
Reproductive Isolation in the Cryptic Species Complex of a Key Pest: Analysis of Mating and Rejection Behaviour of Onion Thrips (Thrips tabaci Lindeman)




Abstract
Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Insect Cultures and Rearing
2.2. Cross-Mating Experiments
2.3. Behaviour of Mating Pairs and Post-Mating Interactions
2.4. Behaviour of Non-Mating Cross Pairs
2.5. Statistical Analysis
2.5.1. Cross-Mating Experiments
2.5.2. Which Sex Approached, Precopulation and Copulation
2.5.3. Post-Mating Interactions
2.5.4. Non-Mating Pairs
3. Results
3.1. Thrips tabaci Lineage Identification
3.2. Cross-Mating Experiments
3.3. Precopulation and Copulation Behaviour
3.4. Post-Mating Interactions
3.5. Behaviour of Non-Mating Pairs
4. Discussion
4.1. Reproductive Isolation in the Onion Thrips Cryptic Species Complex
4.2. Mating Behaviour and Post-Mating Interactions
4.3. Mating of Thelytokous Females and Hybridization
4.4. Activity of the Lineages
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bickford, D.; Lohman, D.J.; Sodhi, N.S.; Ng, P.K.L.; Meier, R.; Winker, K.; Ingram, K.K.; Das, I. Cryptic species as a window on diversity and conservation. Trends Ecol. Evol. 2007, 22, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Stireman, J.O.; Nason, J.D.; Heard, S.B. Host-associated genetic differentiation in phytophagous insects: General phenomenon or isolated exceptions? Evidence from a goldenrod-insect community. Evolution 2005, 59, 2573–2587. [Google Scholar] [CrossRef] [PubMed]
- Loxdale, H.D.; Harvey, J.A. The ‘generalism’ debate: Misinterpreting the term in the empirical literature focusing on dietary breadth in insects. Biol. J. Linn. Soc. 2016, 119, 265–282. [Google Scholar] [CrossRef]
- Struck, T.H.; Feder, J.L.; Bendiksby, M.; Birkeland, S.; Cerca, J.; Gusarov, V.I.; Kistenich, S.; Larsson, K.-H.; Liow, L.H.; Nowak, M.D.; et al. Finding evolutionary processes hidden in cryptic species. Trends Ecol. Evol. 2018, 33, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Pfenninger, M.; Schwenk, K. Cryptic animal species are homogeneously distributed among taxa and biogeographical regions. BMC Evol. Biol. 2007, 7, 121. [Google Scholar] [CrossRef] [PubMed]
- Cook, M.A.; Ozeroff, S.N.; Fitzpatrick, S.M.; Roitberg, B.D. Host-associated differentiation in reproductive behaviour of cecidomyiid midges on cranberry and blueberry. Entomol. Exp. Appl. 2011, 141, 8–14. [Google Scholar] [CrossRef]
- Milenovic, M.; Wosula, E.N.; Rapisarda, C.; Legg, J.P. Impact of host plant species and whitefly species on feeding behavior of Bemisia tabaci. Front. Plant Sci. 2019, 10, 1. [Google Scholar] [CrossRef]
- Rafter, M.A.; Hereward, J.P.; Walter, G.H. Species limits, quarantine risk and the intrigue of a polyphagous invasive pest with highly restricted host relationships in its area of invasion. Evol. Appl. 2013, 6, 1195–1207. [Google Scholar] [CrossRef]
- Kaiser, L.; Le Ru, B.P.; Kaoula, F.; Paillusson, C.; Capdevielle-Dulac, C.; Obonyo, J.O.; Herniou, E.A.; Jancek, S.; Branca, A.; Calatayud, P.-A.; et al. Ongoing ecological speciation in Cotesia sesamiae, a biological control agent of cereal stem borers. Evol. Appl. 2015, 8, 807–820. [Google Scholar] [CrossRef]
- Smith, L.; Cristofaro, M.; Bon, M.-C.; De Biase, A.; Petanović, R.; Vidović, B. The importance of cryptic species and subspecific populations in classic biological control of weeds: A North American perspective. BioControl 2018, 63, 417–425. [Google Scholar] [CrossRef]
- Kobayashi, N.; Shirai, Y.; Tsurusaki, N.; Tamura, K.; Aotsuka, T.; Katakura, H. Two cryptic species of the phytophagous ladybird beetle Epilachna vigintioctopunctata (Coleoptera: Coccinellidae) detected by analyses of mitochondrial DNA and karyotypes, and crossing experiments. Zool. Sci. 2000, 17, 1159–1166. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Liu, S.-S.; De Barro, P.J.; Xu, J.; Luan, J.-B.; Zang, L.-S.; Ruan, Y.-M.; Wan, F.-H. Asymmetric mating interactions drive widespread invasion and displacement in a whitefly. Science 2007, 318, 1769–1772. [Google Scholar] [CrossRef] [PubMed]
- Luan, J.-B.; De Barro, P.J.; Ruan, Y.-M.; Liu, S.-S. Distinct behavioural strategies underlying asymmetric mating interactions between invasive and indigenous whiteflies. Entomol. Exp. Appl. 2013, 146, 186–194. [Google Scholar] [CrossRef]
- Giska, I.; Sechi, P.; Babik, W. Deeply divergent sympatric mitochondrial lineages of the earthworm Lumbricus rubellus are not reproductively isolated. BMC Evol. Biol. 2015, 15, 217. [Google Scholar] [CrossRef] [PubMed]
