
Plant Microbiome Engineering: Hopes or Hypes

 ,
,  ,
,  ,
,  , , ,
, , ,  ,
, .jpg) , and
, and {kind=link}
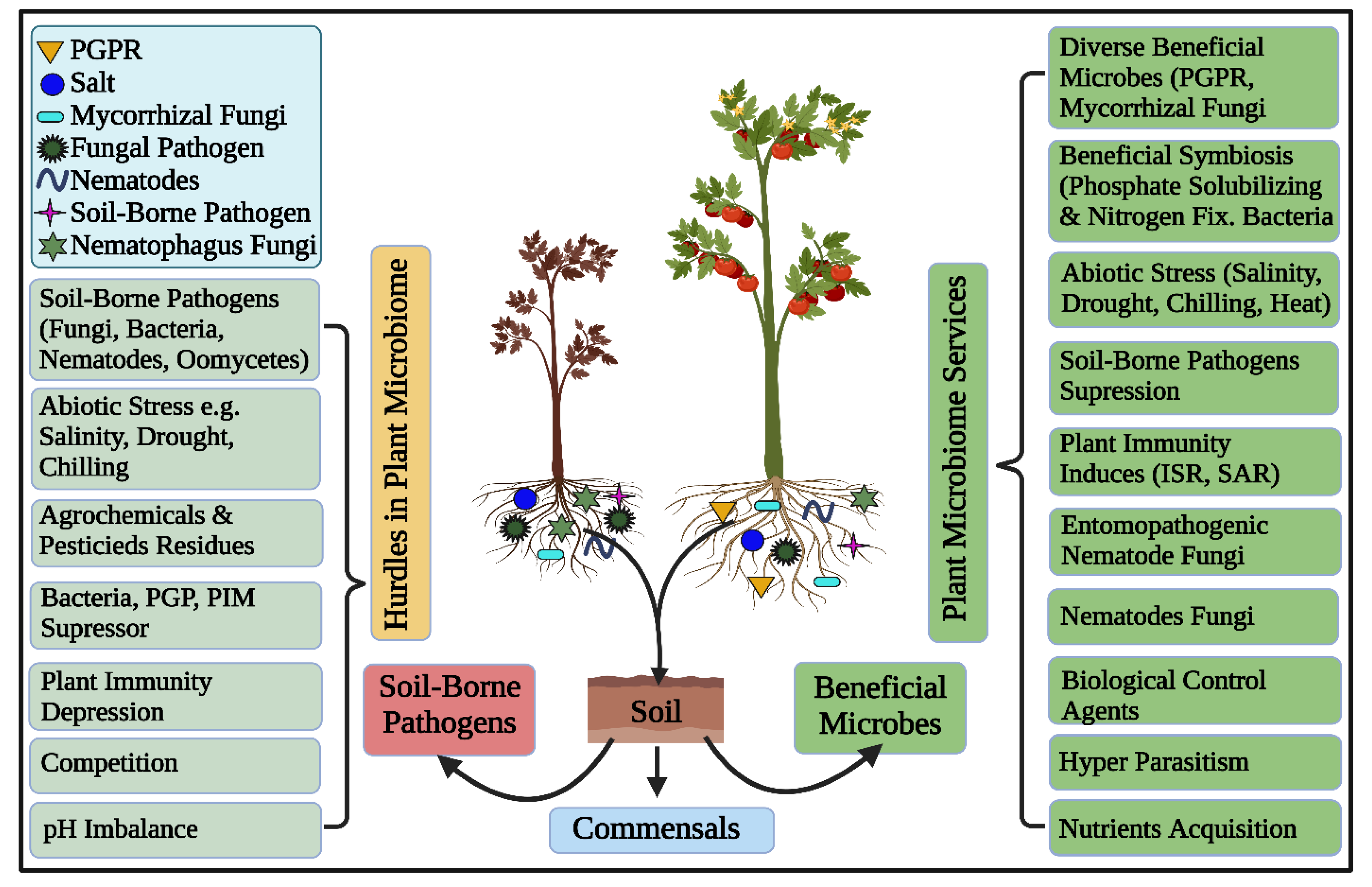{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Plant Microbiome: Composition, Complexity and Diversity
3. The Dilemma of Plant Microbiome Interactions
4. Deciphering the Potential Role of Micro-Macro Fauna in Plant Microbiome Engineering
5. Omics Tools: Putative Role and Limitations in Plant Microbiome Interactions
6. Disease Suppressive Soil: Underlying Mechanism and Manipulation for Healthy Microbiome Engineering
7. Sustainable Approaches
8. Biological Fertilization
9. Conclusions and Future Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thakur, A.; Singh, S.; Dulta, K.; Singh, N.; Ali, B.; Hafeez, A.; Vodnar, D.C.; Marc, R.A. Nutritional evaluation, phytochemical makeup, and antibacterial and antioxidant properties of wild plants utilized as food by the Gaddis, a tribe in the Western Himalayas. Front. Agron. 2022, 4, 1010309. [Google Scholar] [CrossRef]
- Leister, D. Retrograde Signaling in Plants: From Simple to Complex Scenarios. Front. Plant Sci. 2012, 3, 135. [Google Scholar] [CrossRef] [PubMed]
- Dola, D.B.; Mannan, M.A.; Sarker, U.; Al Mamun, M.A.; Islam, T.; Ercisli, S.; Saleem, M.H.; Ali, B.; Pop, O.L.; Marc, R.A. Nano-Iron Oxide Accelerates Growth, Yield, and Quality of Glycine Max Seed in Water Deficits. Front. Plant Sci. 2022, 13, 992535. [Google Scholar] [CrossRef] [PubMed]
- Ikinci, A.; Bolat, I.; Ercisli, S.; Kodad, O. Influence of Rootstocks On Growth, Yield, Fruit Quality And Leaf Mineral Element Contents of Pear cv. ‘Santa Maria’ in Semi-Arid Conditions. Biol. Res. 2014, 47, 71. [Google Scholar] [CrossRef]
- Hussain, S.S.; Rasheed, M.; Saleem, M.H.; Ahmed, Z.I.; Hafeez, A.; Jilani, G.; Alamri, S.; Hashem, M.; Ali, S. Salt Tolerance In Maize With Melatonin Priming To Achieve Sustainability In Yield In Salt Affected Soils. Pak. J. Bot. 2022, 55, 1. [Google Scholar] [CrossRef] [PubMed]
- Amna Ali, B.; Azeem, M.A.; Qayyum, A.; Mustafa, G.; Ahmad, M.A.; Javed, M.T.; Chaudhary, H.J. Bio-Fabricated Silver Nanoparticles: A Sustainable Approach for Augmentation of Plant Growth and Pathogen Control. In Sustainable Agriculture Reviews 53; Springer: Berlin/Heidelberg, Germany, 2021; pp. 345–371. [Google Scholar]
- Bibi, S.; Ullah, S.; Hafeez, A.; Khan, M.N.; Javed, M.A.; Ali, B.; Din, I.U.; Bangash, S.A.K.; Wahab, S.; Wahid, N. Exogenous Ca/Mg Quotient Reduces the Inhibitory Effects of PEG Induced Osmotic Stress on Avena sativa L. Braz. J. Biol. 2022, 84, e264642. [Google Scholar] [CrossRef]
- Naz, R.; Khan, M.S.; Hafeez, A.; Fazil, M.; Khan, M.N.; Ali, B.; Javed, M.A.; Imran, M.; Shati, A.A.; Alfaifi, M.Y. Assessment of Phytoremediation Potential of Native Plant Species Naturally Growing in a Heavy Metal-Polluted Industrial Soils. Braz. J. Biol. 2022, 84, e264473. [Google Scholar] [CrossRef]
- Nawaz, H.; Ali, A.; Saleem, M.H.; Ameer, A.; Hafeez, A.; Alharbi, K.; Ezzat, A.; Khan, A.; Jamil, M.; Farid, G. Comparative Effectiveness of EDTA and Citric Acid Assisted Phytoremediation of Ni Contaminated Soil by Using Canola (Brassica napus). Braz. J. Biol. 2022, 82, e261785. [Google Scholar] [CrossRef]
- Zainab, N.; Din, B.U.; Javed, M.T.; Afridi, M.S.; Mukhtar, T.; Kamran, M.A.; Khan, A.A.; Ali, J.; Jatoi, W.N.; Munis, M.F.H. Deciphering Metal Toxicity Responses of Flax (Linum usitatissimum L.) with Exopolysaccharide and ACC-Deaminase Producing Bacteria in Industrially Contaminated Soils. Plant Physiol. Biochem. 2020, 152, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Ali, J.; Mahmood, T.; Hayat, K.; Afridi, M.S.; Ali, F.; Chaudhary, H.J. Phytoextraction of Cr by Maize (Zea mays L.): The Role of Plant Growth Promoting Endophyte and Citric Acid under Polluted Soil. Arch. Environ. Prot. 2018, 44, 73–82. [Google Scholar]
- Saeed, S.; Ullah, A.; Ullah, S.; Noor, J.; Ali, B.; Khan, M.N.; Hashem, M.; Mostafa, Y.S.; Alamri, S. Validating the Impact of Water Potential and Temperature on Seed Germination of Wheat (Triticum aestivum L.) via Hydrothermal Time Model. Life 2022, 12, 983. [Google Scholar] [CrossRef]
- Mehmood, S.; Khatoon, Z.; Amna; Ahmad, I.; Muneer, M.A.; Kamran, M.A.; Ali, J.; Ali, B.; Chaudhary, H.J.; Munis, M.F.H. Bacillus sp. PM31 Harboring Various Plant Growth-Promoting Activities Regulates Fusarium Dry Rot and Wilt Tolerance in Potato. Arch. Agron. Soil Sci. 2021, 1–15. [Google Scholar] [CrossRef]
- Faryal, S.; Ullah, R.; Khan, M.N.; Ali, B.; Hafeez, A.; Jaremko, M.; Qureshi, K.A. Thiourea-Capped Nanoapatites Amplify Osmotic Stress Tolerance in Zea Mays L. by Conserving Photosynthetic Pigments, Osmolytes Biosynthesis and Antioxidant Biosystems. Molecules 2022, 27, 5744. [Google Scholar] [CrossRef] [PubMed]
- Váry, Z.; Mullins, E.; McElwain, J.C.; Doohan, F.M. The Severity of Wheat Diseases Increases When Plants and Pathogens Are Acclimatized to Elevated Carbon Dioxide. Glob. Change Biol. 2015, 21, 2661–2669. [Google Scholar] [CrossRef]
- Trecate, L.; Sedlakova, B.; Mieslerova, B.; Manstretta, V.; Rossi, V.; Lebeda, A. Effect of Temperature on Infection and Development of Powdery Mildew on Cucumber. Plant Pathol. 2019, 68, 1165–1178. [Google Scholar] [CrossRef]
- Magan, N.; Medina, A.; Aldred, D. Possible Climate-Change Effects on Mycotoxin Contamination of Food Crops Pre-and Postharvest. Plant Pathol. 2011, 60, 150–163. [Google Scholar] [CrossRef]
- Al-Zaban, M.I.; Alhag, S.K.; Dablool, A.S.; Ahmed, A.E.; Alghamdi, S.; Ali, B.; Al-Saeed, F.A.; Saleem, M.H.; Poczai, P. Manufactured Nano-Objects Confer Viral Protection against Cucurbit Chlorotic Yellows Virus (CCYV) Infecting Nicotiana Benthamiana. Microorganisms 2022, 10, 1837. [Google Scholar] [CrossRef] [PubMed]
- Metayi, M.H.; El-Naby, A.; Shimaa, S.; El-Habal, N.A.; Fahmy, H.H.; Abdou, M.S.; Ali, B.; Abdel-Rheim, K.H.; Abdel-Megeed, A. Omani Frankincense Nanoemulsion Formulation Efficacy and Its Latent Effects on Biological Aspects of the Spiny Bollworm Earias insulana (Boisd.). Front. Physiol. 2022, 13, 1001136. [Google Scholar] [CrossRef] [PubMed]
