The Problem of Effort Distribution in Heavy Glycolytic Trials with Special Reference to the 400 m Dash in Track and Field


Abstract
Simple Summary
Abstract
1. Introduction
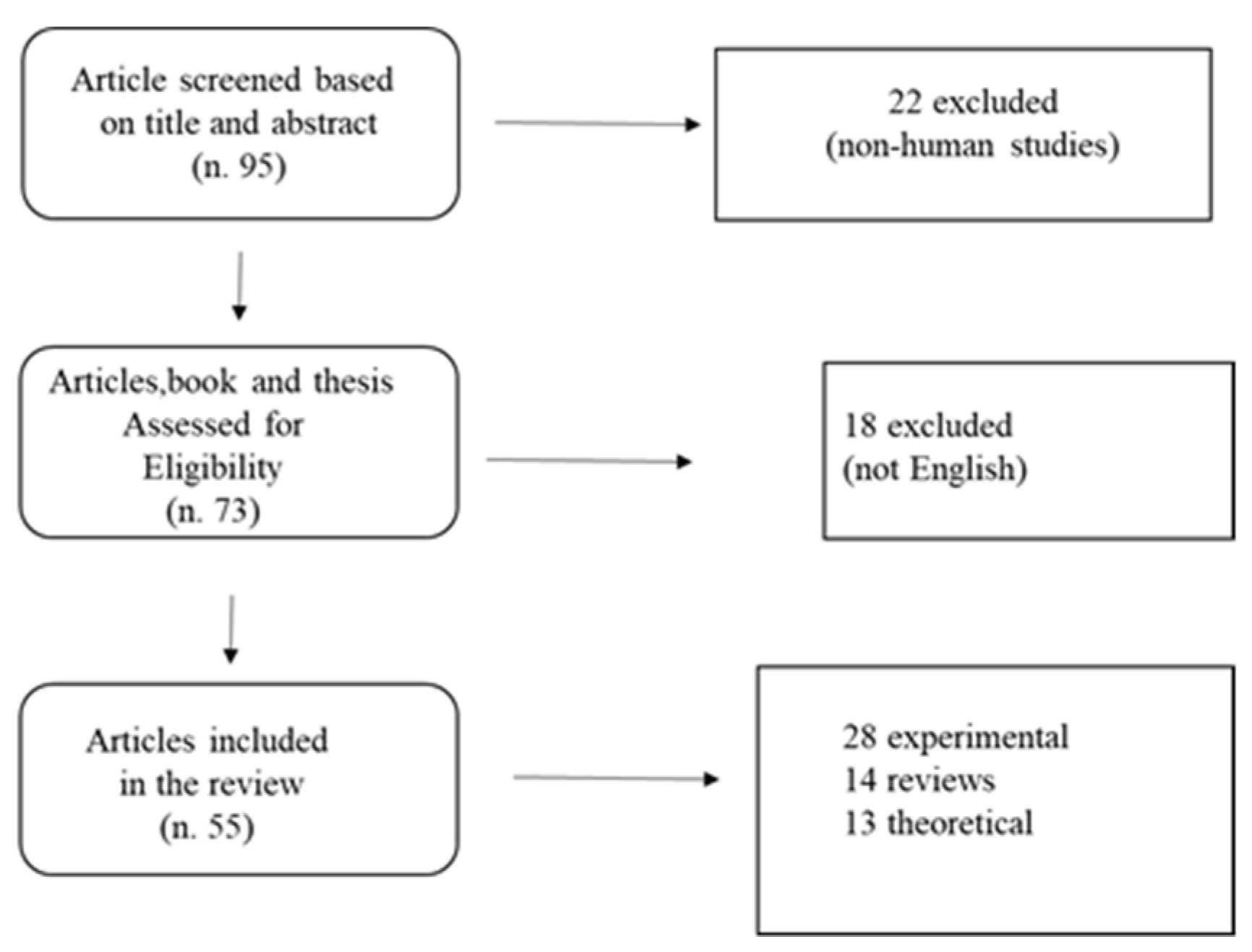
2. Materials and Methods
3. Results
3.1. Acidosis (pH), Lactic Acid (LA) and Limitations to Anaerobic Glycolytic Performance
3.2. Pacing
3.3. Teleanticipation
3.4. Brain and Perception of Effort
4. Discussion
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hill, A.V.; Lupton, H. Muscular Exercise, Lactic Acid, and the Supply and Utilization of Oxygen. QJM 1923, 16, 135–171. [Google Scholar] [CrossRef]
- Jones, J.H.; Lindstedt, S.L. Limits to Maximal Performance. Annu. Rev. Physiol. 1993, 55, 547–569. [Google Scholar] [CrossRef] [PubMed]
- Beckmann, J.; Ehmann, M.; Kossak, T.; Perl, B.; Hähl, W. Volition in Sport. Z. Sportpsychol. 2021, 28, 84–96. [Google Scholar] [CrossRef]
- Wolff, W.; Englert, C. On the Past, Present and Future of Volition Research in Sports. Z. Sportpsychol. 2021, 28, 81–83. [Google Scholar] [CrossRef]
- Bret, C.; Lacour, J.R.; Bourdin, M.; Locatelli, E.; De Angelis, M.; Faina, M.; Rahmani, A.; Messonnier, L. Differences in lactate exchange and removal abilities between high-level African and Caucasian 400-m track runners. Eur. J. Appl. Physiol. 2013, 113, 1489–1498. [Google Scholar] [CrossRef]
- Costill, D.L.; Barnett, A.; Sharp, R.; Fink, W.J.; Katz, A. Leg muscle pH following sprint running. Med. Sci. Sports Exerc. 1983, 15, 325–329. [Google Scholar] [CrossRef]
- Allen, D.G.; Lamb, G.D.; Westerblad, H. Skeletal muscle fatigue: Cellular mechanisms. Physiol. Rev. 2008, 88, 287–332. [Google Scholar] [CrossRef]
- van Hall, G. Lactate kinetics in human tissues at rest and during exercise. Acta Physiol. 2010, 199, 499–508. [Google Scholar] [CrossRef]
- Nummela, A.; Vuorimaa, T.; Rusko, H. Changes in force production, blood lactate and EMG activity in the 400-m sprint. J. Sports Sci. 1992, 10, 217–228. [Google Scholar] [CrossRef]
- Casado, A.; Hanley, B.; Jiménez-Reyes, P.; Renfree, A. Pacing profiles and tactical behaviors of elite runners. J. Sport Health Sci. 2021, 10, 537–549. [Google Scholar] [CrossRef]