- Toon, A.; Daglish, G.J.; Ridley, A.W.; Emery, R.N.; Holloway, J.C.; Walter, G.H. Random mating between two widely divergent mitochondrial lineages of Cryptolestes ferrugineus (Coleoptera: Laemophloeidae): A test of species limits in a phosphine-resistant stored product pest. J. Econ. Entomol. 2016, 109, 2221–2228. [Google Scholar] [CrossRef] [PubMed]
- Butlin, R.; Debelle, A.; Kerth, C.; Snook, R.R.; Beukeboom, L.W.; Castillo Cajas, R.F.; Diao, W.; Maan, M.E.; Paolucci, S.; Weissing, F.J.; et al. What do we need to know about speciation? Trends Ecol. Evol. 2012, 27, 27–39. [Google Scholar] [CrossRef]
- König, K.; Zundel, P.; Krimmer, E.; König, C.; Pollmann, M.; Gottlieb, Y.; Steidle, J.L.M. Reproductive isolation due to prezygotic isolation and postzygotic cytoplasmic incompatibility in parasitoid wasps. Ecol. Evol. 2019, 9, 10694–10706. [Google Scholar] [CrossRef]
- Wongnikong, W.; van Brunschot, S.L.; Hereward, J.P.; De Barro, P.J.; Walter, G.H. Testing mate recognition through reciprocal crosses of two native populations of the whitefly Bemisia tabaci (Gennadius) in Australia. Bull. Entomol. Res. 2020, 110, 328–339. [Google Scholar] [CrossRef]
- Ismail, M.; Brooks, M. Male mating preference of two cryptic species of the herbivorous insect Eccritotarsus catarinensis. Biocontrol Sci. Technol. 2018, 28, 529–543. [Google Scholar] [CrossRef]
- Fernando, L.C.P.; Walter, G.H. Species status of two host-associated populations of Aphytis lingnanensis (Hymenoptera: Aphelinidae) in citrus. Bull. Entomol. Res. 1997, 87, 137–144. [Google Scholar] [CrossRef]
- Nojima, S.; Schal, C.; Webster, F.X.; Santangelo, R.G.; Roelofs, W.L. Identification of the sex pheromone of the german cockroach, Blattella germanica. Science 2005, 307, 1104–1106. [Google Scholar] [CrossRef] [PubMed]
- Howard, R.W.; Blomquist, G.J. Ecological, behavioral, and biochemical aspects of insect hydrocarbons. Annu. Rev. Entomol. 2005, 50, 371–393. [Google Scholar] [CrossRef] [PubMed]
- Jungwirth, S.; Ruther, J.; Pokorny, T. Similar is not the same—mate recognition in a parasitoid wasp. Front. Ecol. Evol. 2021, 9, 646667. [Google Scholar] [CrossRef]
- Buellesbach, J.; Vetter, S.G.; Schmitt, T. Differences in the reliance on cuticular hydrocarbons as sexual signaling and species discrimination cues in parasitoid wasps. Front. Zool. 2018, 15, 22. [Google Scholar] [CrossRef]
- Fattoruso, V.; Anfora, G.; Mazzoni, V. Vibrational communication and mating behavior of the greenhouse whitefly Trialeurodes vaporariorum (Westwood) (Hemiptera: Aleyrodidae). Sci. Rep. 2021, 11, 6543. [Google Scholar] [CrossRef]
- Lewis, T. Thrips: Their Biology, Ecology and Economic Importance; Academic: London, UK, 1973. [Google Scholar]
- Lewis, T. Pest thrips in perspective. In Thrips as Crop Pests; Lewis, T., Ed.; CAB International: Oxon, UK; New York, NY, USA, 1997; pp. 1–13. [Google Scholar]
- Riley, D.G.; Joseph, S.V.; Srinivasan, R.; Diffie, S. Thrips vectors of tospoviruses. J. Integr. Pest Manag. 2011, 1, I1–I10. [Google Scholar] [CrossRef]
- Mound, L.A. Order Thysanoptera Haliday, 1836. In Animal Biodiversity: An Outline of Higher-Level Classification and Survey of Taxonomic Richness (Addenda 2013); Zhang, Z.-Q., Ed.; Zootaxa, 3703; Magnolia Press: Auckland, New Zealand, 2013; pp. 49–50. [Google Scholar] [CrossRef]
- Rugman-Jones, P.F.; Hoddle, M.S.; Stouthamer, R. Nuclear-mitochondrial barcoding exposes the global pest western flower thrips (Thysanoptera: Thripidae) as two sympatric cryptic species in its native California. J. Econ. Entomol. 2010, 103, 877–886. [Google Scholar] [CrossRef]
- Dickey, A.M.; Kumar, V.; Hoddle, M.S.; Funderburk, J.E.; Morgan, J.K.; Jara-Cavieres, A.; Shatters, R.G.J.; Osborne, L.S.; McKenzie, C.L. The Scirtothrips dorsalis species complex: Endemism and invasion in a global pest. PLoS ONE 2015, 10, e0123747. [Google Scholar] [CrossRef]
- Hereward, J.; Hutchinson, J.A.; McCulloch, G.A.; Silva, R.; Walter, G.H. Divergence among generalist herbivores: The Frankliniella schultzei species complex in Australia (Thysanoptera: Thripidae). Arthropod Plant Interact. 2017, 11, 875–887. [Google Scholar] [CrossRef]
- Ghosh, A.; Jagdale, S.S.; Basavaraj; Dietzgen, R.G.; Jain, R.K. Genetics of Thrips palmi (Thysanoptera: Thripidae). J. Pest Sci. 2020, 93, 27–39. [Google Scholar] [CrossRef]
- Heming, B.S. History of the germ line in male and female thrips. In Thrips Biology and Management; Parker, B.L., Skinner, M., Lewis, T., Eds.; Plenum Press: New York, NY, USA, 1995; pp. 505–535. [Google Scholar]
- Boake, C.R.B.; Shelly, T.E.; Kaneshiro, K.Y. Sexual selection in relation to pest-management strategies. Annu. Rev. Entomol. 1996, 41, 211–229. [Google Scholar] [CrossRef] [PubMed]