- Salam, A.; Afridi, M.S.; Javed, M.A.; Saleem, A.; Hafeez, A.; Khan, A.R.; Zeeshan, M.; Ali, B.; Azhar, W.; Sumaira; et al. Nano-Priming against Abiotic Stress: A Way Forward towards Sustainable Agriculture. Sustainability 2022, 14, 14880. [Google Scholar] [CrossRef]
- Dean, R.; Van Kan, J.A.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J. The Top 10 Fungal Pathogens in Molecular Plant Pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef]
- Egamberdieva, D.; Kamilova, F.; Validov, S.; Gafurova, L.; Kucharova, Z.; Lugtenberg, B. High Incidence of Plant Growth-Stimulating Bacteria Associated with the Rhizosphere of Wheat Grown on Salinated Soil in Uzbekistan. Environ. Microbiol. 2008, 10, 1–9. [Google Scholar] [CrossRef]
- Glick, B.R.; Gamalero, E. Recent Developments in the Study of Plant Microbiomes. Microorganisms 2021, 9, 1533. [Google Scholar] [CrossRef] [PubMed]
- Sheth, R.U.; Cabral, V.; Chen, S.P.; Wang, H.H. Manipulating Bacterial Communities by in Situ Microbiome Engineering. Trends Genet. 2016, 32, 189–200. [Google Scholar] [CrossRef]
- Pozo, M.J.; Zabalgogeazcoa, I.; de Aldana, B.R.V.; Martinez-Medina, A. Untapping the Potential of Plant Mycobiomes for Applications in Agriculture. Curr. Opin. Plant Biol. 2021, 60, 102034. [Google Scholar] [CrossRef]
- Orgiazzi, A.; Bardgett, R.D.; Barrios, E. Global Soil Biodiversity Atlas; European Commission: Brussels, Belgium, 2016. [Google Scholar]
- Sokol, N.W.; Slessarev, E.; Marschmann, G.L.; Nicolas, A.; Blazewicz, S.J.; Brodie, E.L.; Firestone, M.K.; Foley, M.M.; Hestrin, R.; Hungate, B.A. Life and Death in the Soil Microbiome: How Ecological Processes Influence Biogeochemistry. Nat. Rev. Microbiol. 2022, 20, 415–430. [Google Scholar] [CrossRef]
- Jain, A.; Das, S. Insight into the Interaction between Plants and Associated Fluorescent Pseudomonas spp. Int. J. Agron. 2016, 2016, 4269010. [Google Scholar] [CrossRef]
- Fahad, S.; Chavan, S.B.; Chichaghare, A.R.; Uthappa, A.R.; Kumar, M.; Kakade, V.; Pradhan, A.; Jinger, D.; Rawale, G.; Yadav, D.K.; et al. Agroforestry Systems for Soil Health Improvement and Maintenance. Sustainability 2022, 14, 14877. [Google Scholar] [CrossRef]
- Mendes, R.; Garbeva, P.; Raaijmakers, J.M. The Rhizosphere Microbiome: Significance of Plant Beneficial, Plant Pathogenic, and Human Pathogenic Microorganisms. FEMS Microbiol. Rev. 2013, 37, 634–663. [Google Scholar] [CrossRef] [PubMed]
- Alexander, M. Introduction to Soil Microbiology, 2nd ed.; John Wiley & Sons: New York, NY, USA, 1977. [Google Scholar]
- Brady, N.C.; Weil, R.R.; Weil, R.R. The Nature and Properties of Soils; Prentice Hall Upper Saddle River: Hoboken, NJ, USA, 2008; Volume 13. [Google Scholar]
- Lynch, J.M. The Terrestrial Environment. In Microbial Ecology: A Conceptual Approach; Wiley: Hoboken, NJ, USA, 1979; pp. 9–67. [Google Scholar]
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The global burden of pathogens and pests on major food crops. Nat. Ecol. Evol. 2019, 3, 430–439. [Google Scholar] [CrossRef]
- Vannier, N.; Agler, M.; Hacquard, S. Microbiota-Mediated Disease Resistance in Plants. PLoS Pathog. 2019, 15, e1007740. [Google Scholar] [CrossRef] [PubMed]
- Finkel, O.M.; Castrillo, G.; Herrera Paredes, S.; Salas González, I.; Dangl, J.L. Understanding and Exploiting Plant Beneficial Microbes. Curr. Opin. Plant Biol. 2017, 38, 155–163. [Google Scholar] [CrossRef]
- Afridi, M.S.; Fakhar, A.; Kumar, A.; Ali, S.; Medeiros, F.H.; Muneer, M.A.; Ali, H.; Saleem, M. Harnessing Microbial Multitrophic Interactions for Rhizosphere Microbiome Engineering. Microbiol. Res. 2022, 265, 127199. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Bai, Z.; Hoefel, D.; Wang, X.; Zhang, L.; Li, Z. Microbial Diversity within the Phyllosphere of Different Vegetable Species. Curr. Res. Top. Appl. Microbiol. Microb. Biotechnol. 2010, 2, 1067–1077. [Google Scholar]
- Ali, M.A.; Naveed, M.; Mustafa, A.; Abbas, A. The Good, the Bad, and the Ugly of Rhizosphere Microbiome. In Probiotics and Plant Health; Kumar, V., Kumar, M., Sharma, S., Prasad, R., Eds.; Springer: Singapore, 2017; pp. 253–290. ISBN 978-981-10-3473-2. [Google Scholar]
- Albright, M.B.; Louca, S.; Winkler, D.E.; Feeser, K.L.; Haig, S.-J.; Whiteson, K.L.; Emerson, J.B.; Dunbar, J. Solutions in Microbiome Engineering: Prioritizing Barriers to Organism Establishment. ISME J. 2022, 16, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Lau, S.-E.; Teo, W.F.A.; Teoh, E.Y.; Tan, B.C. Microbiome Engineering and Plant Biostimulants for Sustainable Crop Improvement and Mitigation of Biotic and Abiotic Stresses. Discov. Food 2022, 2, 9. [Google Scholar] [CrossRef]
- Haider, M.W.; Nafees, M.; Ahmad, I.; Ali, B.; Iqbal, R.; Vodnar, D.C.; Marc, R.A.; Kamran, M.; Saleem, M.H.; Al-Ghamdi, A.A. Postharvest Dormancy-Related Changes of Endogenous Hormones in Relation to Different Dormancy-Breaking Methods of Potato (Solanum tuberosum L.) Tubers. Front. Plant Sci. 2022, 13, 945256. [Google Scholar] [CrossRef]
- Morales-Cedeño, L.R.; del Carmen Orozco-Mosqueda, M.; Loeza-Lara, P.D.; Parra-Cota, F.I.; de Los Santos-Villalobos, S.; Santoyo, G. Plant Growth-Promoting Bacterial Endophytes as Biocontrol Agents of Pre-and Post-Harvest Diseases: Fundamentals, Methods of Application and Future Perspectives. Microbiol. Res. 2021, 242, 126612. [Google Scholar] [CrossRef]
- Ali, B.; Hafeez, A.; Javed, M.A.; Afridi, M.S.; Abbasi, H.A.; Qayyum, A.; Batool, T.; Ullah, A.; Marc, R.A.; Al Jaouni, S.K. Role of Endophytic Bacteria in Salinity Stress Amelioration by Physiological and Molecular Mechanisms of Defense: A Comprehensive Review. South Afr. J. Bot. 2022, 151, 33–46. [Google Scholar] [CrossRef]
- Biessy, A.; Novinscak, A.; Blom, J.; Léger, G.; Thomashow, L.S.; Cazorla, F.M.; Josic, D.; Filion, M. Diversity of Phytobeneficial Traits Revealed by Whole-Genome Analysis of Worldwide-Isolated Phenazine-Producing Pseudomonas spp. Environ. Microbiol. 2019, 21, 437–455. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, J.; Yang, N.; Wen, Z.; Sun, X.; Chai, Y.; Ma, Z. Wheat Microbiome Bacteria Can Reduce Virulence of a Plant Pathogenic Fungus by Altering Histone Acetylation. Nat. Commun. 2018, 9, 3429. [Google Scholar] [CrossRef]
