
Genome Characterisation of an Isoprene-Degrading Alcaligenes sp. Isolated from a Tropical Restored Forest

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Enrichment and Isolation of Isoprene-Degrading Bacteria
2.2. Initial Identification of Isoprene-Degrading Bacteria by 16S rRNA Gene Sequencing
2.3. Test for Isoprene Degradation by Bacterial Isolates
2.4. Amplification of the isoA Gene
2.5. Preparation of Genomic DNA for Genome Analysis
2.6. Genome Sequencing and Genome Assembly
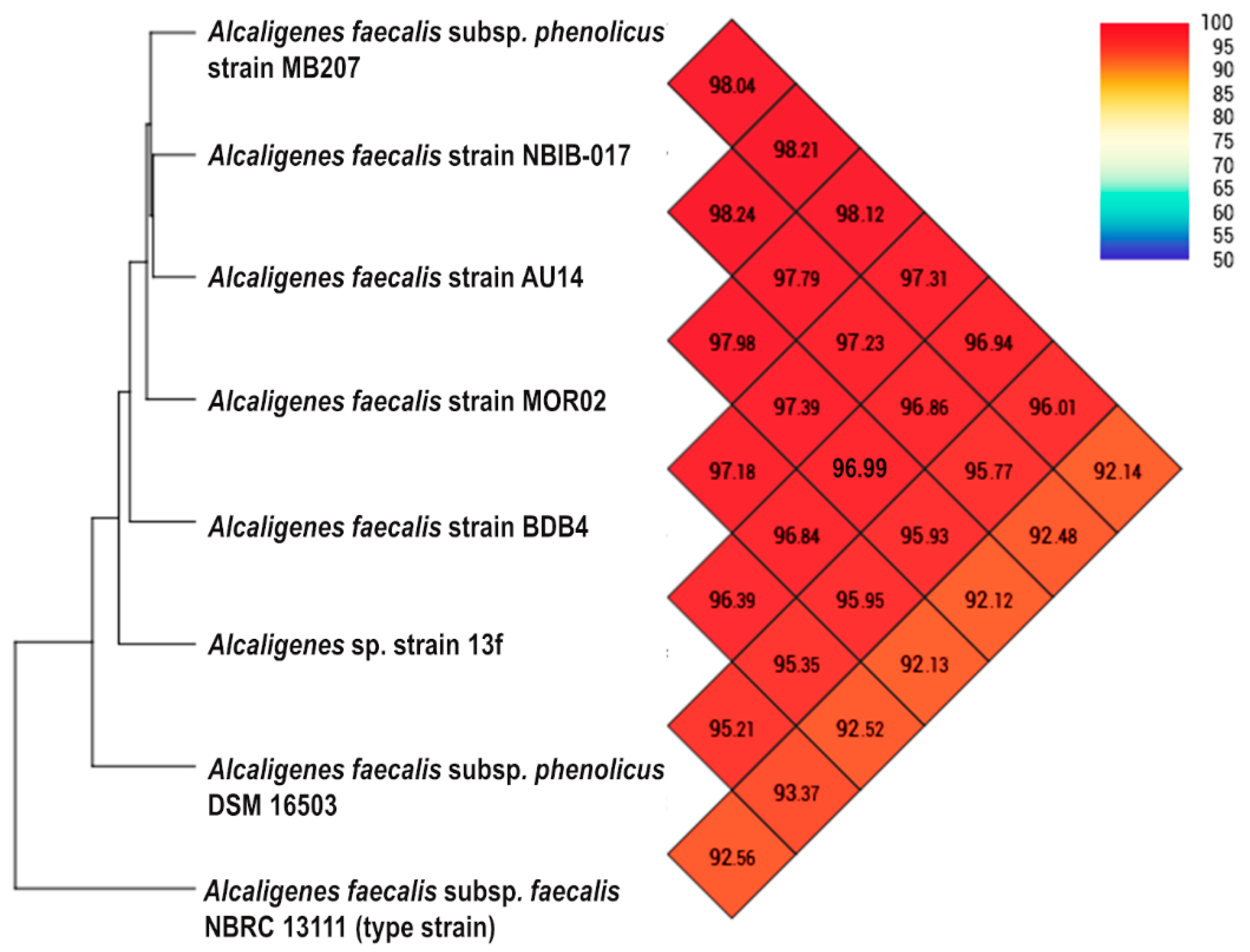
2.7. Phylogenomic Characterisation and Analysis of Average Nucleotide Identity (ANI)
2.8. Genome Analysis and Comparison
3. Results and Discussion
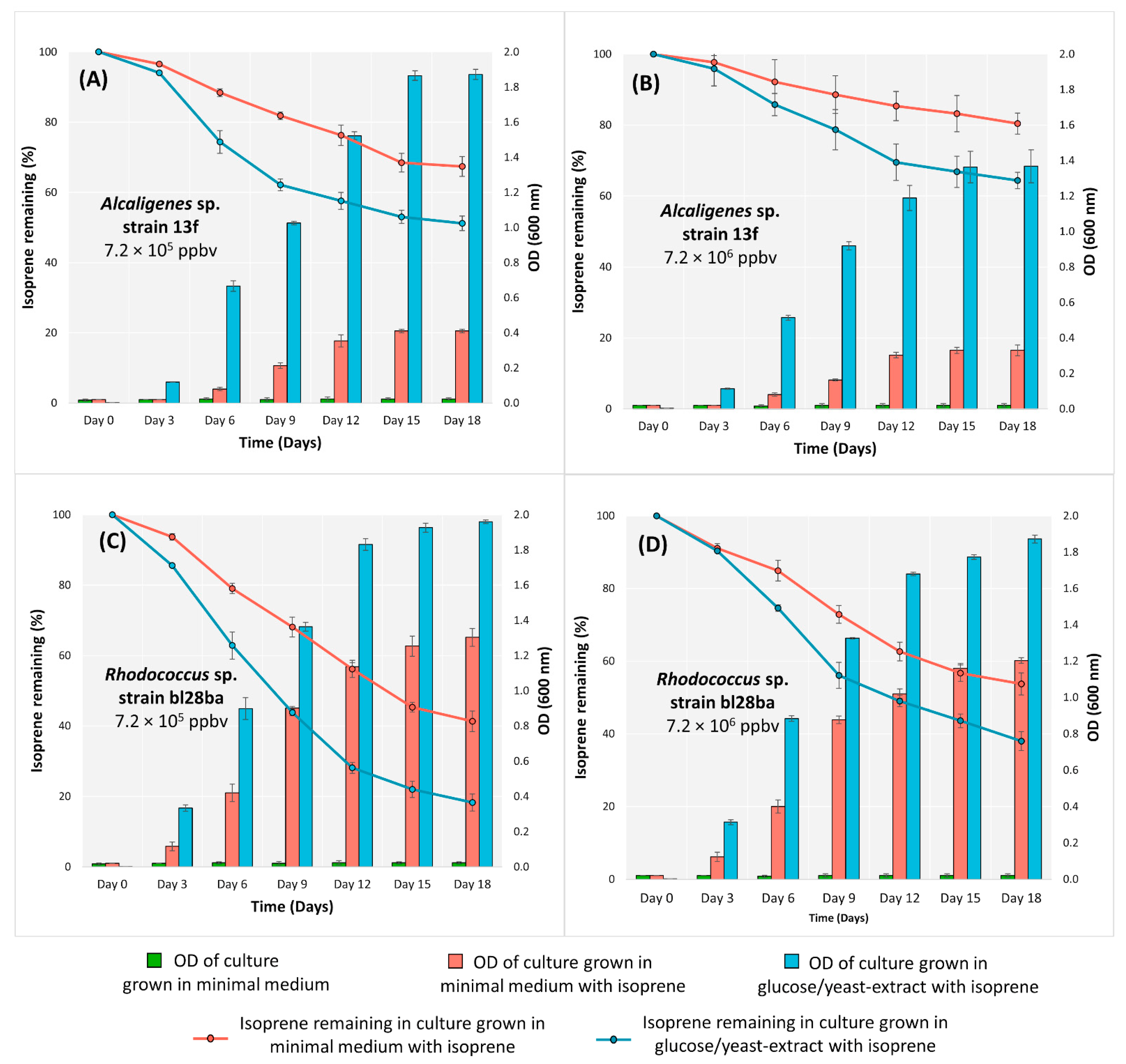
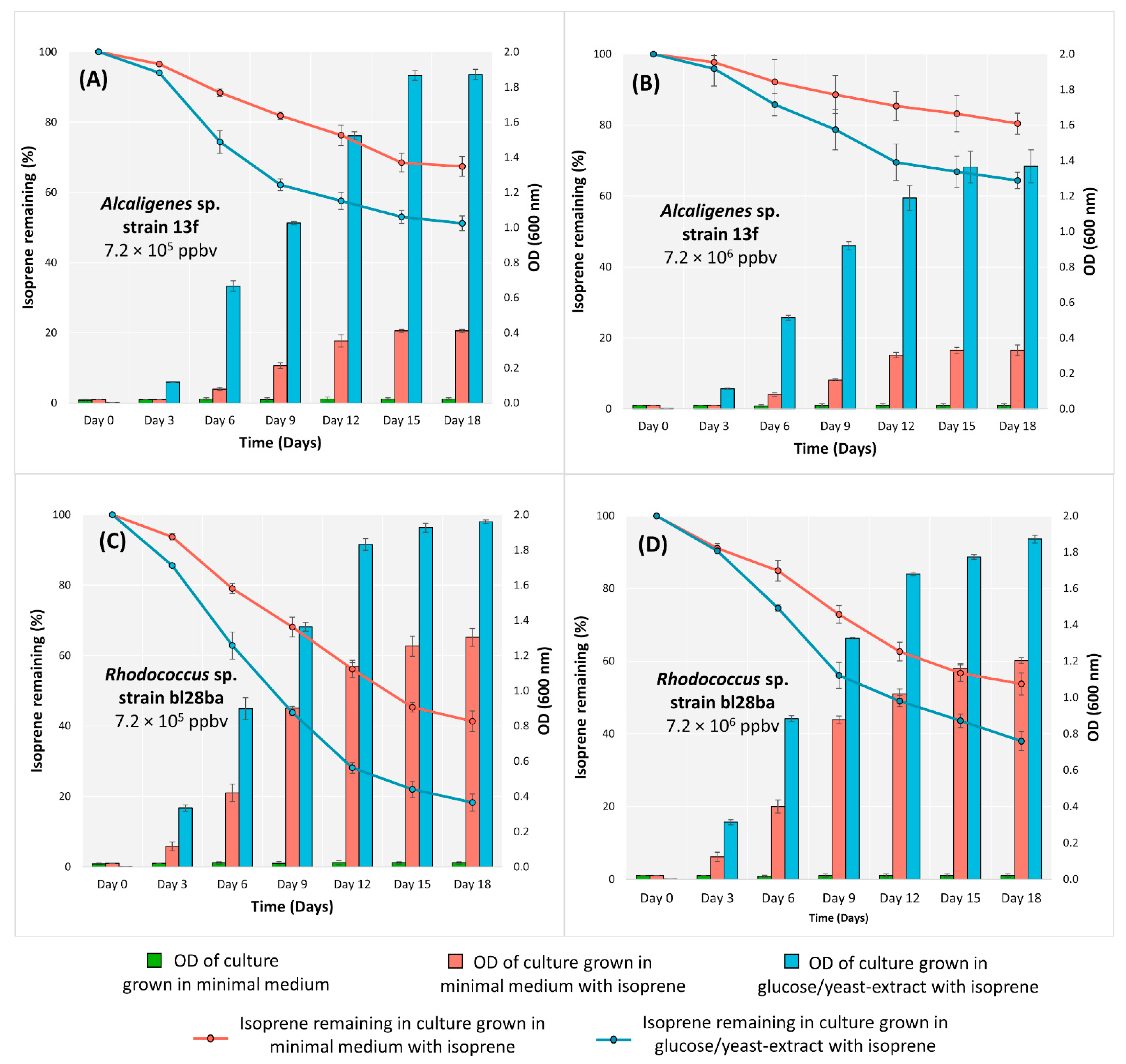
3.1. Growth Characteristics and Isoprene Degradation of Alcaligenes sp. Strain 13f
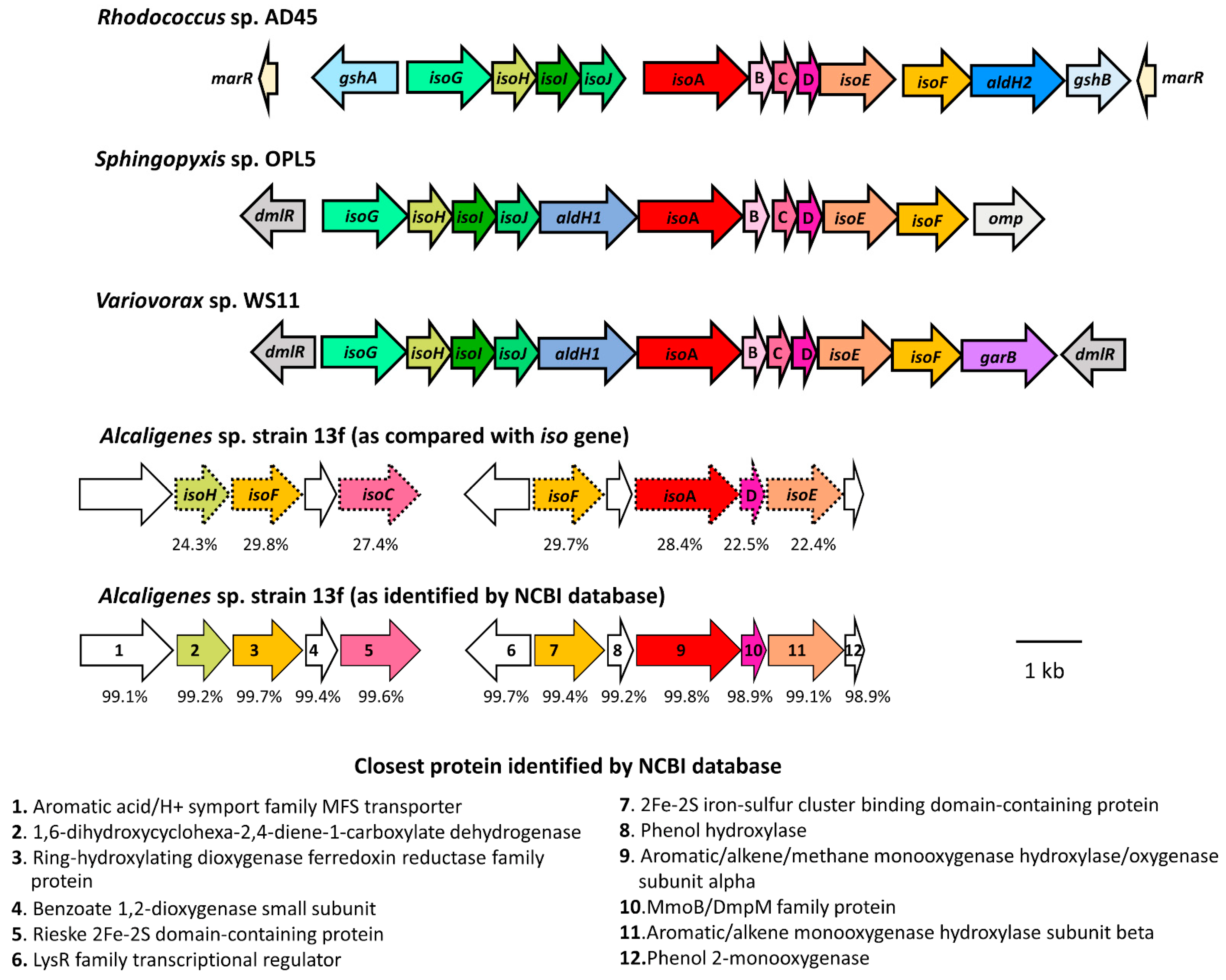
3.2. Examination of isoA Gene in Alcaligenes sp. Strain 13f
3.3. Overall Characteristics of the Genome of Alcaligenes sp. Strain 13f
3.4. Genome Comparison between Alcaligenes sp. Strain 13f and Other Isoprene-Degrading Bacteria
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Müller, J.-F.; Stavrakou, T.; Wallens, S.; De Smedt, I.; Van Roozendael, M.; Potosnak, M.J.; Rinne, J.; Munger, B.; Goldstein, A.; Guenther, A.B. Global isoprene emissions estimated using MEGAN, ECMWF analyses and a detailed canopy environment model. Atmos. Chem. Phys. 2008, 8, 1329–1341. [Google Scholar] [CrossRef] [Green Version]
- Sharkey, T.D.; Wiberley, A.E.; Donohue, A.R. isoprene emission from plants: Why and how. Ann. Bot. 2008, 101, 5–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arneth, A.; Monson, R.K.; Schurgers, G.; Niinemets, Ü.; Palmer, P.I. Why are estimates of global terrestrial isoprene emissions so similar (and why is this not so for monoterpenes)? Atmos. Chem. Phys. 2008, 8, 4605–4620. [Google Scholar] [CrossRef] [Green Version]