- Keller, J.B. A Theory of Competitive Running. Phys. Today 1973, 26, 42–47. [Google Scholar] [CrossRef]
- Reardon, J.C. Optimal Pacing for Running 400 m and 800 m Track Races. Available online: https://arxiv.org/abs/1204.0313 (accessed on 5 October 2021).
- Martin-Acero, R.; Rodriguez, F.A.; Codina-Trenzano, A.; Jimenez-Reyes, P. Model for individual pacing strategies in the 400 metres. New Stud. Athl. 2017, 32, 27–44. [Google Scholar]
- Vittori, C. The development and training of young 400 m runners. New Stud. Athl. 1991, 6, 35–46. [Google Scholar]
- Saraslanidis, P.J.; Panoutsakopoulos, V.; Tsalis, G.A.; Kyprianou, E. The effect of different first 200-m pacing strategies on blood lactate and biomechanical parameters of the 400-m sprint. Eur. J. Appl. Physiol. 2011, 111, 1579–1590. [Google Scholar] [CrossRef] [PubMed]
- Coppenolle, H. Analysis of 200 m intermediate times for 400 m world class runners. Track Field Quaterly Rev. 1980, 80, 37–39. [Google Scholar]
- Wyatt, R.; Gunby, P. Optimal Pacing of 400 m and 800 m Races; A Standard Microeconomics Approach. Working Paper 12/2016. Available online: https://ir.canterbury.ac.nz/handle/10092/15227 (accessed on 12 November 2021).
- Hanon, C.; Claire Thomas, C. Effects of optimal pacing strategies for 400-, 800-, and 1500-m races on the V_O2 response. J. Sports Sci. Taylor Fr. SSH J. 2011, 29, 905–912. [Google Scholar] [CrossRef]
- Pollitt, L.; Walker, J.; Tucker, C.; Bissas, A. Biomechanical Report for the IAAF World Championships in London. 400 m Men’s. Available online: https://www.worldathletics.org/about-iaaf/documents/research-centre (accessed on 12 November 2021).
- German Athletic Federation. Biomechanical Analysis of Selected Events at 12 IAAF World Championship, Berlin 2009. Available online: https://www.bing.com/search?q=20.%09German+Athletic+Federation.+Biomechanical+analysis+of+selected+events+at+12+IAAF++World+Championship%2C+Berlin+2009.+&form=ANNTH1&refig=18647e74c80d4e018140b30978b713da (accessed on 12 November 2021).