- Foster, S.P.; Harris, M.O. Behavioral manipulation methods for insect pest-management. Annu. Rev. Entomol. 1997, 42, 123–146. [Google Scholar] [CrossRef] [PubMed]
- Moritz, G. Structrure, growth and development. In Thrips as Crop Pests; Lewis, T., Ed.; CAB International: Oxon, UK; New York, NY, USA, 1997; pp. 15–63. [Google Scholar]
- Vershinina, A.O.; Kuznetsova, V.G. Parthenogenesis in Hexapoda: Entognatha and non-holometabolous insects. J. Zool. Syst. Evol. Res. 2016, 54, 257–268. [Google Scholar] [CrossRef]
- Kirk, W.D.J. Aggregation and mating of thrips in flowers of Calystegia sepium. Ecol. Entomol. 1985, 10, 433–440. [Google Scholar] [CrossRef]
- Terry, I.L.; Gardner, D. Male mating swarms in Frankliniella occidentalis (Pergande) (Thysanoptera: Thripidae). J. Insect Behav. 1990, 3, 133–141. [Google Scholar] [CrossRef]
- Akinyemi, A.O.; Subramanian, S.; Mfuti, D.K.; Pope, T.W.; Tamiru, A.; Kirk, W.D.J. Mating behaviour, mate choice and female resistance in the bean flower thrips (Megalurothrips sjostedti). Sci. Rep. 2021, 11, 14504. [Google Scholar] [CrossRef]
- Kirk, W.D.J.; De Kogel, W.J.; Koschier, E.H.; Teulon, D.A.J. Semiochemicals for thrips and their use in pest management. Annu. Rev. Entomol. 2021, 66, 101–119. [Google Scholar] [CrossRef]
- Terry, I.; Schneider, M. Copulatory behaviour and mating frequency of the western flower thrips, Frankliniella occidentalis (Insecta: Thysanoptera). J. Pure Appl. Zool. 1993, 4, 339–354. [Google Scholar]
- Akinyemi, A.O.; Kirk, W.D.J. Experienced males recognise and avoid mating with non-virgin females in the western flower thrips. PLoS ONE 2019, 14, e0224115. [Google Scholar] [CrossRef]
- Milne, M.; Walter, G.H.; Milne, J.R. Mating behavior and species status of host-associated populations of the polyphagous thrips, Frankliniella schultzei. J. Insect Behav. 2007, 20, 331–346. [Google Scholar] [CrossRef]
- Rafter, M.A.; Walter, G.H. Mate recognition in the south african citrus thrips Scirtothrips aurantii (Faure) and cross-mating tests with populations from Australia and South Africa. J. Insect Behav. 2013, 26, 780–795. [Google Scholar] [CrossRef]
- Krueger, S.; Jilge, M.; Mound, L.; Moritz, G.B. Reproductive behavior of Echinothrips americanus (Thysanoptera: Thripidae). J. Insect Sci. 2017, 17, 66. [Google Scholar] [CrossRef] [PubMed]
- Li, X.-W.; Jiang, H.-X.; Zhang, X.-C.; Shelton, A.M.; Feng, J.-N. Post-mating interactions and their effects on fitness of female and male Echinothrips americanus (Thysanoptera: Thripidae), a new insect pest in China. PLoS ONE 2014, 9, e87725. [Google Scholar] [CrossRef] [PubMed]
- Krueger, S.; Moritz, G.; Lindemann, P.; Radisch, D.; Tschuch, G. Male pheromones influence the mating behavior of Echinothrips americanus. J. Chem. Ecol. 2016, 42, 294–299. [Google Scholar] [CrossRef]
- Li, X.-W.; Fail, J.; Shelton, A.M. Female multiple matings and male harassment and their effects on fitness of arrhenotokous Thrips tabaci (Thysanoptera: Thripidae). Behav. Ecol. Sociobiol. 2015, 69, 1585–1595. [Google Scholar] [CrossRef]
- Zawirska, I. Untersuchungen über zwei biologische typen von Thrips tabaci Lind. (Thysanoptera, Thripidae) in der VR Polen. Arch. Phytopathol. Plant Prot. 1976, 12, 411–422. [Google Scholar] [CrossRef]
- Shelton, A.M.; Wilsey, W.T.; Schmaedick, M.A. Management of onion thrips (Thysanoptera: Thripidae) on cabbage by using plant resistance and insecticides. J. Econ. Entomol. 1998, 91, 329–333. [Google Scholar] [CrossRef]
- Shelton, A.M.; Plate, J.; Chen, M. Advances in control of onion thrips (Thysanoptera: Thripidae) in cabbage. J. Econ. Entomol. 2008, 101, 438–443. [Google Scholar] [CrossRef][Green Version]
- Jenser, G.; Szénási, Á. Review of the biology and vector capability of Thrips tabaci Lindeman (Thysanoptera: Thripidae). Acta Phytopathol. Entomol. Hung. 2004, 39, 137–155. [Google Scholar] [CrossRef]
- Diaz-Montano, J.; Fuchs, M.; Nault, B.A.; Fail, J.; Shelton, A.M. Onion thrips (Thysanoptera: Thripidae): A global pest of increasing concern in onion. J. Econ. Entomol. 2011, 104, 1–13. [Google Scholar] [CrossRef]
- Pappu, H.R.; Jones, R.A.C.; Jain, R.K. Global status of tospovirus epidemics in diverse cropping systems: Successes achieved and challenges ahead. Virus Res. 2009, 141, 219–236. [Google Scholar] [CrossRef] [PubMed]
- Brunner, P.C.; Chatzivassiliou, E.K.; Katis, N.I.; Frey, J.E. Host-associated genetic differentiation in Thrips tabaci (Insecta; Thysanoptera), as determined from mtDNA sequence data. Heredity 2004, 93, 364–370. [Google Scholar] [CrossRef] [PubMed]
