
Dietary Habits and Tusk Usage of Shovel-Tusked Gomphotheres from Florida: Evidence from Stereoscopic Wear of Molars and Upper and Lower Tusks



Abstract
Simple Summary
Abstract
1. Introduction
1.1. Background
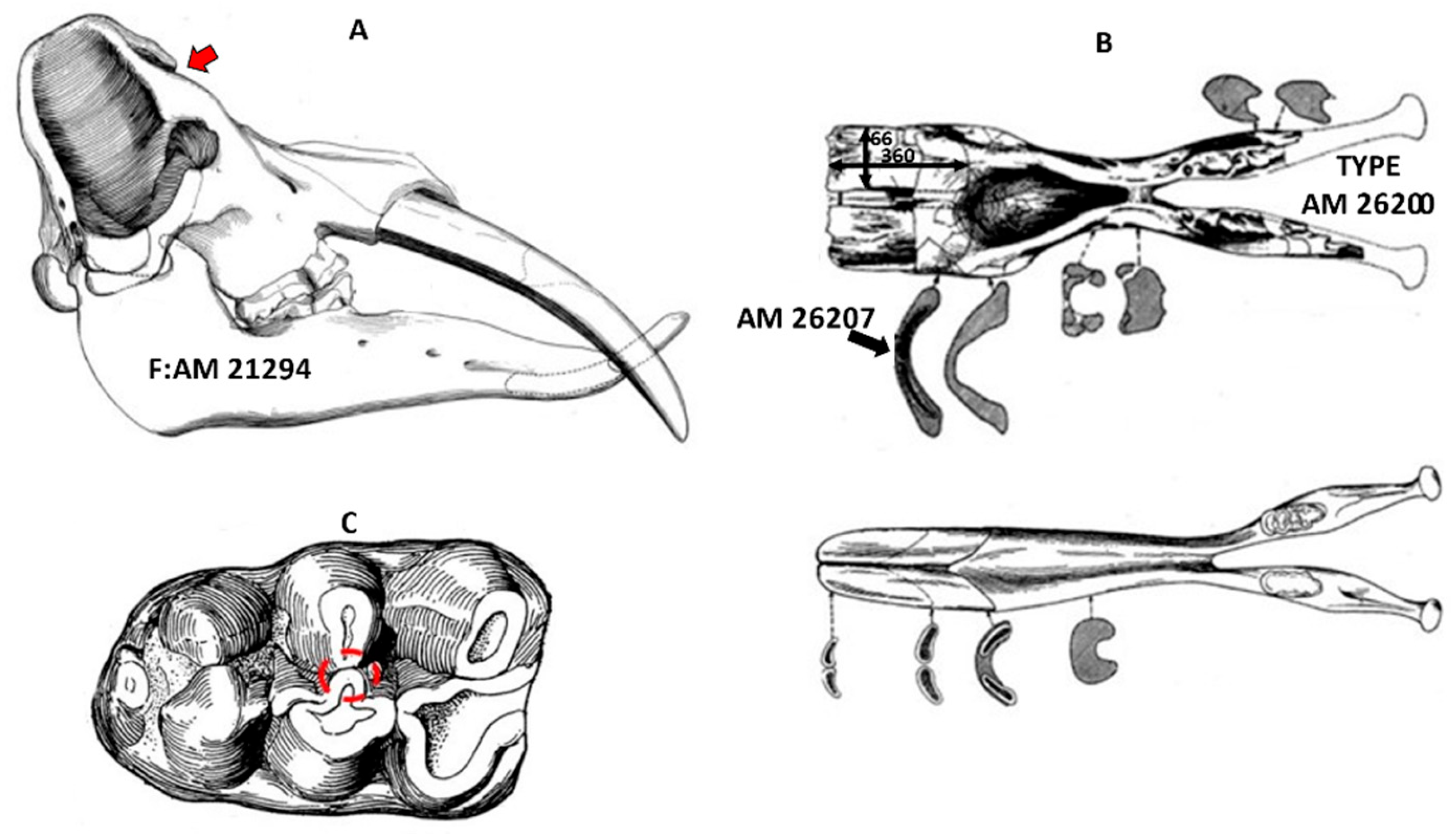
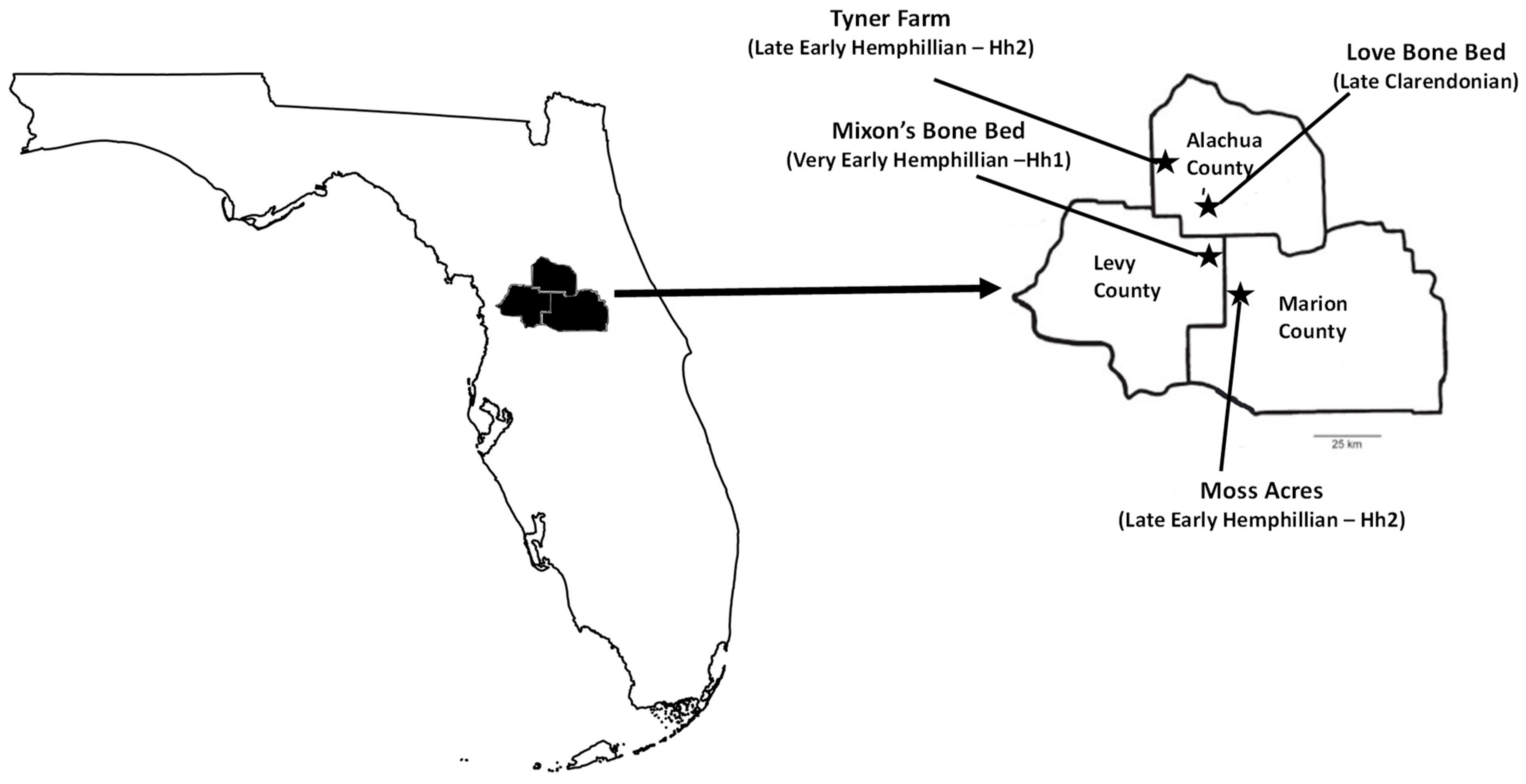
1.2. Fossil Localities
