga I. Stanishevskaya. 2022. "Identification of Key Candidate Genes in Runs of Homozygosity of the Genome of Two Chicken Breeds, Associated with Cold Adaptation" Biology 11, no. 4: 547. https://doi.org/10.3390/biology11040547
APA StyleFedorova, E. S., Dementieva, N. V., Shcherbakov, Y. S., & Stanishevskaya, O. I. (2022). Identification of Key Candidate Genes in Runs of Homozygosity of the Genome of Two Chicken Breeds, Associated with Cold Adaptation. Biology, 11(4), 547. https://doi.org/10.3390/biology11040547