- Downie, D.A. Baubles, bangles, and biotypes: A critical review of the use and abuse of the biotype concept. J. Insect Sci. 2010, 10, 176. [Google Scholar] [CrossRef] [PubMed]
- Toda, S.; Murai, T. Phylogenetic analysis based on mitochondrial COI gene sequences in Thrips tabaci Lindeman (Thysanoptera: Thripidae) in relation to reproductive forms and geographic distribution. Appl. Entomol. Zool. 2007, 42, 309–316. [Google Scholar] [CrossRef]
- Kobayashi, K.; Hasegawa, E. Discrimination of reproductive forms of Thrips tabaci (Thysanoptera: Thripidae) by PCR with sequence specific primers. J. Econ. Entomol. 2012, 105, 555–559. [Google Scholar] [CrossRef]
- Jacobson, A.L.; Booth, W.; Vargo, E.L.; Kennedy, G.G. Thrips tabaci population genetic structure and polyploidy in relation to competency as a vector of Tomato Spotted Wilt Virus. PLoS ONE 2013, 8, e54484. [Google Scholar] [CrossRef]
- Fekrat, L.; Manzari, S.; Shishehbor, P. Morphometric and molecular variation in Thrips tabaci Lindeman (Thysanoptera: Thripidae) populations on onion and tobacco in Iran. J. Agric. Sci. Technol. 2014, 16, 1505–1516. [Google Scholar]
- Almási, A.; Tóbiás, I.; Bujdos, L.; Jenser, G. Molecular characterisation of Thrips tabaci Lindeman, 1889 (Thysanoptera: Thripidae) populations in Hungary based on the ITS2 sequences. Acta Zool. Acad. Sci. Hung. 2016, 62, 157–164. [Google Scholar] [CrossRef]
- Farkas, P.; György, Z.; Tóth, A.; Sojnóczki, A.; Fail, J. A simple molecular identification method of the Thrips tabaci (Thysanoptera: Thripidae) cryptic species complex. Bull. Entomol. Res. 2020, 110, 397–405. [Google Scholar] [CrossRef]
- Király, K.D.; Reiter, D.; Farkas, P.; Sojnóczki, A.; Fail, J. A dohánytripsz (Thrips tabaci Lindeman, 1889) fajkomplex. [The onion thrips (Thrips tabaci LINDEMAN, 1889) species complex]. Növényvédelem 2015, 51, 317–324, (In Hungarian, with English Summary). [Google Scholar]
- Fail, J. Speciation in Thrips tabaci LINDEMAN, 1889 (Thysanoptera): The current state of knowledge and its consequences. Pol. J. Entomol. 2016, 85, 93–104. [Google Scholar] [CrossRef][Green Version]
- Li, X.-W.; Fail, J.; Wang, P.; Feng, J.-N.; Shelton, A.M. Performance of arrhenotokous and thelytokous Thrips tabaci (Thysanoptera: Thripidae) on onion and cabbage and its implications on evolution and pest management. J. Econ. Entomol. 2014, 107, 1526–1534. [Google Scholar] [CrossRef] [PubMed]
- Sogo, K.; Miura, K.; Aizawa, M.; Watanabe, T.; Stouthamer, R. Genetic structure in relation to reproduction mode in Thrips tabaci (Insecta: Thysanoptera). Appl. Entomol. Zool. 2015, 50, 73–77. [Google Scholar] [CrossRef]
- Chatzivassiliou, E.K.; Peters, D.; Katis, N.I. The efficiency by which Thrips tabaci populations transmit Tomato spotted wilt virus depends on their host preference and reproductive strategy. Phytopathology 2002, 92, 603–609. [Google Scholar] [CrossRef][Green Version]
- Jenser, G.; Almási, A.; Fail, J.; Tóbiás, I. Differences in the vector efficiency of Thrips tabaci in Europe and North America. Acta Phytopathol. Entomol. Hung. 2011, 46, 311–317. [Google Scholar] [CrossRef]
- Tedeschi, R.; Ciuffo, M.; Mason, G.; Roggero, P.; Tavella, L. Transmissibility of four tospoviruses by a thelytokous population of Thrips tabaci from Liguria, Northwestern Italy. Phytoparasitica 2001, 29, 37–45. [Google Scholar] [CrossRef]
- Westmore, G.C.; Poke, F.S.; Allen, G.R.; Wilson, C.R. Genetic and host-associated differentiation within Thrips tabaci Lindeman (Thysanoptera: Thripidae) and its links to Tomato spotted wilt virus-vector competence. Heredity 2013, 111, 210–215. [Google Scholar] [CrossRef][Green Version]
- Li, X.-W.; Wang, P.; Fail, J.; Shelton, A.M. Detection of gene flow from sexual to asexual lineages in Thrips tabaci (Thysanoptera: Thripidae). PLoS ONE 2015, 10, e0138353. [Google Scholar] [CrossRef][Green Version]
- Kobayashi, K.; Yoshimura, J.; Hasegawa, E. Coexistence of sexual individuals and genetically isolated asexual counterparts in a thrips. Sci. Rep. 2013, 3, 3286. [Google Scholar] [CrossRef]
- IBM Corp. IBM SPSS Statistics for Windows, Version 25.0; IBM Corp: Armonk, NY, USA, 2017. [Google Scholar]
- Microsoft Corporation. Microsoft Excel, Version 2019; Microsoft Corporation: Redmond, WA, USA, 2018; Available online: https://office.microsoft.com/excel (accessed on 3 November 2021).
- Marascuilo, L.A.; McSweeney, M. Nonparametric post hoc comparisons for trend. Psychol. Bull. 1967, 67, 401–412. [Google Scholar] [CrossRef]
- Tabachnick, B.G.; Fidell, L.S. Using Multivariate Statistics, 5th ed.; Pearson Education/Allyn and Bacon: Boston, MA, USA, 2007. [Google Scholar]