- Guenther, A.B.; Jiang, X.; Heald, C.L.; Sakulyanontvittaya, T.; Duhl, T.; Emmons, L.K.; Wang, X. The model of emissions of gases and aerosols from Nature version 2.1 (MEGAN2.1): An extended and updated framework for modeling biogenic emissions. Geosci. Model Dev. 2012, 5, 1471–1492. [Google Scholar] [CrossRef] [Green Version]
- Pacifico, F.; Harrison, S.P.; Jones, C.D.; Sitch, S. Isoprene emissions and climate. Atmos. Environ. 2009, 43, 6121–6135. [Google Scholar] [CrossRef]
- McGenity, T.J.; Crombie, A.T.; Murrell, J.C. Microbial cycling of Isoprene, the most abundantly produced biological volatile organic compound on earth. ISME J. 2018, 12, 931–941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cleveland, C.C.; Yavitt, J.B. Consumption of atmospheric isoprene in soil. Geophys. Res. Lett. 1997, 24, 2379–2382. [Google Scholar] [CrossRef] [Green Version]
- Murrell, J.C.; McGenity, T.J.; Crombie, A.T. Microbial metabolism of isoprene: A much-neglected climate-active gas. Microbiology 2020, 166, 600–613. [Google Scholar] [CrossRef] [PubMed]
- Gray, C.M.; Helmig, D.; Fierer, N. Bacteria and fungi associated with isoprene consumption in soil. Elem. Sci. Anth. 2015, 3, 000053. [Google Scholar] [CrossRef] [Green Version]
- Cleveland, C.C.; Yavitt, J.B. Microbial consumption of atmospheric isoprene in a temperate forest soil. Appl. Environ. Microbiol. 1998, 64, 172–177. [Google Scholar] [CrossRef] [Green Version]
- Pegoraro, E.; Abrell, L.; van Haren, J.; Barron-Gafford, G.; Grieve, K.; Malhi, Y.; Murthy, R.; Lin, G. The effect of elevated atmospheric CO2 and drought on sources and sinks of isoprene in a temperate and tropical rainforest mesocosm. Glob. Change Biol. 2005, 11, 1234–1246. [Google Scholar] [CrossRef]
- Pegoraro, E.; Rey, A.; Abrell, L.; Haren, J.V.; Lin, G. Drought effect on isoprene production and consumption in biosphere 2 tropical rainforest. Glob. Chang. Biol. 2006, 12, 456–469. [Google Scholar] [CrossRef]
- Acuña Alvarez, L.A.; Exton, D.A.; Timmis, K.N.; Suggett, D.J.; McGenity, T.J. Characterization of marine isoprene-degrading communities. Environ. Microbiol. 2009, 11, 3280–3291. [Google Scholar] [CrossRef] [PubMed]