- Chin-A-Woeng, T.F.C.; Bloemberg, G.V.; van der Bij, A.J.; van der Drift, K.M.G.M.; Schripsema, J.; Kroon, B.; Scheffer, R.J.; Keel, C.; Bakker, P.A.H.M.; Tichy, H.-V.; et al. Biocontrol by Phenazine-1-Carboxamide-Producing Pseudomonas chlororaphis PCL1391 of Tomato Root Rot Caused by Fusarium oxysporum f. sp. Radicis-Lycopersici. MPMI 1998, 11, 1069–1077. [Google Scholar] [CrossRef]
- Jones, P.; Garcia, B.J.; Furches, A.; Tuskan, G.A.; Jacobson, D. Plant Host-Associated Mechanisms for Microbial Selection. Front. Plant Sci. 2019, 10, 862. [Google Scholar] [CrossRef] [PubMed]
- Zipfel, C.; Oldroyd, G.E.D. Plant Signalling in Symbiosis and Immunity. Nature 2017, 543, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Coaker, G.; Zhou, J.-M.; Dong, X. Plant Immune Mechanisms: From Reductionistic to Holistic Points of View. Mol. Plant 2020, 13, 1358–1378. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, P.J.P.; Colaianni, N.R.; Fitzpatrick, C.R.; Dangl, J.L. Beyond Pathogens: Microbiota Interactions with the Plant Immune System. Curr. Opin. Microbiol. 2019, 49, 7–17. [Google Scholar] [CrossRef]
- Köhl, J.; Kolnaar, R.; Ravensberg, W.J. Mode of Action of Microbial Biological Control Agents Against Plant Diseases: Relevance Beyond Efficacy. Front. Plant Sci. 2019, 10, 845. [Google Scholar] [CrossRef]
- Arif, I.; Batool, M.; Schenk, P.M. Plant Microbiome Engineering: Expected Benefits for Improved Crop Growth and Resilience. Trends Biotechnol. 2020, 38, 1385–1396. [Google Scholar] [CrossRef]
- Wassermann, B.; Cernava, T.; Müller, H.; Berg, C.; Berg, G. Seeds of Native Alpine Plants Host Unique Microbial Communities Embedded in Cross-Kingdom Networks. Microbiome 2019, 7, 108. [Google Scholar] [CrossRef]
- Liu, Z.; Beskrovnaya, P.; Melnyk, R.A.; Hossain, S.S.; Khorasani, S.; O’Sullivan, L.R.; Wiesmann, C.L.; Bush, J.; Richard, J.D.; Haney, C.H. A Genome-Wide Screen Identifies Genes in Rhizosphere-Associated Pseudomonas Required to Evade Plant Defenses. mBio 2018, 9, e00433-e18. [Google Scholar] [CrossRef]
- Snelders, N.C.; Rovenich, H.; Petti, G.C.; Rocafort, M.; van den Berg, G.C.M.; Vorholt, J.A.; Mesters, J.R.; Seidl, M.F.; Nijland, R.; Thomma, B.P.H.J. Microbiome Manipulation by a Soil-Borne Fungal Plant Pathogen Using Effector Proteins. Nat. Plants 2020, 6, 1365–1374. [Google Scholar] [CrossRef]
- Kamilova, F.; Lamers, G.; Lugtenberg, B. Biocontrol Strain Pseudomonas fluorescens WCS365 Inhibits Germination of Fusarium oxysporum Spores in Tomato Root Exudate as Well as Subsequent Formation of New Spores. Environ. Microbiol. 2008, 10, 2455–2461. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, V.B.; Akland, K.; Johnson, N.C.; Bowker, M.A. Do Soil Inoculants Accelerate Dryland Restoration? A Simultaneous Assessment of Biocrusts and Mycorrhizal Fungi. Restor. Ecol. 2020, 28, S115–S126. [Google Scholar] [CrossRef]
- Fonseca, E.d.S.; Peixoto, R.S.; Rosado, A.S.; Balieiro, F.d.C.; Tiedje, J.M.; Rachid, C.T.C.d.C. The Microbiome of Eucalyptus Roots under Different Management Conditions and Its Potential for Biological Nitrogen Fixation. Microb. Ecol. 2018, 75, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Valladares, F.; Gianoli, E.; Gómez, J.M. Ecological Limits to Plant Phenotypic Plasticity. New Phytol. 2007, 176, 749–763. [Google Scholar] [CrossRef] [PubMed]
- Tardieu, F. Plant Response to Environmental Conditions: Assessing Potential Production, Water Demand, and Negative Effects of Water Deficit. Front. Physiol. 2013, 4, 17. [Google Scholar] [CrossRef]
- Dombrowski, N.; Schlaeppi, K.; Agler, M.T.; Hacquard, S.; Kemen, E.; Garrido-Oter, R.; Wunder, J.; Coupland, G.; Schulze-Lefert, P. Root Microbiota Dynamics of Perennial Arabis Alpina Are Dependent on Soil Residence Time but Independent of Flowering Time. ISME J. 2017, 11, 43–55. [Google Scholar] [CrossRef]
- Peiffer, J.A.; Spor, A.; Koren, O.; Jin, Z.; Tringe, S.G.; Dangl, J.L.; Buckler, E.S.; Ley, R.E. Diversity and Heritability of the Maize Rhizosphere Microbiome under Field Conditions. Proc. Natl. Acad. Sci. USA 2013, 110, 6548–6553. [Google Scholar] [CrossRef]
- Lauber, C.L.; Hamady, M.; Knight, R.; Fierer, N. Pyrosequencing-Based Assessment of Soil PH as a Predictor of Soil Bacterial Community Structure at the Continental Scale. Appl. Environ. Microbiol. 2009, 75, 5111–5120. [Google Scholar] [CrossRef]
- Jolly, W.M.; Cochrane, M.A.; Freeborn, P.H.; Holden, Z.A.; Brown, T.J.; Williamson, G.J.; Bowman, D.M.J.S. Climate-Induced Variations in Global Wildfire Danger from 1979 to 2013. Nat. Commun. 2015, 6, 7537. [Google Scholar] [CrossRef]
- Tardif, S.; Yergeau, É.; Tremblay, J.; Legendre, P.; Whyte, L.G.; Greer, C.W. The Willow Microbiome Is Influenced by Soil Petroleum-Hydrocarbon Concentration with Plant Compartment-Specific Effects. Front. Microbiol. 2016, 7, 1363. [Google Scholar] [CrossRef]
- Fierer, N.; Jackson, R.B. The Diversity and Biogeography of Soil Bacterial Communities. Proc. Natl. Acad. Sci. USA 2006, 103, 626–631. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, R.I.; Thomson, B.C.; James, P.; Bell, T.; Bailey, M.; Whiteley, A.S. The Bacterial Biogeography of British Soils. Environ. Microbiol. 2011, 13, 1642–1654. [Google Scholar] [CrossRef] [PubMed]
- Ke, J.; Wang, B.; Yoshikuni, Y. Microbiome Engineering: Synthetic Biology of Plant-Associated Microbiomes in Sustainable Agriculture. Trends Biotechnol. 2021, 39, 244–261. [Google Scholar] [CrossRef] [PubMed]
- Kaul, S.; Choudhary, M.; Gupta, S.; Dhar, M.K. Engineering Host Microbiome for Crop Improvement and Sustainable Agriculture. Front. Microbiol. 2021, 12, 635917. [Google Scholar] [CrossRef]
- Ma, J.; Ali, S.; Saleem, M.H.; Mumtaz, S.; Yasin, G.; Ali, B.; Al-Ghamdi, A.A.; Elshikh, M.S.; Vodnar, D.C.; Marc, R.A.; et al. Short-Term Responses of Spinach (Spinacia oleracea L.) to the Individual and Combinatorial Effects of Nitrogen, Phosphorus and Potassium and Silicon in the Soil Contaminated by Boron. Front. Plant Sci. 2022, 13, 983156. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Saleem, M.H.; Ali, B.; Rasheed, R.; Ashraf, M.A.; Aziz, H.; Ercisli, S.; Riaz, S.; Elsharkawy, M.M.; Hussain, I.; et al. Impact of Foliar Application of Syringic Acid on Tomato (Solanum lycopersicum L.) under Heavy Metal Stress-Insights into Nutrient Uptake, Redox Homeostasis, Oxidative Stress, and Antioxidant Defense. Front. Plant Sci. 2022, 13, 950120. [Google Scholar] [CrossRef]