- Bonduriansky, R. The evolution of male mate choice in insects: A synthesis of ideas and evidence. Biol. Rev. 2001, 76, 305–339. [Google Scholar] [CrossRef] [PubMed]
- Miller, S.E.; Legan, A.W.; Flores, Z.A.; Ng, H.Y.; Sheehan, M.J. Strong, but incomplete, mate choice discrimination between two closely related species of paper wasp. Biol. J. Linn. Soc. 2019, 126, 614–622. [Google Scholar] [CrossRef] [PubMed]
- Puniamoorthy, N. Behavioural barriers to reproduction may evolve faster than sexual morphology among populations of a dung fly (Sepsidae). Anim. Behav. 2014, 98, 139–148. [Google Scholar] [CrossRef]
- Engqvist, L.; Sauer, K.P. Determinants of sperm transfer in the scorpionfly Panorpa cognata: Male variation, female condition and copulation duration. J. Evol. Biol. 2003, 16, 1196–1204. [Google Scholar] [CrossRef] [PubMed]
- Reinhold, K.; Engqvist, L.; Consul, A.; Ramm, S.A. Male birch catkin bugs vary copula duration to invest more in matings with novel females. Anim. Behav. 2015, 109, 161–166. [Google Scholar] [CrossRef]
- Mendelson, T.C.; Shaw, K.L. The (mis)concept of species recognition. Trends Ecol. Evol. 2012, 27, 421–427. [Google Scholar] [CrossRef]
- Olaniran, O.A.; Sudhakar, A.V.S.; Drijfhout, F.P.; Dublon, I.A.N.; Hall, D.R.; Hamilton, J.G.C.; Kirk, W.D.J. A male-predominant cuticular hydrocarbon, 7-methyltricosane, is used as a contact pheromone in the western flower thrips Frankliniella occidentalis. J. Chem. Ecol. 2013, 39, 559–568. [Google Scholar] [CrossRef]
- Chen, T.; Li, Q.; Qi, G.; Gao, Y.; Zhao, C.; Lu, L. Cuticular hydrocarbon pattern as a chemotaxonomy marker to assess six species of thrips. J. Asia Pac. Entomol. 2020, 23, 1255–1263. [Google Scholar] [CrossRef]
- Wiley, E.O. The evolutionary species concept reconsidered. Syst. Zool. 1978, 27, 17–26. [Google Scholar] [CrossRef]
- de Queiroz, K. Species concepts and species delimitation. Syst. Biol. 2007, 56, 879–886. [Google Scholar] [CrossRef]
- Sobel, J.M.; Chen, G.F.; Watt, L.R.; Schemske, D.W. The biology of speciation. Evolution 2010, 64, 295–315. [Google Scholar] [CrossRef] [PubMed]
- Forbes, A.A.; Devine, S.N.; Hippee, A.C.; Tvedte, E.S.; Ward, A.K.G.; Widmayer, H.A.; Wilson, C.J. Revisiting the particular role of host shifts in initiating insect speciation. Evolution 2017, 71, 1126–1137. [Google Scholar] [CrossRef] [PubMed]
- Kirk, W.D.J.; Hamilton, J.G.C. Evidence for a male-produced sex pheromone in the western flower thrips Frankliniella occidentalis. J. Chem. Ecol. 2004, 30, 167–174. [Google Scholar] [CrossRef]
- Zhang, P.-J.; Zhu, X.-Y.; Lu, Y.-B. Behavioural and chemical evidence of a male-produced aggregation pheromone in the flower thrips Frankliniella intonsa. Physiol. Entomol. 2011, 36, 317–320. [Google Scholar] [CrossRef]
- Krueger, S.; Wilfer, A.; Tschuch, G.; Lindemann, P.; Moritz, G. First detection of a female-specific volatile substance in thrips. In Proceedings of the XIth International Symposium on Thysanoptera and Tospoviruses, Kunming, China, 21–25 September 2019. [Google Scholar]
- Akinyemi, A.O. The Copulation Behaviour of the Western Flower Thrips. Ph.D. Thesis, Keele University, Keele, UK, 2018. [Google Scholar]
- Magnhagen, C. Predation risk as a cost of reproduction. Trends Ecol. Evol. 1991, 6, 183–186. [Google Scholar] [CrossRef]
- Gwynne, D.T. Does copulation increase the risk of predation? Trends Ecol. Evol. 1989, 4, 54–56. [Google Scholar] [CrossRef]
- Fairbairn, D.J. Costs of loading associated with mate-carrying in the waterstrider, Aquarius remigis. Behav. Ecol. 1993, 4, 224–231. [Google Scholar] [CrossRef]
- Kemp, D.J. Costly copulation in the wild: Mating increases the risk of parasitoid-mediated death in swarming locusts. Behav. Ecol. 2012, 23, 191–194. [Google Scholar] [CrossRef][Green Version]
- Almbro, M.; Kullberg, C. The downfall of mating: The effect of mate-carrying and flight muscle ratio on the escape ability of a pierid butterfly. Behav. Ecol. Sociobiol. 2009, 63, 413–420. [Google Scholar] [CrossRef]
- Lehmann, G.U.C.; Siozios, S.; Bourtzis, K.; Reinhold, K.; Lehmann, A.W. Thelytokous parthenogenesis and the heterogeneous decay of mating behaviours in a bushcricket (Orthopterida). J. Zool. Syst. Evol. Res. 2011, 49, 102–109. [Google Scholar] [CrossRef]
- van der Kooi, C.J.; Schwander, T. On the fate of sexual traits under asexuality. Biol. Rev. 2014, 89, 805–819. [Google Scholar] [CrossRef]
- Kumm, S. Reproduction, Progenesis, and Embryogenesis of Thrips (Thysanoptera, Insecta). Ph.D. Thesis, Martin-Luther-Universitat, Halle-Wittenberg, Halle, Germany, 2002. [Google Scholar]