- El Khawand, M.; Crombie, A.T.; Johnston, A.; Vavlline, D.V.; McAuliffe, J.C.; Latone, J.A.; Primak, Y.A.; Lee, S.-K.; Whited, G.M.; McGenity, T.J.; et al. Isolation of isoprene degrading bacteria from soils, development of IsoA gene probes and identification of the active isoprene-degrading soil community using DNA-stable isotope probing: Isoprene-degrading bacteria. Env. Microbiol. 2016, 18, 2743–2753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnston, A.; Crombie, A.T.; Khawand, M.E.; Sims, L.; Whited, G.M.; McGenity, T.J.; Murrell, J.C. Identification and characterisation of isoprene-degrading bacteria in an estuarine environment. Env. Microbiol. 2017, 19, 3526–3537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Hylckama Vlieg, J.E.T.; Leemhuis, H.; Spelberg, J.H.L.; Janssen, D.B. Characterization of the gene cluster involved in isoprene metabolism in Rhodococcus sp. strain AD45. J. Bacteriol. 2000, 182, 1956–1963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibson, L.; Larke-Mej, N.L. Complete genome of isoprene degrading Nocardioides sp. WS12. Microorganisms 2020, 8, 889. [Google Scholar] [CrossRef] [PubMed]
- Dawson, R.A.; Larke-Mejía, N.L.; Crombie, A.T.; Ul Haque, M.F.; Murrell, J.C. Isoprene oxidation by the gram-negative model bacterium Variovorax sp. WS11. Microorganisms 2020, 8, 349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larke-Mejía, N.L.; Carrión, O.; Crombie, A.T.; McGenity, T.J.; Murrell, J.C. Sphingopyxis sp. strain OPL5, an isoprene-degrading bacterium from the Sphingomonadaceae family isolated from oil palm leaves. Microorganisms 2020, 8, 1557. [Google Scholar] [CrossRef] [PubMed]
- Crombie, A.T.; Khawand, M.E.; Rhodius, V.A.; Fengler, K.A.; Miller, M.C.; Whited, G.M.; McGenity, T.J.; Murrell, J.C. Regulation of plasmid-encoded isoprene metabolism in Rhodococcus, a representative of an important link in the global isoprene cycle. Env. Microbiol. 2015, 17, 3314–3329. [Google Scholar] [CrossRef] [Green Version]