- Ma, J.; Saleem, M.H.; Yasin, G.; Mumtaz, S.; Qureshi, F.F.; Ali, B.; Ercisli, S.; Alhag, S.K.; Ahmed, A.E.; Vodnar, D.C.; et al. Individual and Combinatorial Effects of SNP and NaHS on Morpho-Physio-Biochemical Attributes and Phytoextraction of Chromium through Cr-Stressed Spinach (Spinacia oleracea L.). Front. Plant Sci. 2022, 13, 973740. [Google Scholar] [CrossRef]
- Saleem, K.; Asghar, M.A.; Saleem, M.H.; Raza, A.; Kocsy, G.; Iqbal, N.; Ali, B.; Albeshr, M.F.; Bhat, E.A. Chrysotile-Asbestos-Induced Damage in Panicum Virgatum and Phleum Pretense Species and Its Alleviation by Organic-Soil Amendment. Sustainability 2022, 14, 10824. [Google Scholar] [CrossRef]
- Din Umar, U.; Ahmed, N.; Zafar, M.Z.; Rehman, A.; Naqvi, S.A.H.; Zulfiqar, M.A.; Malik, M.T.; Ali, B.; Saleem, M.H.; Marc, R.A. Micronutrients Foliar and Drench Application Mitigate Mango Sudden Decline Disorder and Impact Fruit Yield. Agronomy 2022, 12, 2449. [Google Scholar] [CrossRef]
- Ali, S.; Ullah, S.; Khan, M.N.; Khan, W.M.; Razak, S.A.; Wahab, S.; Hafeez, A.; Khan Bangash, S.A.; Poczai, P. The Effects of Osmosis and Thermo-Priming on Salinity Stress Tolerance in Vigna radiata L. Sustainability 2022, 14, 12924. [Google Scholar] [CrossRef]
- Ali, B.; Wang, X.; Saleem, M.H.; Azeem, M.A.; Afridi, M.S.; Nadeem, M.; Ghazal, M.; Batool, T.; Qayyum, A.; Alatawi, A. Bacillus Mycoides PM35 Reinforces Photosynthetic Efficiency, Antioxidant Defense, Expression of Stress-Responsive Genes, and Ameliorates the Effects of Salinity Stress in Maize. Life 2022, 12, 219. [Google Scholar] [CrossRef]
- Zainab, N.; Khan, A.A.; Azeem, M.A.; Ali, B.; Wang, T.; Shi, F.; Alghanem, S.M.; Hussain Munis, M.F.; Hashem, M.; Alamri, S. PGPR-Mediated Plant Growth Attributes and Metal Extraction Ability of Sesbania Sesban L. in Industrially Contaminated Soils. Agronomy 2021, 11, 1820. [Google Scholar] [CrossRef]
- Khan, M.A.; Adnan, M.; Basir, A.; Fahad, S.; Hafeez, A.; Saleem, M.H.; Ahmad, M.; Gul, F.; Subhan, D.F.; Alamri, S.; et al. Impact of tillage and potassium levels and sources on growth, yield and yield attributes of wheat. Pak. J. Bot. 2022, 55, 30848. [Google Scholar] [CrossRef]
- Mukhtar, T.; Ali, F.; Rafique, M.; Ali, J.; Afridi, M.S.; Smith, D.; Mehmood, S.; Souleimanov, A.; Jellani, G.; Sultan, T. Biochemical Characterization and Potential of Bacillus Safensis Strain SCAL1 to Mitigate Heat Stress in Solanum lycopersicum L. J. Plant Growth Regul. 2022, 139, 569–577. [Google Scholar] [CrossRef]
- Cao, Y.; Pi, H.; Chandrangsu, P.; Li, Y.; Wang, Y.; Zhou, H.; Xiong, H.; Helmann, J.D.; Cai, Y. Antagonism of Two Plant-Growth Promoting Bacillus Velezensis Isolates against Ralstonia Solanacearum and Fusarium oxysporum. Sci. Rep. 2018, 8, 4360. [Google Scholar] [CrossRef]
- Chandrasekaran, M.; Chun, S.C. Expression of PR-Protein Genes and Induction of Defense-Related Enzymes by Bacillus Subtilis CBR05 in Tomato (Solanum lycopersicum) Plants Challenged with Erwinia carotovora subsp. carotovora. Biosci. Biotechnol. Biochem. 2016, 80, 2277–2283. [Google Scholar] [CrossRef]
- Vega, C.; Rodríguez, M.; Llamas, I.; Béjar, V.; Sampedro, I. Silencing of Phytopathogen Communication by the Halotolerant PGPR Staphylococcus Equorum Strain EN21. Microorganisms 2019, 8, 42. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Singh, D.; Gupta, A.; Pandey, K.D.; Singh, P.K.; Kumar, A. Plant Growth Promoting Rhizobacteria: Application in Biofertilizers and Biocontrol of Phytopathogens. In PGPR Amelioration in Sustainable Agriculture; Elsevier: Amsterdam, The Netherlands, 2019; pp. 41–66. [Google Scholar]
- Mukhtar, T.; Afridi, M.S.; McArthur, R.; Van Hamme, J.D.; Rineau, F.; Mahmood, T.; Amna; Sumaira; Zahid, M.; Salam, A. Draft Genome Sequence of Bacillus Pumilus SCAL1, an Endophytic Heat-Tolerant Plant Growth-Promoting Bacterium. Genome Announc. 2018, 6, e00306–e00318. [Google Scholar] [CrossRef] [PubMed]
- Sanders, C.L.; Eginger, K.H. Optimizing Care of the Mechanically Ventilated Patient in the Emergency Department through the Utilization of Validated Sedation Scoring Scales. J. Emerg. Nurs. 2017, 43, 84–86. [Google Scholar] [CrossRef] [PubMed]
- Afridi, M.S.; Mahmood, T.; Salam, A.; Mukhtar, T.; Mehmood, S.; Ali, J.; Khatoon, Z.; Bibi, M.; Javed, M.T.; Sultan, T. Induction of Tolerance to Salinity in Wheat Genotypes by Plant Growth Promoting Endophytes: Involvement of ACC Deaminase and Antioxidant Enzymes. Plant Physiol. Biochem. 2019, 139, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Afridi, M.S.; Van Hamme, J.D.; Bundschuh, J.; Khan, M.N.; Salam, A.; Waqar, M.; Munis, M.F.H.; Chaudhary, H.J. Biotechnological Approaches in Agriculture and Environmental Management-Bacterium Kocuria Rhizophila 14ASP as Heavy Metal and Salt-Tolerant Plant Growth-Promoting Strain. Biologia 2021, 76, 3091–3105. [Google Scholar] [CrossRef]
- Raj, M.; Kumar, R.; Lal, K.; Sirisha, L.; Chaudhary, R.; Patel, S.K. Dynamic Role of Plant Growth Promoting Rhizobacteria (PGPR) in Agriculture. Int. J. Chem. Stud. 2020, 8, 105–110. [Google Scholar] [CrossRef]
- Bennett, N.C.; Gardiner, R.A.; Hooper, J.D.; Johnson, D.W.; Gobe, G.C. Molecular Cell Biology of Androgen Receptor Signalling. Int. J. Biochem. Cell Biol. 2010, 42, 813–827. [Google Scholar] [CrossRef] [PubMed]
- Ravanbakhsh, M.; Sasidharan, R.; Voesenek, L.A.C.J.; Kowalchuk, G.A.; Jousset, A. Microbial Modulation of Plant Ethylene Signaling: Ecological and Evolutionary Consequences. Microbiome 2018, 6, 52. [Google Scholar] [CrossRef]
- Mhlongo, M.I.; Piater, L.A.; Madala, N.E.; Labuschagne, N.; Dubery, I.A. The Chemistry of Plant-Microbe Interactions in the Rhizosphere and the Potential for Metabolomics to Reveal Signaling Related to Defense Priming and Induced Systemic Resistance. Front. Plant Sci. 2018, 9, 112. [Google Scholar] [CrossRef]
- Remy, W.; Taylor, T.N.; Hass, H.; Kerp, H. Four Hundred-Million-Year-Old Vesicular Arbuscular Mycorrhizae. Proc. Natl. Acad. Sci. USA 1994, 91, 11841–11843. [Google Scholar] [CrossRef]