- Del Bene, G.; Cavallo, V.; Lupetti, P.; Dallai, R. Ultrastructure of the accessory gland in the parthenogenetic thrips Heliothrips haemorrhoidalis (Bouché) (Thysanoptera: Thripidae). Int. J. Insect Morphol. Embryol. 1998, 27, 255–261. [Google Scholar] [CrossRef]
- Arakaki, N.; Miyoshi, T.; Noda, H. Wolbachia-mediated parthenogenesis in the predatory thrips Franklinothrips vespiformis (Thysanoptera: Insecta). Proc. R. Soc. Lond. B Biol. Sci. 2001, 268, 1011–1016. [Google Scholar] [CrossRef] [PubMed]
- Nakao, S.; Yabu, S. Ethological and chemical discrimination between thelytokous and arrhenotokous Thrips nigropilosus Uzel, with discussion of taxonomy. Jpn. J. Appl. Entomol. Zool. 1998, 42, 77–83. [Google Scholar] [CrossRef]
- Normark, B.B. The evolution of alternative genetic systems in insects. Annu. Rev. Entomol. 2003, 48, 397–423. [Google Scholar] [CrossRef]
- Aizawa, M.; Watanabe, T.; Kumano, A.; Miyatake, T.; Sonoda, S. Cypermethrin resistance and reproductive types in onion thrips, Thrips tabaci (Thysanoptera: Thripidae). J. Pestic. Sci. 2016, 41, 167–170. [Google Scholar] [CrossRef]
- Monceau, K.; Moreau, J.; Poidatz, J.; Bonnard, O.; Thiéry, D. Behavioral syndrome in a native and an invasive hymenoptera species. Insect Sci. 2015, 22, 541–548. [Google Scholar] [CrossRef]
- Rodrigues, A.S.; Botina, L.; Nascimento, C.P.; Gontijo, L.M.; Torres, J.B.; Guedes, R.N.C. Ontogenic behavioral consistency, individual variation and fitness consequences among lady beetles. Behav. Process. 2016, 131, 32–39. [Google Scholar] [CrossRef]
- Kralj-Fišer, S.; Hebets, E.A.; Kuntner, M. Different patterns of behavioral variation across and within species of spiders with differing degrees of urbanization. Behav. Ecol. Sociobiol. 2017, 71, 125. [Google Scholar] [CrossRef]
- Riefler, J.; Koschier, E.H. Comparing behavioural patterns of Thrips tabaci Lindeman on leek and cucumber. J. Insect Behav. 2009, 22, 111–120. [Google Scholar] [CrossRef]
- Reitz, S.R.; Funderburk, J.E.; Waring, S.M. Differential predation by the generalist predator Orius insidiosus on congeneric species of thrips that vary in size and behavior. Entomol. Exp. Appl. 2006, 119, 179–188. [Google Scholar] [CrossRef]
- Kirk, W.D.J. Pollen-feeding in thrips (Insecta: Thysanoptera). J. Zool. 1984, 204, 107–117. [Google Scholar] [CrossRef]
- Isenhour, D.J.; Yeargan, K.V. Interactive behavior of Orius insidiosus [Hem.: Anthocoridae] and Sericothrips variabilis [Thys.: Thripidae]: Predator searching strategies and prey escape tactics. Entomophaga 1981, 26, 213–219. [Google Scholar] [CrossRef]
- González-Orellana, J.; López-Guillén, G.; Malo, E.A.; Goldarazena, A.; Cruz-López, L. Behavioural and electrophysiological responses of Liothrips jatrophae (Thysanoptera: Phlaeothripidae) to conspecific extracts and some of its identified compounds. Physiol. Entomol. 2021. early view, online version of record before inclusion in an issue. [Google Scholar] [CrossRef]

{kind=link}
| Steps of the Mating Behaviour Sequence | Description |
|---|---|
| Phase 1: Interaction | The specimens are close enough (approximately 1 mm) to sense each other, and at least one of them shows an obvious change in behaviour. |
| Phase 2: Contact | Physical contact, irrespective of which specimen touched the other. |
| Phase 3: Male mounting | Male starts to climb on the back of the female. |
| Phase 4: Bend abdomen | Male curls his abdomen beneath that of the female. |
| Phase 5: Successful mating | Prolonged contact between the end of the male and female abdomen. |
| Behaviour Parameters Observed | Description |
|---|---|
| Interaction event | The specimens are close enough (approximately 1 mm) to sense each other, and at least one of them shows an obvious change in behaviour, but they do not necessarily contact physically, although the interaction event often starts with physical contact. The end of a given interaction event is when the specimens depart from each other for more than 2 mm without an obvious sign of looking after each other, or they might stay closer but without any sign of sensing each other. |
| The sex of the approaching specimen | The sex of the thrips that approached the other at the start of a given interaction event. |
| Contact position | Position of male and female during initial contact (head to head, female head to male thorax or abdomen, male head to female thorax or abdomen). |
| Female rejection | Female arching and/or flipping her abdomen up and down when a male contacts or mounts her. |
| Male rejection | Male starts to climb on the back of the female, but he stops later and discontinues mating with the female, and either reverses the movement immediately, or climbs down from the female’s back a few seconds later. |