- van Hylckama Vlieg, J.E.T.; Kingma, J.; van den Wijngaard, A.J.; Janssen, D.B. A glutathione S-transferase with activity towards cis-1,2-dichloroepoxyethane is involved in isoprene utilization by Rhodococcus sp. strain AD45. Appl. Environ. Microbiol. 1998, 64, 2800–2805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Hylckama Vlieg, J.E.T.; Kingma, J.; Kruizinga, W.; Janssen, D.B. Purification of a glutathione S-transferase and a glutathione conjugate-specific dehydrogenase involved in isoprene metabolism in Rhodococcus sp. strain AD45. J. Bacteriol. 1999, 181, 2094–2101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrión, O.; Larke-Mejía, N.L.; Gibson, L.; Farhan Ul Haque, M.; Ramiro-García, J.; McGenity, T.J.; Murrell, J.C. Gene probing reveals the widespread distribution, diversity and abundance of isoprene-degrading bacteria in the environment. Microbiome 2018, 6, 219. [Google Scholar] [CrossRef] [PubMed]
- Joo, H.-S.; Hirai, M.; Shoda, M. Piggery wastewater treatment using Alcaligenes Faecalis strain No. 4 with heterotrophic nitrification and aerobic denitrification. Water Res. 2006, 40, 3029–3036. [Google Scholar] [CrossRef] [PubMed]
- Van Trappen, S.; Tan, T.-L.; Samyn, E.; Vandamme, P. Alcaligenes aquatilis sp. nov., a novel bacterium from sediments of the Weser Estuary, Germany, and a salt marsh on Shem Creek in Charleston Harbor, USA. Int. J. Syst. Evol. Microbiol. 2005, 55, 2571–2575. [Google Scholar] [CrossRef] [PubMed]
- Abbas, S.; Ahmed, I.; Iida, T.; Lee, Y.-J.; Busse, H.-J.; Fujiwara, T.; Ohkuma, M. A heavy-metal tolerant novel bacterium, Alcaligenes pakistanensis sp. nov., isolated from industrial effluent in Pakistan. Antonie Van Leeuwenhoek 2015, 108, 859–870. [Google Scholar] [CrossRef]
- Mahjoubi, M.; Aliyu, H.; Cappello, S.; Naifer, M.; Souissi, Y.; Cowan, D.A.; Cherif, A. The genome of Alcaligenes aquatilis strain BU33N: Insights into hydrocarbon degradation capacity. PLoS ONE 2019, 14, e0221574. [Google Scholar] [CrossRef] [PubMed]
- Castellani, A.; Chalmers, A.J. Manual of Tropical Medicine, 3rd ed.; Williams Wood and Co.: New York, NK, USA, 1919; pp. 959–960. [Google Scholar]