- Koziol, L.; Rieseberg, L.H.; Kane, N.; Bever, J.D. Reduced Drought Tolerance during Domestication and the Evolution of Weediness Results from Tolerance–Growth Trade-Offs. Evol. Int. J. Org. Evol. 2012, 66, 3803–3814. [Google Scholar] [CrossRef]
- Ahmad, M.; Ishaq, M.; Shah, W.A.; Adnan, M.; Fahad, S.; Saleem, M.H.; Khan, F.U.; Mussarat, M.; Khan, S.; Ali, B. Managing Phosphorus Availability from Organic and Inorganic Sources for Optimum Wheat Production in Calcareous Soils. Sustainability 2022, 14, 7669. [Google Scholar] [CrossRef]
- Roberts, D.P.; Maul, J.E.; McKenna, L.F.; Emche, S.E.; Meyer, S.L.F.; Collins, R.T.; Bowers, J.H. Selection of Genetically Diverse Trichoderma spp. Isolates for Suppression of Phytophthora capsici on Bell Pepper. Can. J. Microbiol. 2010, 56, 864–873. [Google Scholar] [CrossRef]
- Contreras-Cornejo, H.A.; Macías-Rodríguez, L.; del-Val, E.; Larsen, J. Ecological Functions of Trichoderma spp. and Their Secondary Metabolites in the Rhizosphere: Interactions with Plants. FEMS Microbiol. Ecol. 2016, 92, fiw036. [Google Scholar] [CrossRef]
- Haraguchi, S.; Yoshiga, T. Potential of the Fungal Feeding Nematode Aphelenchus Avenae to Control Fungi and the Plant Parasitic Nematode Ditylenchus Destructor Associated with Garlic. Biol. Control. 2020, 143, 104203. [Google Scholar] [CrossRef]
- Lagerlöf, J.; Insunza, V.; Lundegårdh, B.; Rämert, B. Interaction between a Fungal Plant Disease, Fungivorous Nematodes and Compost Suppressiveness. Acta Agric. Scand. Sect. B-Soil Plant Sci. 2011, 61, 372–377. [Google Scholar] [CrossRef]
- Martinuz, A.; Zewdu, G.; Ludwig, N.; Grundler, F.; Sikora, R.A.; Schouten, A. The Application of Arabidopsis Thaliana in Studying Tripartite Interactions among Plants, Beneficial Fungal Endophytes and Biotrophic Plant-Parasitic Nematodes. Planta 2015, 241, 1015–1025. [Google Scholar] [CrossRef]
- Burki, F. The Eukaryotic Tree of Life from a Global Phylogenomic Perspective. Cold Spring Harb. Perspect. Biol. 2014, 6, a016147. [Google Scholar] [CrossRef]
- Mahmud, K.; Missaoui, A.; Lee, K.; Ghimire, B.; Presley, H.W.; Makaju, S. Rhizosphere Microbiome Manipulation for Sustainable Crop Production. Curr. Plant Biol. 2021, 27, 100210. [Google Scholar] [CrossRef]
- SOLANKI, M.K.; Solanki, A.C.; Rai, S.; Srivastava, S.; Kashyap, B.K.; Divvela, P.K.; Kumar, S.; Yandigeri, M.; Kashyap, P.L.; Srivastava, A.K. Functional Interplay between Antagonistic Bacteria and Rhizoctonia solani in the Tomato Plant Rhizosphere. Front. Microbiol. 2022, 13, 990850. [Google Scholar] [CrossRef] [PubMed]
- Krome, K.; Rosenberg, K.; Dickler, C.; Kreuzer, K.; Ludwig-Müller, J.; Ullrich-Eberius, C.; Scheu, S.; Bonkowski, M. Soil Bacteria and Protozoa Affect Root Branching via Effects on the Auxin and Cytokinin Balance in Plants. Plant Soil 2010, 328, 191–201. [Google Scholar] [CrossRef]
- Hünninghaus, M.; Koller, R.; Kramer, S.; Marhan, S.; Kandeler, E.; Bonkowski, M. Changes in Bacterial Community Composition and Soil Respiration Indicate Rapid Successions of Protist Grazers during Mineralization of Maize Crop Residues. Pedobiologia 2017, 62, 1–8. [Google Scholar] [CrossRef]
- Jousset, A.; Lara, E.; Wall, L.G.; Valverde, C. Secondary Metabolites Help Biocontrol Strain Pseudomonas fluorescens CHA0 To Escape Protozoan Grazing. Appl. Environ. Microbiol. 2006, 72, 7083–7090. [Google Scholar] [CrossRef]
- Jousset, A.; Bonkowski, M. The Model Predator Acanthamoeba Castellanii Induces the Production of 2,4, DAPG by the Biocontrol Strain Pseudomonas fluorescens Q2-87. Soil Biol. Biochem. 2010, 42, 1647–1649. [Google Scholar] [CrossRef]
- Bonkowski, M.; Brandt, F. Do Soil Protozoa Enhance Plant Growth by Hormonal Effects? Soil Biol. Biochem. 2002, 34, 1709–1715. [Google Scholar] [CrossRef]
- Brazelton, J.N.; Pfeufer, E.E.; Sweat, T.A.; Gardener, B.B.M.; Coenen, C. 2,4-Diacetylphloroglucinol Alters Plant Root Development. Mol. Plant-Microbe Interact. 2008, 21, 1349–1358. [Google Scholar] [CrossRef]
- Kuppardt, A.; Fester, T.; Härtig, C.; Chatzinotas, A. Rhizosphere Protists Change Metabolite Profiles in Zea Mays. Front. Microbiol. 2018, 9, 857. [Google Scholar] [CrossRef] [PubMed]
- Springett, J.; Gray, R. The Interaction between Plant Roots and Earthworm Burrows in Pasture. Soil Biol. Biochem. 1997, 29, 621–625. [Google Scholar] [CrossRef]
- Yasmin, S.; D’Souza, D. Effects of Pesticides on the Growth and Reproduction of Earthworm: A Review. Appl. Environ. Soil Sci. 2010, 2010, e678360. [Google Scholar] [CrossRef]
- Brown, G.G.; Barois, I.; Lavelle, P. Regulation of Soil Organic Matter Dynamics and Microbial Activityin the Drilosphere and the Role of Interactionswith Other Edaphic Functional Domains§§Paper Presented at the 16th World Congress of Soil Science, 20–26 August 1998, Montpellier, France. Eur. J. Soil Biol. 2000, 36, 177–198. [Google Scholar] [CrossRef]
- Caravaca, F.; Roldán, A. Effect of Eisenia Foetida Earthworms on Mineralization Kinetics, Microbial Biomass, Enzyme Activities, Respiration and Labile C Fractions of Three Soils Treated with a Composted Organic Residue. Biol. Fertil. Soils 2003, 38, 45–51. [Google Scholar] [CrossRef]
- Gong, X.; Jiang, Y.; Zheng, Y.; Chen, X.; Li, H.; Hu, F.; Liu, M.; Scheu, S. Earthworms Differentially Modify the Microbiome of Arable Soils Varying in Residue Management. Soil Biol. Biochem. 2018, 121, 120–129. [Google Scholar] [CrossRef]
- Bedano, J.C.; Vaquero, F.; Domínguez, A.; Rodríguez, M.P.; Wall, L.; Lavelle, P. Earthworms Contribute to Ecosystem Process in No-till Systems with High Crop Rotation Intensity in Argentina. Acta Oecologica 2019, 98, 14–24. [Google Scholar] [CrossRef]
- Guenet, B.; Neill, C.; Bardoux, G.; Abbadie, L. Is There a Linear Relationship between Priming Effect Intensity and the Amount of Organic Matter Input? Appl. Soil Ecol. 2010, 46, 436–442. [Google Scholar] [CrossRef]
- Wei, S.; Jacquiod, S.; Philippot, L.; Blouin, M.; Sørensen, S.J. Spatial Analysis of the Root System Coupled to Microbial Community Inoculation Shed Light on Rhizosphere Bacterial Community Assembly. Biol. Fertil. Soils 2021, 57, 973–989. [Google Scholar] [CrossRef]