- Hanon, C.; Lepretre, P.-M.; Bishop, D.; Thomas, C. Oxygen uptake and blood metabolic responses to a 400-m run. Eur. J. Appl. Physiol. 2010, 109, 233–240. [Google Scholar] [CrossRef]
- van Ingen Schenau, G.J.; de Koning, J.J.; de Groot, G. The distribution of anaerobic energy in 1000 and 4000 m cycling bouts. Int. J. Sports Med. 1992, 13, 447–451. [Google Scholar] [CrossRef]
- Yamamoto, K.; Miyashiro, K.; Naito, H.; Kigoshi, K.; Tanigawa, S.; Byun, K.O.; Miyashita, K.; Ogata, M. The relationship between race pattern and performance in the men’s 400-m sprint. Taiikugaku Kenkyu Jpn. J. Phys. Educ. Health Sport Sci. 2014, 59, 159–173. [Google Scholar] [CrossRef][Green Version]
- Tucker, R.; Noakes, T.D. The physiological regulation of pacing strategy during exercise: A critical review. Br. J. Sports Med. 2009, 43, e1. [Google Scholar] [CrossRef]
- Ulmer, H.V. Concept of an extracellular regulation of muscular-metabolic rate during heavy exercise in humans by psychophysiologial feedback. Experimentia 1996, 52, 416–420. [Google Scholar] [CrossRef] [PubMed]
- Edwards, A.M.; Bentley, M.B.; Mann, M.E.; Seaholme, T.S. Self-pacing in interval training: A teleoanticipatory approach. Psychophysiology 2011, 48, 136–141. [Google Scholar] [CrossRef] [PubMed]
- Ulmer, H.V.; Schulze, H. Zeitbezogene Zielantizipation als Teil motorischen Lernens bei 1- bis 10-miniitiger Haltearbeit. In Motodiagnostik-Mototherapie; Scholle, H.-C., Struppler, A., Freund, H.-J., Hefder, H., Schumann, N.P., Eds.; Universitgtsverlag Jena: Jena, Germany, 1995; pp. 311–315. [Google Scholar]
- Noakes, T.D. Is it Time to Retire the A.V. Hill Model? Sports Med. 2011, 41, 263–277. [Google Scholar] [CrossRef] [PubMed]
- Shephard, R.J. Is it time to retire the ‘central governor’? Sports Med. 2009, 39, 709–721. [Google Scholar] [CrossRef]
- Marino, F.E. If only I were paramecium too! A case for the complex, intelligent system of anticipatory regulation in fatigue. Fatigue Biomed. Health Behav. 2014, 2, 185–201. [Google Scholar] [CrossRef]
- Noakes, T.D.; Peltonen, J.E.; Rusko, H.K. Evidence that a central governor regulates exercise performance during acute hypoxia and hyperoxia. J. Exp. Biol. 2001, 204, 3225–3234. [Google Scholar] [CrossRef]
- Marcora, S. Do we really need a central governor to explain brain regulation of exercise performance? Eur. J. Appl. Physiol. 2008, 104, 929–931. [Google Scholar] [CrossRef]
- Åstrand, P.O.; Rodahl, K.; Dahl, H.A.; Strømme, S.B. Textbook of Work Physiology: Physiological Bases of Exercise. Human kinetics: Champaign, IL, USA, 2003; p. 628. [Google Scholar]
- Edwards, A.; Polman, R. Pacing and awareness: Brain regulation of physical activity. Sports Med. 2013, 43, 1057–1064. [Google Scholar] [CrossRef]
- Smits, B.L.; Pepping, G.J.; Hettinga, F.J. Pacing and decision making in sport and exercise: The roles of perception and action in the regulation of exercise intensity. Sports Med. 2014, 44, 763–775. [Google Scholar] [CrossRef]
- Ansley, L.; Robson, P.J.; St Clair, G.A.; Noakes Timothy, D. Evidence for anticipatory pacing strategies during supramaximal exercise lasting more than 30 s. Med. Sci. Sports Exerc. 2004, 36, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Poole, D.C.; Rossiter, H.B.; Brooks, G.A.; Gladden, L.B. The anaerobic threshold: 50+ years of controversy. J. Physiol. 2021, 599, 737–767. [Google Scholar] [CrossRef] [PubMed]
- Demello, J.J.; Cureton, K.J.; Boineau, R.E.; Singh, M.M. Ratings of perceived exertion at the lactate threshold in trained and untrained men and women. Med. Sci. Sports Exerc. 1987, 19, 354–362. [Google Scholar] [CrossRef] [PubMed]
- Matsui, T. Exhaustive endurance exercise activates brain glycogen breakdown and lactate production more than insulin-induced hypoglycemia. Am. J. Physiol. Integr. Comp. Physiol. 2021, 320, R500–R507. [Google Scholar] [CrossRef]
- Matsui, T.; Omuro, H.; Liu, Y.F.; Soya, M.; Shima, T.; McEwen, B.S.; Soya, H. Astrocytic glycogen-derived lactate fuels the brain during exhaustive exercise to maintain endurance capacity. Proc. Natl. Acad. Sci. USA 2017, 114, 6358–6363. [Google Scholar] [CrossRef]