- Rehfuss, M.; Urban, J. Alcaligenes faecalis subsp. phenolicus subsp. nov. a phenol-degrading, denitrifying bacterium isolated from a graywater bioprocessor. Syst. Appl. Microbiol. 2005, 28, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Lin, D.; Zheng, Y.; Zhang, Q.; Yin, Y.; Cai, L.; Fang, H.; Yu, Y. Biodegradation of DDT by Stenotrophomonas sp. DDT-1: Characterization and genome functional analysis. Sci. Rep. 2016, 6, 21332. [Google Scholar] [CrossRef] [Green Version]
- Ewers, J.; Freier-Schröder, D.; Knackmuss, H.J. Selection of trichloroethene (TCE) degrading bacteria that resist inactivation by TCE. Arch. Microbiol. 1990, 154, 410–413. [Google Scholar] [CrossRef]
- Srivastva, N.; Shukla, A.K.; Singh, R.S.; Upadhyay, S.N.; Dubey, S.K. Characterization of bacterial isolates from rubber dump site and their use in biodegradation of isoprene in batch and continuous bioreactors. Bioresour. Technol. 2015, 188, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Elliott, S.; Navakitbumrung, P.; Kuarak, C.; Zangkum, S.; Anusarnsunthorn, V.; Blakesley, D. Selecting framework tree species for restoring seasonally dry tropical forests in northern Thailand based on field performance. For. Ecol. Manag. 2003, 184, 177–191. [Google Scholar] [CrossRef]
- Uttarotai, T.; McKew, B.A.; Benyahia, F.; Murrell, J.C.; Mhuantong, W.; Wangkarn, S.; Chitov, T.; Bovonsombut, S.; McGenity, T.J. Isoprene-degrading bacteria from soils associated with tropical economic crops and framework forest trees. Microorganisms 2021, 9, 1024. [Google Scholar] [CrossRef]
- Griffiths, R.I.; Whiteley, A.S.; O’Donnell, A.G.; Bailey, M.J. Rapid method for coextraction of DNA and RNA from natural environments for analysis of ribosomal DNA- and rRNA-based microbial community composition. Appl. Environ. Microbiol. 2000, 66, 5488–5491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Window 95/98/NT. Nucl. Acids. Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Murphy, G. Isoprene Degradation in the Terrestrial Environment. Ph.D. Thesis, University of Essex, Colchester, UK, 2017. [Google Scholar]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Wattam, A.R.; Abraham, D.; Dalay, O.; Disz, T.L.; Driscoll, T.; Gabbard, J.L.; Gillespie, J.J.; Gough, R.; Hix, D.; Kenyon, R.; et al. PATRIC, the bacterial bioinformatics database and analysis resource. Nucleic Acids Res. 2014, 42, D581–D591. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, T.; Yamada, K.D.; Tomii, K.; Katoh, K. Parallelization of MAFFT for large-scale multiple sequence alignments. Bioinformatics 2018, 34, 2490–2492. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive tree of life (ITOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, I.; Ouk Kim, Y.; Park, S.-C.; Chun, J. OrthoANI: An improved algorithm and software for calculating average nucleotide identity. Int. J. Syst. Evol. Microbiol. 2016, 66, 1100–1103. [Google Scholar] [CrossRef] [PubMed]
- Mikheenko, A.; Prjibelski, A.; Saveliev, V.; Antipov, D.; Gurevich, A. Versatile genome assembly evaluation with QUAST-LG. Bioinformatics 2018, 34, i142–i150. [Google Scholar] [CrossRef] [PubMed]
- Manni, M.; Berkeley, M.R.; Seppey, M.; Simão, F.A.; Zdobnov, E.M. BUSCO update: Novel and streamlined workflows along with broader and deeper phylogenetic coverage for scoring of eukaryotic, prokaryotic, and viral genomes. Mol. Biol. Evol. 2021, 38, 4647–4654. [Google Scholar] [CrossRef]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI Prokaryotic Genome Annotation Pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Srivastva, N.; Vishwakarma, P.; Bhardwaj, Y.; Singh, A.; Manjunath, K.; Dubey, S.K. Kinetic and molecular analyses reveal isoprene degradation potential of Methylobacterium sp. Bioresour. Technol. 2017, 242, 87–91. [Google Scholar] [CrossRef]
- Lalucat, J.; Mulet, M.; Gomila, M.; García-Valdés, E. Genomics in bacterial taxonomy: Impact on the genus Pseudomonas. Genes 2020, 11, 139. [Google Scholar] [CrossRef] [Green Version]