- Martínez-Medina, A.; Fernandez, I.; Lok, G.B.; Pozo, M.J.; Pieterse, C.M.; Van Wees, S.C. Shifting from Priming of Salicylic Acid-to Jasmonic Acid-Regulated Defences by Trichoderma Protects Tomato against the Root Knot Nematode Meloidogyne Incognita. New Phytol. 2017, 213, 1363–1377. [Google Scholar] [CrossRef] [PubMed]
- Elmer, W.H. Influence of Earthworm Activity on Soil Microbes and Soilborne Diseases of Vegetables. Plant Dis. 2009, 93, 175–179. [Google Scholar] [CrossRef]
- Stephens, P.M.; Davoren, C.W. Influence of the Earthworms Aporrectodea Trapezoides and A. Rosea on the Disease Severity of Rhizoctonia Solani on Subterranean Clover and Ryegrass. Soil Biol. Biochem. 1997, 29, 511–516. [Google Scholar] [CrossRef]
- Bernard, L.; Chapuis-Lardy, L.; Razafimbelo, T.; Razafindrakoto, M.; Pablo, A.-L.; Legname, E.; Poulain, J.; Brüls, T.; O’Donohue, M.; Brauman, A.; et al. Endogeic Earthworms Shape Bacterial Functional Communities and Affect Organic Matter Mineralization in a Tropical Soil. ISME J. 2012, 6, 213–222. [Google Scholar] [CrossRef]
- Shan, J.; Liu, J.; Wang, Y.; Yan, X.; Guo, H.; Li, X.; Ji, R. Digestion and Residue Stabilization of Bacterial and Fungal Cells, Protein, Peptidoglycan, and Chitin by the Geophagous Earthworm Metaphire Guillelmi. Soil Biol. Biochem. 2013, 64, 9–17. [Google Scholar] [CrossRef]
- Khomyakov, N.V.; Kharin, S.A.; Nechitailo, T.Y.; Golyshin, P.N.; Kurakov, A.V.; Byzov, B.A.; Zvyagintsev, D.G. Reaction of Microorganisms to the Digestive Fluid of Earthworms. Microbiology 2007, 76, 45–54. [Google Scholar] [CrossRef]
- Fujii, K.; Ikeda, K.; Yoshida, S. Isolation and Characterization of Aerobic Microorganisms with Cellulolytic Activity in the Gut of Endogeic Earthworms. Int. Microbiol. 2012, 15, 121–130. [Google Scholar] [CrossRef]
- Hong, S.W.; Lee, J.S.; Chung, K.S. Effect of Enzyme Producing Microorganisms on the Biomass of Epigeic Earthworms (Eisenia fetida) in Vermicompost. Bioresour. Technol. 2011, 102, 6344–6347. [Google Scholar] [CrossRef]
- McLean, M.A.; Migge-Kleian, S.; Parkinson, D. Earthworm Invasions of Ecosystems Devoid of Earthworms: Effects on Soil Microbes. Biol. Invasions 2006, 8, 1257–1273. [Google Scholar] [CrossRef]
- Roman-Reyna, V.; Pinili, D.; Borja, F.N.; Quibod, I.L.; Groen, S.C.; Alexandrov, N.; Mauleon, R.; Oliva, R. Characterization of the Leaf Microbiome from Whole-Genome Sequencing Data of the 3000 Rice Genomes Project. Rice 2020, 13, 72. [Google Scholar] [CrossRef] [PubMed]
- Nunan, N.; Wu, K.; Young, I.M.; Crawford, J.W.; Ritz, K. Spatial Distribution of Bacterial Communities and Their Relationships with the Micro-Architecture of Soil. FEMS Microbiol. Ecol. 2003, 44, 203–215. [Google Scholar] [CrossRef] [PubMed]
- Villegas, L.M.; Pimenta, P.F.P. Metagenomics, Paratransgenesis and the Anopheles Microbiome: A Portrait of the Geographical Distribution of the Anopheline Microbiota Based on a Meta-Analysis of Reported Taxa. Mem. Inst. Oswaldo Cruz 2014, 109, 672–684. [Google Scholar] [CrossRef] [PubMed]
- Anwar, T.; Qureshi, H.; Parveen, N.; Mahmood, S.; Haider, M.Z.; Mumtaz, S.; Nawaz, H.; Khan, S.A.; Hafeez, A.; Tipu, M.I.; et al. Herbicidal effectiveness of wild poisonous plant Rhazya stricta using different media by the sandwich method. Pak. J. Bot. 2023, 55, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Dollive, S.; Peterfreund, G.L.; Sherrill-Mix, S.; Bittinger, K.; Sinha, R.; Hoffmann, C.; Nabel, C.S.; Hill, D.A.; Artis, D.; Bachman, M.A.; et al. A Tool Kit for Quantifying Eukaryotic RRNA Gene Sequences from Human Microbiome Samples. Genome Biol. 2012, 13, R60. [Google Scholar] [CrossRef] [PubMed]
- Nicora, G.; Zucca, S.; Limongelli, I.; Bellazzi, R.; Magni, P. A Machine Learning Approach Based on ACMG/AMP Guidelines for Genomic Variant Classification and Prioritization. Sci. Rep. 2022, 12, 2517. [Google Scholar] [CrossRef]
- Ali, B.; Wang, X.; Saleem, M.H.; Hafeez, A.; Afridi, M.S.; Khan, S.; Ullah, I.; do Amaral Júnior, A.T.; Alatawi, A.; Ali, S. PGPR-Mediated Salt Tolerance in Maize by Modulating Plant Physiology, Antioxidant Defense, Compatible Solutes Accumulation and Bio-Surfactant Producing Genes. Plants 2022, 11, 345. [Google Scholar] [CrossRef]
- Rafique, M.; Hayat, K.; Mukhtar, T.; Khan, A.A.; Afridi, M.S.; Hussain, T.; Sultan, T.; Munis, M.F.H.; Imran, M.; Chaudhary, H.J. Bacterial Biofilm Formation and Its Role against Agricultural Pathogens. In The Battle against Microbial Pathogens: Basic Science, Technological Advances and Educational Programs; Formatex Research Center: Badajoz, Spain, 2015; pp. 373–382. [Google Scholar]
- Saikkonen, K.; Nissinen, R.; Helander, M. Toward Comprehensive Plant Microbiome Research. Front. Ecol. Evol. 2020, 8, 61. [Google Scholar] [CrossRef]
- Cheng, Z.; Duan, J.; Hao, Y.; McConkey, B.J.; Glick, B.R. Identification of Bacterial Proteins Mediating the Interactions between Pseudomonas putida UW4 and Brassica napus (Canola). Mol. Plant Microbe Interact. 2009, 22, 686–694. [Google Scholar] [CrossRef]
- Novello, G.; Gamalero, E.; Bona, E.; Boatti, L.; Mignone, F.; Massa, N.; Cesaro, P.; Lingua, G.; Berta, G. The Rhizosphere Bacterial Microbiota of Vitis vinifera cv. Pinot Noir in an Integrated Pest Management Vineyard. Front. Microbiol. 2017, 8, 1528. [Google Scholar] [CrossRef]
- Ali, S.; Badshah, G.; Ali, U.; Siddique Afridi, M.; Shamim, A.; Khan, A.; Luiz Felipe Soares, F.; Rocha Alencar Menezes, L.; Theodoro Rezende, V.; Barison, A.; et al. Leaf Tissue Metabolomics Fingerprinting of Citronella gongonha Mart. by 1H HR-MAS NMR. Sci. Rep. 2022, 12, 17624. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Rech, K.S.; Badshah, G.; Soares, F.L.; Barison, A. 1H HR-MAS NMR-Based Metabolomic Fingerprinting to Distinguish Morphological Similarities and Metabolic Profiles of Maytenus ilicifolia, a Brazilian Medicinal Plant. J. Nat. Prod. 2021, 84, 1707–1714. [Google Scholar] [CrossRef]
- Ali, S.; Badshah, G.; D’Oca, C.D.R.M.; Ramos Campos, F.; Nagata, N.; Khan, A.; de Fátima Costa Santos, M.; Barison, A. High-Resolution Magic Angle Spinning (HR-MAS) NMR-Based Fingerprints Determination in the Medicinal Plant Berberis laurina. Molecules 2020, 25, 3647. [Google Scholar] [CrossRef]