- Faulkner, J.; Parfitt, G.; Eston, R. The rating of perceived exertion during competitive running scales with time. Psychophysiology 2008, 45, 977–985. [Google Scholar] [CrossRef]
- Eston, R.; Stansfield, R.; Westoby, P.; Parfitt, G. Effect of deception and expected exercise duration on psychological and physiological variables during treadmill running and cycling. Psychophysiology 2012, 49, 462–469. [Google Scholar] [CrossRef]
- Borg, G.A.V. Psychophysical bases of perceived exertion. Med. Sci. Sports Exerc. 1982, 14, 377–381. [Google Scholar] [CrossRef]
- Cafarelli, E.; Cain, W.S.; Stevens, J.C. Effort of Dynamic Exercise: Influence of Load, Duration, and Task. Ergonomics 1977, 20, 147–158. [Google Scholar] [CrossRef]
- Hamilton, A.R.; Behm, D.G. The effect of prior knowledge of test endpoint on non-local muscle fatigue. Eur. J. Appl. Physiol. 2017, 117, 651–663. [Google Scholar] [CrossRef]
- Wingfield, G.; Marino, F.; Skein, M. The influence of knowledge of performance endpoint on pacing strategies, perception of effort, and neural activity during 30-km cycling time trials. Physiol. Rep. 2018, 6, e13892. [Google Scholar] [CrossRef] [PubMed]
- Jones, H.S.; Williams, E.L.; Bridge, C.A.; Marchant, D.; Midgley, A.W.; Micklewright, D.; Mc Naughton, L.R. Physiological and psychological effects of deception on pacing strategy and performance: A review. Sports Med. 2013, 43, 1243–1257. [Google Scholar] [CrossRef] [PubMed]
- Seashore, R.H. Studies in motor rhythm. Psychol. Monogr. 1926, 36, 142–189. [Google Scholar] [CrossRef]
- Louie, A. Robert Rosen’s anticipatory systems. Foresight—The journal of future studies strategic thinking and policy. Foresight 2010, 12, 18–29. [Google Scholar] [CrossRef]
- Rosen, R. Anticipatory Systems: Philosophical, Mathematical & Methodological Foundations; Pergamon Press: Oxford, UK, 1985. [Google Scholar]
- Knicker, A.J.; Renshaw, I.; Oldham, A.R.; Cairns, S.P. Interactive processes link the multiple symptoms of fatigue in sport competition. Sports Med. 2011, 41, 307–328. [Google Scholar] [CrossRef]
- Hettinga, F.J.; Konings, M.J.; Pepping, G.J. The Science of Racing against Opponents: Affordance Competition and the Regulation of Exercise Intensity in Head-to-Head Competition. Front. Physiol. 2017, 28, 118. [Google Scholar] [CrossRef]
- Konings, M.J.; Hettinga, F.J. Pacing Decision Making in Sport and the Effects of Interpersonal Competition: A Critical Review. Sports Med. 2018, 48, 1829–1843. [Google Scholar] [CrossRef]
- Konings, M.J.; Hettinga, F.J. Objectifying Tactics: Athlete and Race Variability in Elite Short-Track Speed Skating. Int. J. Sports Physiol. Perform. 2018, 13, 170–175. [Google Scholar] [CrossRef]
- Williams, E.L.; Jones, H.S.; Sparks, S.A.; Midgley, A.W.; Marchant, D.C.; Bridge, C.A.; Mcnaughton, L.R. Altered Psychological Responses to Different Magnitudes of Deception during Cycling. Med. Sci. Sports Exerc. 2015, 47, 2423–2430. [Google Scholar] [CrossRef]
- Konings, M.J.; Parkinson, J.; Micklewright, D.; Zijdewind, I.; Hettinga, F.J. Willingness to Tolerate Higher Levels of Peripheral Fatigue Might Explain the Performance Improvement When Riding Against Opponents. In Vienna: Book of Abstract European College of Sport Sciences; European College of Sport Science: Cologne, Germany, 2016. [Google Scholar]


{kind=link}
{kind=link}
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cicchella, A. The Problem of Effort Distribution in Heavy Glycolytic Trials with Special Reference to the 400 m Dash in Track and Field. Biology 2022, 11, 216. https://doi.org/10.3390/biology11020216
Cicchella A. The Problem of Effort Distribution in Heavy Glycolytic Trials with Special Reference to the 400 m Dash in Track and Field. Biology. 2022; 11(2):216. https://doi.org/10.3390/biology11020216
Chicago/Turabian StyleCicchella, Antonio. 2022. "The Problem of Effort Distribution in Heavy Glycolytic Trials with Special Reference to the 400 m Dash in Track and Field" Biology 11, no. 2: 216. https://doi.org/10.3390/biology11020216
APA StyleCicchella, A. (2022). The Problem of Effort Distribution in Heavy Glycolytic Trials with Special Reference to the 400 m Dash in Track and Field. Biology, 11(2), 216. https://doi.org/10.3390/biology11020216