- Goris, J.; Konstantinidis, K.T.; Klappenbach, J.A.; Coenye, T.; Vandamme, P.; Tiedje, J.M.Y. DNA–DNA hybridization values and their relationship to whole-genome sequence similarities. Int. J. Syst. Evol. Microbiol. 2007, 57, 81–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konstantinidis, K.T.; Tiedje, J.M. Genomic insights that advance the species definition for prokaryotes. Proc. Natl. Acad. Sci. USA 2005, 102, 2567–2572. [Google Scholar] [CrossRef] [Green Version]
- Usyskin-Tonne, A.; Hadar, Y.; Minz, D. Altering N2O emissions by manipulating wheat root bacterial community. Sci. Rep. 2019, 9, 7613. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Heine, T.; Zimmerling, J.; Ballmann, A.; Kleeberg, S.B.; Rückert, C.; Busche, T.; Winkler, A.; Kalinowski, J.; Poetsch, A.; Scholtissek, A.; et al. On the enigma of glutathione-dependent styrene degradation in Gordonia rubripertincta CWB2. Appl. Environ. Microbiol. 2018, 84, e00154-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panke, S.; Witholt, B.; Schmid, A.; Wubbolts, M.G. Towards a biocatalyst for (S)-styrene oxide production: Characterization of the styrene degradation pathway of Pseudomonas sp. strain VLB120. Appl. Environ. Microbiol. 1998, 64, 2032–2043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McClay, K.; Fox, B.G.; Steffan, R.J. Toluene monooxygenase-catalyzed epoxidation of alkenes. Appl. Environ. Microbiol. 2000, 66, 1877–1882. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genomic Information of Alcaligenes sp. Strain 13f | Value |
|---|---|
| Total length (bp) | 4,402,996 |
| GC content (%) | 56.29 |
| Number of contigs | 86 |
| Largest contig (bp) | 320,621 |
| N50 (bp) | 180,050 |
| Completeness (%) | 100 |
| Protein coding sequences | 3942 |
| rRNAs (5S, 16S, 23S) | 1, 1, 1 |
| tRNAs | 53 |
| ncRNAs | 4 |
| Pseudogenes (total) | 38 |
| Alcaligenes sp. Strain 13f Protein Sequence Accession No. | Closest Protein Identified by BLASTp (NCBI Database) (>98% Identity) | Closest Protein in Iso Cluster | % Identity to Amino Acid Sequence in Iso Cluster of Isoprene Degrader | ||
|---|---|---|---|---|---|
| Rhodococcus AD45 | Sphingopyxis OPL5 | Variovorax WS11 | |||
| WP_226348813 | 1,6-dihydroxycyclohexa-2,4-diene-1-carboxylate dehydrogenase | IsoH | - | 23.98 | 24.29 |