- Ocampos, F.M.; Menezes, L.R.; Dutra, L.M.; Santos, M.F.; Ali, S.; Barison, A. NMR in Chemical Ecology: An Overview Highlighting the Main NMR Approaches. eMagRes 2007, 6, 325–342. [Google Scholar]
- Levy, A.; Salas Gonzalez, I.; Mittelviefhaus, M.; Clingenpeel, S.; Herrera Paredes, S.; Miao, J.; Wang, K.; Devescovi, G.; Stillman, K.; Monteiro, F.; et al. Genomic Features of Bacterial Adaptation to Plants. Nat. Genet. 2017, 50, 138–150. [Google Scholar] [CrossRef] [PubMed]
- Wahab, A.; Abdi, G.; Saleem, M.H.; Ali, B.; Ullah, S.; Shah, W.; Mumtaz, S.; Yasin, G.; Muresan, C.C.; Marc, R.A. Plants’ Physio-Biochemical and Phyto-Hormonal Responses to Alleviate the Adverse Effects of Drought Stress: A Comprehensive Review. Plants 2022, 11, 1620. [Google Scholar] [CrossRef] [PubMed]
- Javed, S.; Azeem, M.; Mahmood, S.; Al-Anazi, K.M.; Farah, M.A.; Ali, S.; Ali, B. Biotransformation of Agricultural Wastes into Lovastatin and Optimization of a Fermentation Process Using Response Surface Methodology (RSM). Agronomy 2022, 12, 2848. [Google Scholar] [CrossRef]
- Afridi, M.S.; Javed, M.A.; Ali, S.; De Medeiros, F.H.V.; Ali, B.; Salam, A.; Marc, R.A.; Alkhalifah, D.H.M.; Selim, S.; Santoyo, G. New Opportunities in Plant Microbiome Engineering for Increasing Agricultural Sustainability under Stressful Conditions. Front. Plant Sci. 2022, 13, 899464. [Google Scholar] [CrossRef] [PubMed]
- Guerra, C.A.; Heintz-Buschart, A.; Sikorski, J.; Chatzinotas, A.; Guerrero-Ramírez, N.; Cesarz, S.; Beaumelle, L.; Rillig, M.C.; Maestre, F.T.; Delgado-Baquerizo, M. Blind Spots in Global Soil Biodiversity and Ecosystem Function Research. Nat. Commun. 2020, 11, 3870. [Google Scholar] [CrossRef]
- Aryantha, I.P.; Cross, R.; Guest, D.I. Suppression of Phytophthora cinnamomi in Potting Mixes Amended with Uncomposted and Composted Animal Manures. Phytopathology 2000, 90, 775–782. [Google Scholar] [CrossRef]
- Shi, W.; Li, M.; Wei, G.; Tian, R.; Li, C.; Wang, B.; Lin, R.; Shi, C.; Chi, X.; Zhou, B.; et al. The Occurrence of Potato Common Scab Correlates with the Community Composition and Function of the Geocaulosphere Soil Microbiome. Microbiome 2019, 7, 14. [Google Scholar] [CrossRef] [PubMed]
- Uroz, S.; Courty, P.E.; Oger, P. Plant Symbionts Are Engineers of the Plant-Associated Microbiome. Trends Plant Sci. 2019, 24, 905–916. [Google Scholar] [CrossRef]
- Durán, P.; Tortella, G.; Viscardi, S.; Barra, P.J.; Carrión, V.J.; Mora, M.d.l.L.; Pozo, M.J. Microbial Community Composition in Take-All Suppressive Soils. Front. Microbiol. 2018, 9, 2198. [Google Scholar] [CrossRef]
- Penton, C.R.; Gupta, V.V.S.R.; Tiedje, J.M.; Neate, S.M.; Ophel-Keller, K.; Gillings, M.; Harvey, P.; Pham, A.; Roget, D.K. Fungal Community Structure in Disease Suppressive Soils Assessed by 28S LSU Gene Sequencing. PLoS ONE 2014, 9, e93893. [Google Scholar] [CrossRef]
- Szabó, M.; Csepregi, K.; Gálber, M.; Virányi, F.; Fekete, C. Control Plant-Parasitic Nematodes with Trichoderma Species and Nematode-Trapping Fungi: The Role of Chi18-5 and Chi18-12 Genes in Nematode Egg-Parasitism. Biol. Control 2012, 63, 121–128. [Google Scholar] [CrossRef]
- Askary, T.H. Nematophagous Fungi as Biocontrol Agents of Phytonematodes. In Biocontrol Agents Phytonematodes; CAB International: Wallingford, UK, 2015; pp. 81–125. [Google Scholar] [CrossRef]
- Jayaraman, S.; Naorem, A.K.; Lal, R.; Dalal, R.C.; Sinha, N.K.; Patra, A.K.; Chaudhari, S.K. Disease-Suppressive Soils—Beyond Food Production: A Critical Review. J. Soil Sci. Plant Nutr. 2021, 21, 1437–1465. [Google Scholar] [CrossRef]
- Abiven, S.; Menasseri, S.; Chenu, C. The Effects of Organic Inputs over Time on Soil Aggregate Stability–A Literature Analysis. Soil Biol. Biochem. 2009, 41, 1–12. [Google Scholar] [CrossRef]
- Bailey, K.L.; Lazarovits, G. Suppressing Soil-Borne Diseases with Residue Management and Organic Amendments. Soil Tillage Res. 2003, 72, 169–180. [Google Scholar] [CrossRef]
- Yang, Y.; Awasthi, M.K.; Bao, H.; Bie, J.; Lei, S.; Lv, J. Exploring the Microbial Mechanisms of Organic Matter Transformation during Pig Manure Composting Amended with Bean Dregs and Biochar. Bioresour. Technol. 2020, 313, 123647. [Google Scholar] [CrossRef] [PubMed]
- Ali, B.; Hafeez, A.; Ahmad, S.; Javed, M.A.; Afridi, M.S.; Dawoud, T.M.; Almaary, K.S.; Muresan, C.C.; Marc, R.A.; Alkhalifah, D.H.M. Bacillus Thuringiensis PM25 Ameliorates Oxidative Damage of Salinity Stress in Maize via Regulating Growth, Leaf Pigments, Antioxidant Defense System, and Stress Responsive Gene Expression. Front. Plant Sci. 2022, 13, 921668. [Google Scholar] [CrossRef] [PubMed]
- Spokas, K.A.; Novak, J.M.; Venterea, R.T. Biochar’s Role as an Alternative N-Fertilizer: Ammonia Capture. Plant Soil 2012, 350, 35–42. [Google Scholar] [CrossRef]
- Eyles, A.; Bound, S.A.; Oliver, G.; Corkrey, R.; Hardie, M.; Green, S.; Close, D.C. Impact of Biochar Amendment on the Growth, Physiology and Fruit of a Young Commercial Apple Orchard. Trees 2015, 29, 1817–1826. [Google Scholar] [CrossRef]
- Rafique, M.; Ortas, I.; Ahmed, I.A.; Rizwan, M.; Afridi, M.S.; Sultan, T.; Chaudhary, H.J. Potential Impact of Biochar Types and Microbial Inoculants on Growth of Onion Plant in Differently Textured and Phosphorus Limited Soils. J. Environ. Manag. 2019, 247, 672–680. [Google Scholar] [CrossRef] [PubMed]
- Mehta, C.M.; Palni, U.; Franke-Whittle, I.H.; Sharma, A.K. Compost: Its Role, Mechanism and Impact on Reducing Soil-Borne Plant Diseases. Waste Manag. 2014, 34, 607–622. [Google Scholar] [CrossRef] [PubMed]
- Baltar, F.; Palovaara, J.; Unrein, F.; Catala, P.; Horňák, K.; Šimek, K.; Vaqué, D.; Massana, R.; Gasol, J.M.; Pinhassi, J. Marine Bacterial Community Structure Resilience to Changes in Protist Predation under Phytoplankton Bloom Conditions. ISME J. 2016, 10, 568–581. [Google Scholar] [CrossRef] [PubMed]
- Savary, S.; Ficke, A.; Aubertot, J.-N.; Hollier, C. Crop Losses Due to Diseases and Their Implications for Global Food Production Losses and Food Security. Food Secur. 2012, 4, 519–537. [Google Scholar] [CrossRef]