| Escape response | At the end of a given interaction event, the specimen leaves the area in an excited manner (for example by running away), usually with a rapid change in behaviour. |
| Male identification behaviour | In close proximity to the female, for a brief period of time movement of any body part of the male is paused, including walking or running and moving the antennae, which are usually held fixed more or less high. |
| Step in the Mating Behaviour Sequence | T♀ + T♂ | L1♀ + L1♂ | L2♀ + L1♂ | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Both Specimens Used for the First Time | One or Both Specimens Used for the Second Time | p Value | Both Specimens Used for the First Time | One or Both Specimens Used for the Second Time | p Value | Both Specimens Used for the First Time | One or Both Specimens Used for the Second Time | p Value | |
| Interaction | 75% | 50% | =0.128 | 90% | 70% | =0.212 | 80% | 77% | =0.572 |
| (n = 12) | (n = 30) | (n = 10) | (n = 23) | (n = 15) | (n = 26) | ||||
| Successful mating | 67% | 47% | =0.204 | 80% | 65% | =0.339 | 53% | 54% | =0.614 |
| (n = 12) | (n = 30) | (n = 10) | (n = 23) | (n = 15) | (n = 26) | ||||
| Successful mating (if interaction occurred) | 89% | 93% | =0.620 | 89% | 94% | =0.600 | 67% | 70% | =0.573 |
| (n = 9) | (n = 15) | (n = 9) | (n = 16) | (n = 12) | (n = 20) | ||||
| Step in the Mating Behaviour Sequence | T♀ + T♂ | T♀ + L1♂ | L1♀ + L1♂ | L1♀ + T♂ | L2♀ + L1♂ | L2♀ + T♂ | p Value |
|---|---|---|---|---|---|---|---|
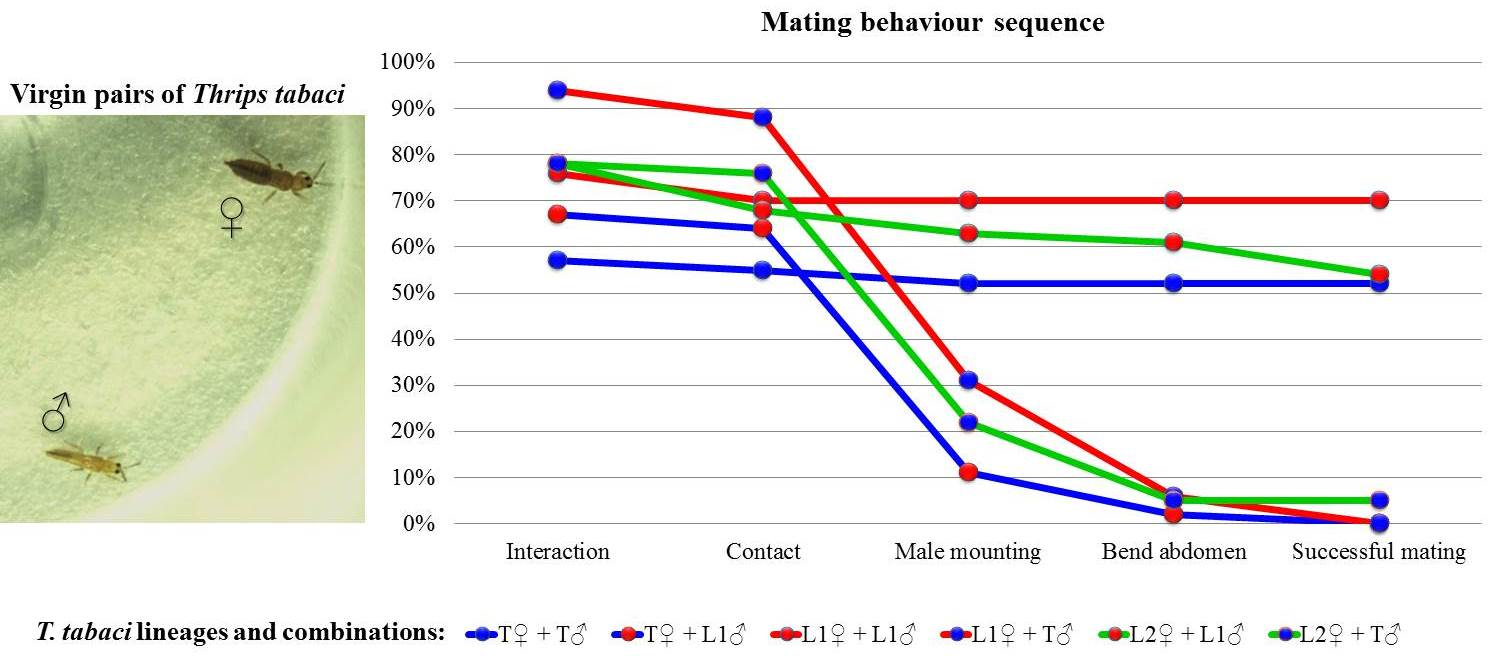
| Interaction | 57% | 67% | 76% | 94% | 78% | 78% | <0.010 |
| (n = 42) | (n = 45) | (n = 33) | (n = 32) | (n = 41) | (n = 41) | ||
| a | ab | ab | b | ab | ab | ||
| Contact | 96% | 97% | 92% | 93% | 88% | 97% | =0.720 |
| (n = 24) | (n = 30) | (n = 25) | (n = 30) | (n = 32) | (n = 32) | ||
| Male mounting | 96% | 17% | 100% | 36% | 93% | 29% | <0.001 |
| (n = 23) | (n = 29) | (n = 23) | (n = 28) | (n = 28) | (n = 31) | ||
| b | a | b | a | b | a | ||
| Bend abdomen | 100% | 20% | 100% | 20% | 96% | 22% | <0.001 |
| (n = 22) | (n = 5) | (n = 23) | (n = 10) | (n = 26) | (n = 9) | ||
| b | a | b | a | b | a | ||
| Successful mating 1 | 100% | 0% | 100% | 0% | 88% | 100% | =0.103 |
| (n = 22) | (n = 1) | (n = 23) | (n = 2) | (n = 25) | (n = 2) | ||
| Remating 2 | 14% | N.A. | 9% | N.A. | 0% | 0% | N.A. |
| (n = 22) | (n = 23) | (n = 22) | (n = 2) |
| T♀ + T♂ | T♀ + L1♂ | L1♀ + L1♂ | L1♀ + T♂ | L2♀ + L1♂ | L2♀ + T♂ | p Value | |
|---|---|---|---|---|---|---|---|
| Time until first interaction event (s) | 110.75 ± 131.88 | 149.43 ± 157.57 | 90.20 ± 107.85 | 98.43 ± 127.62 | 97.59 ± 130.37 | 80.09 ± 91.61 | =0.677 |
| (n = 24) | (n = 30) | (n = 25) | (n = 30) | (n = 32) | (n = 32) | ||
| Time between first contact and first male mounting (s) | 4.95 ± 12.42 | 157.60 ± 88.63 | 52.70 ± 144.01 | 168.50 ± 183.70 | 14.62 ± 29.73 | 113.78 ± 175.82 | <0.001 |
| (n = 22) | (n = 5) | (n = 23) | (n = 10) | (n = 26) | (n = 9) | ||
| a | c | ab | bc | ab | bc |
| Which Sex Approached | p Values of Comparisons | |||||
|---|---|---|---|---|---|---|
| Male Was Approaching | Female Was Approaching | Both Were Approaching | Male vs. Female | Male vs. Both | Female vs. Both | |
| T♀ + T♂ | 76.5% | 23.5% | 0.0% | <0.050 | <0.001 | =0.125 |
| (n = 17) | b | a | a | |||
| T♀ + L1♂ | 71.4% | 17.9% | 10.7% | <0.010 | <0.001 | =0.727 |
| (n = 28) | b | a | a | |||
| L1♀ + L1♂ | 42.9% | 52.4% | 4.8% | =0.824 | <0.050 | <0.010 |
| (n = 21) | b | b | a | |||
| L1♀ + T♂ | 14.3% | 75.0% | 10.7% | <0.010 | >0.999 | <0.001 |
| (n = 28) | a | b | a | |||
| L2♀ + L1♂ | 73.1% | 19.2% | 7.7% | <0.010 | <0.001 | =0.453 |
| (n = 26) | b | a | a | |||
| L2♀ + T♂ | 23.3% | 66.7% | 10.0% | <0.050 | =0.344 | <0.001 |
| (n = 30) | a | b | a | |||
| Which Approached | p Values of Comparisons | ||||||
|---|---|---|---|---|---|---|---|
| Male Was Approaching | Female Was Approaching | Both Were Approaching | Male vs. Female | Male vs. Both | Female vs. Both | ||
| T♀ + T♂ (n = 15) | Percentage | 80.0% | 13.3% | 6.7% | <0.050 | <0.010 | >0.999 |
| b | a | a | |||||
| Adj. residuals | 1.4 | −1.5 | −0.1 | ||||
| L1♀ + L1♂ (n = 20) | Percentage | 50.0% | 45.0% | 5.0% | >0.999 | <0.050 | <0.050 |
| b | b | a | |||||
| Adj. residuals | 1.7 | 2.2 * | −0.5 | ||||
| L2♀ + L1♂ (n = 19) | Percentage | 68.4% | 21.1% | 10.5% | <0.050 | <0.010 | =0.687 |
| b | a | a | |||||
| Adj. residuals | 0.4 | −0.8 | 0.6 | ||||
| T♀ + T♂ (n = 21) | L1♀ + L1♂ (n = 20) | L2♀ + L1♂ (n = 19) | |||
|---|---|---|---|---|---|
| Percentage | p Value (One-Sample z-Test) | Percentage | p Value (One-Sample z-Test) | Percentage | p Value (One-Sample z-Test) |