| WP_226348814 | Ring-hydroxylating dioxygenase ferredoxin reductase family protein | IsoF | 30.36 | 28.97 | 29.75 |
| WP_226348815 | 2Fe-2S iron-sulfur cluster binding domain-containing protein | IsoF | 29.08 | 29.46 | 29.68 |
| WP_226348817 | Aromatic/alkene/methane monooxygenase hydroxylase/oxygenase subunit alpha | IsoA | 27.77 | 28.09 | 28.36 |
| WP_003800437 | MmoB/DmpM family protein | IsoD | 23.94 | 26.77 | 22.54 |
| WP_226348818 | Aromatic/alkene monooxygenase hydroxylase subunit beta | IsoE | 24.2 | 20.18 | 22.36 |
| WP_226348895 | Rieske 2Fe-2S domain-containing protein | IsoC | - | - | 27.42 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Uttarotai, T.; Sutheeworapong, S.; Crombie, A.T.; Murrell, J.C.; Mhuantong, W.; Noirungsee, N.; Wangkarn, S.; Bovonsombut, S.; McGenity, T.J.; Chitov, T. Genome Characterisation of an Isoprene-Degrading Alcaligenes sp. Isolated from a Tropical Restored Forest. Biology 2022, 11, 519. https://doi.org/10.3390/biology11040519
Uttarotai T, Sutheeworapong S, Crombie AT, Murrell JC, Mhuantong W, Noirungsee N, Wangkarn S, Bovonsombut S, McGenity TJ, Chitov T. Genome Characterisation of an Isoprene-Degrading Alcaligenes sp. Isolated from a Tropical Restored Forest. Biology. 2022; 11(4):519. https://doi.org/10.3390/biology11040519
Chicago/Turabian StyleUttarotai, Toungporn, Sawannee Sutheeworapong, Andrew T. Crombie, J. Colin Murrell, Wuttichai Mhuantong, Nuttapol Noirungsee, Sunanta Wangkarn, Sakunnee Bovonsombut, Terry J. McGenity, and Thararat Chitov. 2022. "Genome Characterisation of an Isoprene-Degrading Alcaligenes sp. Isolated from a Tropical Restored Forest" Biology 11, no. 4: 519. https://doi.org/10.3390/biology11040519
APA StyleUttarotai, T., Sutheeworapong, S., Crombie, A. T., Murrell, J. C., Mhuantong, W., Noirungsee, N., Wangkarn, S., Bovonsombut, S., McGenity, T. J., & Chitov, T. (2022). Genome Characterisation of an Isoprene-Degrading Alcaligenes sp. Isolated from a Tropical Restored Forest. Biology, 11(4), 519. https://doi.org/10.3390/biology11040519