- Rich, J.R.; Rahi, G.S. Suppression of Meloidogyne Javanica and M. Incognita on Tomato with Ground Seed of Castor, Crotalaria, Hairy Indigo, and Wheat. Nematropica 1995, 25, 159–164. [Google Scholar]
- Kokalis-Burelle, N.; Rodríguez-Kábana, R. Changes in Populations of Soil Microorganisms, Nematodes, and Enzyme Activity Associated with Application of Powdered Pine Bark. Plant Soil 1994, 162, 169–175. [Google Scholar] [CrossRef]
- Culbreath, A.K.; Rodriguez-Kabana, R.; Morgan-Jones, G. The Use of Hemicellulosic Waste Matter for Reduction of the Phytotoxic Effects of Chitin and Control of Root-Knot Nematodes. Nematropica 1985, 15, 49–75. [Google Scholar]
- Mian, I.H.; Godoy, G.; Shelby, R.A.; Rodriguez-Kabana, R.; Morgan-Jones, G. Chitin Amendments for Control of Meloidogyne Arenaria in Infested Soil. Nematropica 1982, 13, 71–84. [Google Scholar]
- Kloepper, J.W. Plant Growth-Promoting Rhizobacteria (Other Systems). Azospirillum/Plant Assoc. 1994, 187, 137–166. [Google Scholar]
- Berne, C.; Kysela, D.T.; Brun, Y.V. A Bacterial Extracellular DNA Inhibits Settling of Motile Progeny Cells within a Biofilm. Mol. Microbiol. 2010, 77, 815–829. [Google Scholar] [CrossRef] [PubMed]
- Qtu, J.; Hallmann, J.; Kokalis-Burelle, N.; Weaver, D.B.; Rodríguez-Kábana, R.; Tuzun, S. Activity and Differential Induction of Chitinase Isozymes in Soybean Cultivars Resistant or Susceptible to Root-Knot Nematodes. J. Nematol. 1997, 29, 523. [Google Scholar] [PubMed]
- Coventry, E.; Noble, R.; Mead, A.; Whipps, J.M. Suppression of Allium White Rot (Sclerotium cepivorum) in Different Soils Using Vegetable Wastes. Eur. J. Plant Pathol. 2005, 111, 101–112. [Google Scholar] [CrossRef]
- Negm, N.A.; Aiad, I.A.; Tawfik, S.M. Screening for Potential Antimicrobial Activities of Some Cationic Uracil Biocides Against Wide-Spreading Bacterial Strains. J. Surfactants Deterg. 2010, 13, 503–511. [Google Scholar] [CrossRef]
- Castaño, R.; Borrero, C.; Avilés, M. Organic Matter Fractions by SP-MAS 13C NMR and Microbial Communities Involved in the Suppression of Fusarium Wilt in Organic Growth Media. Biol. Control 2011, 58, 286–293. [Google Scholar] [CrossRef]
- Cuiyue, D. Effects of Biological Metabolism Of Metasequoia Glyptostroboides On Nutrient Element Content and Enzyme Activity in Seedling Soil. Turk. J. Agric. For. 2021, 45, 642–650. [Google Scholar]
- Akram, N.A.; Saleem, M.H.; Shafiq, S.; Naz, H.; Farid-ul-Haq, M.; Ali, B.; Shafiq, F.; Iqbal, M.; Jaremko, M.; Qureshi, K.A. Phytoextracts as Crop Biostimulants and Natural Protective Agents—A Critical Review. Sustainability 2022, 14, 14498. [Google Scholar] [CrossRef]
- Gul, A.; Salam, A.; Afridi, M.S.; Bangash, N.K.; Ali, F.; Ali, M.Y.; Khan, S.; Mubeeen, R. Effect of Urea, Bio-Fertilizers and Their Interaction on the Growth, Yield and Yield Attributes of Cyamopsis tetragonoloba. Indian J. Agric. Res. 2019, 53, 423–428. [Google Scholar] [CrossRef]
- Saini, A.; Manuja, S.; Kumar, S.; Hafeez, A.; Ali, B.; Poczai, P. Impact of Cultivation Practices and Varieties on Productivity, Profitability, and Nutrient Uptake of Rice (Oryza sativa L.) and Wheat (Triticum aestivum L.) Cropping System in India. Agriculture 2022, 12, 1678. [Google Scholar] [CrossRef]
- Farooq, T.H.; Rafay, M.; Basit, H.; Shakoor, A.; Shabbir, R.; Riaz, M.U.; Ali, B.; Kumar, U.; Qureshi, K.A.; Jaremko, M. Morpho-physiological growth performance and phytoremediation capabilities of selected xerophyte grass species toward Cr and Pb stress. Front. Plant Sci. 2022, 13, 997120. [Google Scholar] [CrossRef] [PubMed]
- Adnan, M.; Fahad, S.; Saleem, M.H.; Ali, B.; Mussart, M.; Ullah, R.; Arif, M.; Ahmad, M.; Shah, W.A.; Romman, M. Comparative Efficacy of Phosphorous Supplements with Phosphate Solubilizing Bacteria for Optimizing Wheat Yield in Calcareous Soils. Sci. Rep. 2022, 12, 11997. [Google Scholar] [CrossRef] [PubMed]
- Hayat, R.; Ali, S.; Amara, U.; Khalid, R.; Ahmed, I. Soil Beneficial Bacteria and Their Role in Plant Growth Promotion: A Review. Ann. Microbiol. 2010, 60, 579–598. [Google Scholar] [CrossRef]
- Cojocaru, A.; Vlase, L.; Munteanu, N.; Stan, T.; Teliban, G.C.; Burducea, M.; Stoleru, V. Dynamic of Phenolic Compounds, Antioxidant Activity, and Yield of Rhubarb under Chemical, Organic and Biological Fertilization. Plants 2020, 9, 355. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Sai, G.; Jousset, A.; Qing Yun, Z.; Hua Song, W.; Rong, L.; Kowalchuk, G.A.; Qi Rong, S. Bio-Fertilizer Application Induces Soil Suppressiveness against Fusarium Wilt Disease by Reshaping the Soil Microbiome. Soil Biol. Biochem. 2017, 114, 238–247. [Google Scholar]
- Cai, F.; Pang, G.; Li, R.-X.; Li, R.; Gu, X.-L.; Shen, Q.-R.; Chen, W. Bioorganic Fertilizer Maintains a More Stable Soil Microbiome than Chemical Fertilizer for Monocropping. Biol. Fertil. Soils 2017, 53, 861–872. [Google Scholar] [CrossRef]
- Seo, J.M.; Dao, H.T.T.; Park, B.B. Growth and Nutrient Responses of Betula platyphylla, Larix kaempferi, and Chamaecyparis obtusa to Different Application Methods of Solid Compound Fertilizer. Tur. J. Agric. For. 2021, 45, 235–265. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Afridi, M.S.; Ali, S.; Salam, A.; César Terra, W.; Hafeez, A.; Sumaira; Ali, B.; S. AlTami, M.; Ameen, F.; Ercisli, S.; et al. Plant Microbiome Engineering: Hopes or Hypes. Biology 2022, 11, 1782. https://doi.org/10.3390/biology11121782
Afridi MS, Ali S, Salam A, César Terra W, Hafeez A, Sumaira, Ali B, S. AlTami M, Ameen F, Ercisli S, et al. Plant Microbiome Engineering: Hopes or Hypes. Biology. 2022; 11(12):1782. https://doi.org/10.3390/biology11121782
Chicago/Turabian StyleAfridi, Muhammad Siddique, Sher Ali, Abdul Salam, Willian César Terra, Aqsa Hafeez, Sumaira, Baber Ali, Mona S. AlTami, Fuad Ameen, Sezai Ercisli, and et al. 2022. "Plant Microbiome Engineering: Hopes or Hypes" Biology 11, no. 12: 1782. https://doi.org/10.3390/biology11121782
APA StyleAfridi, M. S., Ali, S., Salam, A., César Terra, W., Hafeez, A., Sumaira, Ali, B., S. AlTami, M., Ameen, F., Ercisli, S., Marc, R. A., Medeiros, F. H. V., & Karunakaran, R. (2022). Plant Microbiome Engineering: Hopes or Hypes. Biology, 11(12), 1782. https://doi.org/10.3390/biology11121782