| 95.2% | <0.001 | 80% | <0.050 | 89.5% | <0.010 |
| Contact Position | p Values of Comparisons | |||||
|---|---|---|---|---|---|---|
| Head to Head | Male Head to Female Thorax or Abdomen | Female Head to Male Thorax or Abdomen | Male Head to Female vs. Female Head to Male | Male Head to Female vs. Head to Head | Female Head to Male vs. Head to Head | |
| T♀ + T♂ (n = 22) | 45.5% | 54.5% | 0.0% | <0.001 | =0.832 | <0.001 |
| b | b | a | ||||
| L1♀ + L1♂ (n = 23) | 47.8% | 39.1% | 13.0% | =0.146 | =0.824 | =0.057 |
| a | a | a | ||||
| L2♀ + L1♂ (n = 22) | 40.9% | 40.9% | 18.2% | =0.267 | >0.999 | =0.267 |
| a | a | a | ||||
| Rejected | ||
|---|---|---|
| T♀ + T♂ (n = 22) | Percentage | 40.9% |
| Adj. residuals | −0.10 | |
| L1♀ + L1♂ (n = 23) | Percentage | 26.1% |
| Adj. residuals | −1.88 + | |
| L2♀ + L1♂ (n = 22) | Percentage | 59.1% |
| Adj. residuals | 2.01 * |
| Mean Duration of | T♀ + T♂ | L1♀ + L1♂ | L2♀ + L1♂ | df | F | p Value |
|---|---|---|---|---|---|---|
| (n = 21) | (n = 21) | (n = 21) | ||||
| Precopulation 1 | 15.67 ± 13.15 | 13.05 ± 7.22 | 16.29 ± 9.42 | 2;60 | 0.637 | =0.532 |
| Precopulation 2 | 11.10 ± 9.91 | 10.19 ± 4.95 | 14.86 ± 9.66 | 2;60 | 1.510 | =0.229 |
| Precopulation 3 | 7.71 ± 10.00 | 5.67 ± 3.62 | 9.57 ± 8.63 | 2;60 | 1.650 | =0.201 |
| Copulation | 176.10 ± 26.02 | 140.43 ± 16.64 | 155.57 ± 13.90 | 2;60 | 18.194 | <0.001 |
| c | a | b |
| T♀ + T♂ | L1♀ + L1♂ | L2♀ + L1♂ | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Mean Ratio of Interaction Events with | Before Copulation | After Copulation | p Value | Before Copulation | After Copulation | p Value | Before Copulation | After Copulation | p Value |
| Male mounting 1 | 0.94 | 0.65 | <0.001 | 0.81 | 0.47 | <0.001 | 0.89 | 0.75 | =0.169 |
| (n = 22) | (n = 20) | (n = 23) | (n = 15) | (n = 22) | (n = 22) | ||||
| Male rejection 2 | 0.05 | 0.76 | <0.001 | 0.00 | 0.81 | <0.001 | 0.05 | 0.88 | <0.001 |
| (n = 22) | (n = 20) | (n = 23) | (n = 13) | (n = 22) | (n = 22) | ||||
| Female rejection 2 | 0.41 | 0.87 | <0.001 | 0.28 | 0.29 | =0.844 | 0.45 | 0.69 | =0.107 |
| (n = 22) | (n = 20) | (n = 23) | (n = 13) | (n = 22) | (n = 22) | ||||
| Category 1: | Category 2: | Category 3: | Category 4: | Category 5: | ||
|---|---|---|---|---|---|---|
| 0 Interaction Event | 1 Interaction Event | 2–6 Interaction Events | 7–9 Interaction Events | 10 or More Interaction Events | ||
| T♀ + L1♂ (n = 45) | Percentage | 33.3% | 20.0% | 31.1% | 8.9% | 6.7% |
| Adj. residuals | 2.2 * | 1.1 | −0.8 | 0.1 | −2.5 * | |
| L1♀ + T♂ (n = 32) | Percentage | 6.3% | 9.4% | 46.9% | 3.1% | 34.4% |
| Adj. residuals | −2.6 ** | −1.1 | 1.6 | −1.3 | 2.8 ** | |
| L2♀ + T♂ (n = 39) | Percentage | 23.1% | 15.4% | 30.8% | 12.8% | 17.9% |
| Adj. residuals | 0.1 | <0.1 | −0.7 | 1.1 | <0.1 | |
| T♀ + L1♂ | L1♀ + T♂ | L2♀ + T♂ | p Value | |||||
|---|---|---|---|---|---|---|---|---|
| Escape Response | No Escape Response | Escape Response | No Escape Response | Escape Response | No Escape Response | |||
| Females | Percentage | 36.8% | 63.2% | 8.4% | 91.6% | 18.3% | 81.6% | <0.001 |
| (n = 35) | (n = 60) | (n = 12) | (n = 131) | (n = 22) | (n = 98) | |||
| Adj. residuals | 5.1 *** | −5.1 *** | −4.3 *** | 4.3 *** | −0.3 | 0.3 | ||
| Males | Percentage | 28.7% | 71.3% | 55.9% | 44.1% | 37.5% | 62.5% | <0.001 |
| (n = 27) | (n = 67) | (n = 80) | (n = 63) | (n = 45) | (n = 75) | |||
| Adj. residuals | −3.2 ** | 3.2 ** | 4.2 *** | −4.2 *** | −1.4 | 1.4 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Király, K.D.; Ladányi, M.; Fail, J. Reproductive Isolation in the Cryptic Species Complex of a Key Pest: Analysis of Mating and Rejection Behaviour of Onion Thrips (Thrips tabaci Lindeman). Biology 2022, 11, 396. https://doi.org/10.3390/biology11030396
Király KD, Ladányi M, Fail J. Reproductive Isolation in the Cryptic Species Complex of a Key Pest: Analysis of Mating and Rejection Behaviour of Onion Thrips (Thrips tabaci Lindeman). Biology. 2022; 11(3):396. https://doi.org/10.3390/biology11030396
Chicago/Turabian StyleKirály, Kristóf Domonkos, Márta Ladányi, and József Fail. 2022. "Reproductive Isolation in the Cryptic Species Complex of a Key Pest: Analysis of Mating and Rejection Behaviour of Onion Thrips (Thrips tabaci Lindeman)" Biology 11, no. 3: 396. https://doi.org/10.3390/biology11030396
APA StyleKirály, K. D., Ladányi, M., & Fail, J. (2022). Reproductive Isolation in the Cryptic Species Complex of a Key Pest: Analysis of Mating and Rejection Behaviour of Onion Thrips (Thrips tabaci Lindeman). Biology, 11(3), 396. https://doi.org/10.3390/biology11030396