1.3. Aims of the Study
1.4. Microwear
2. Materials and Methods
2.1. Sample
2.2. Microwear Technique
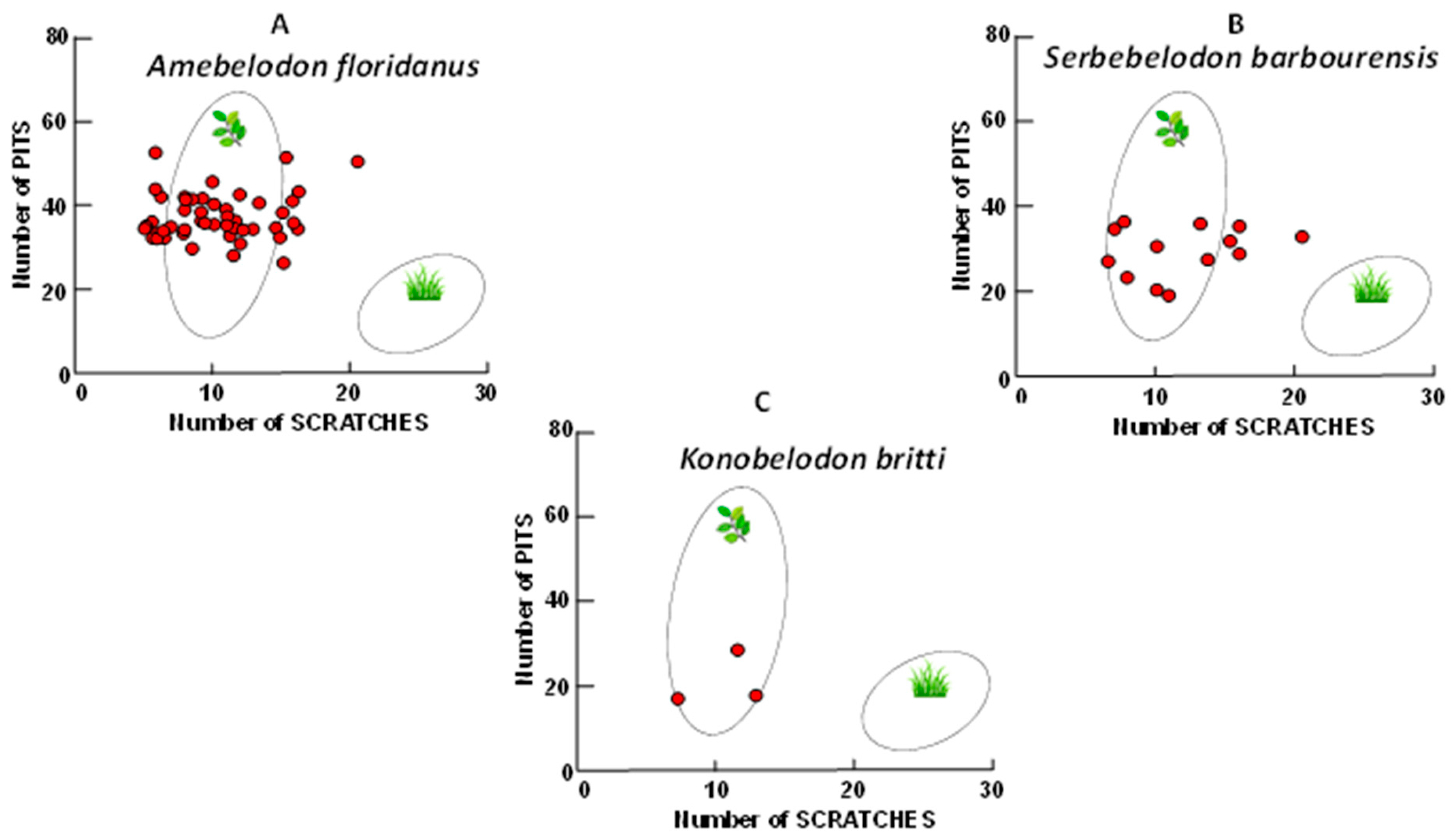
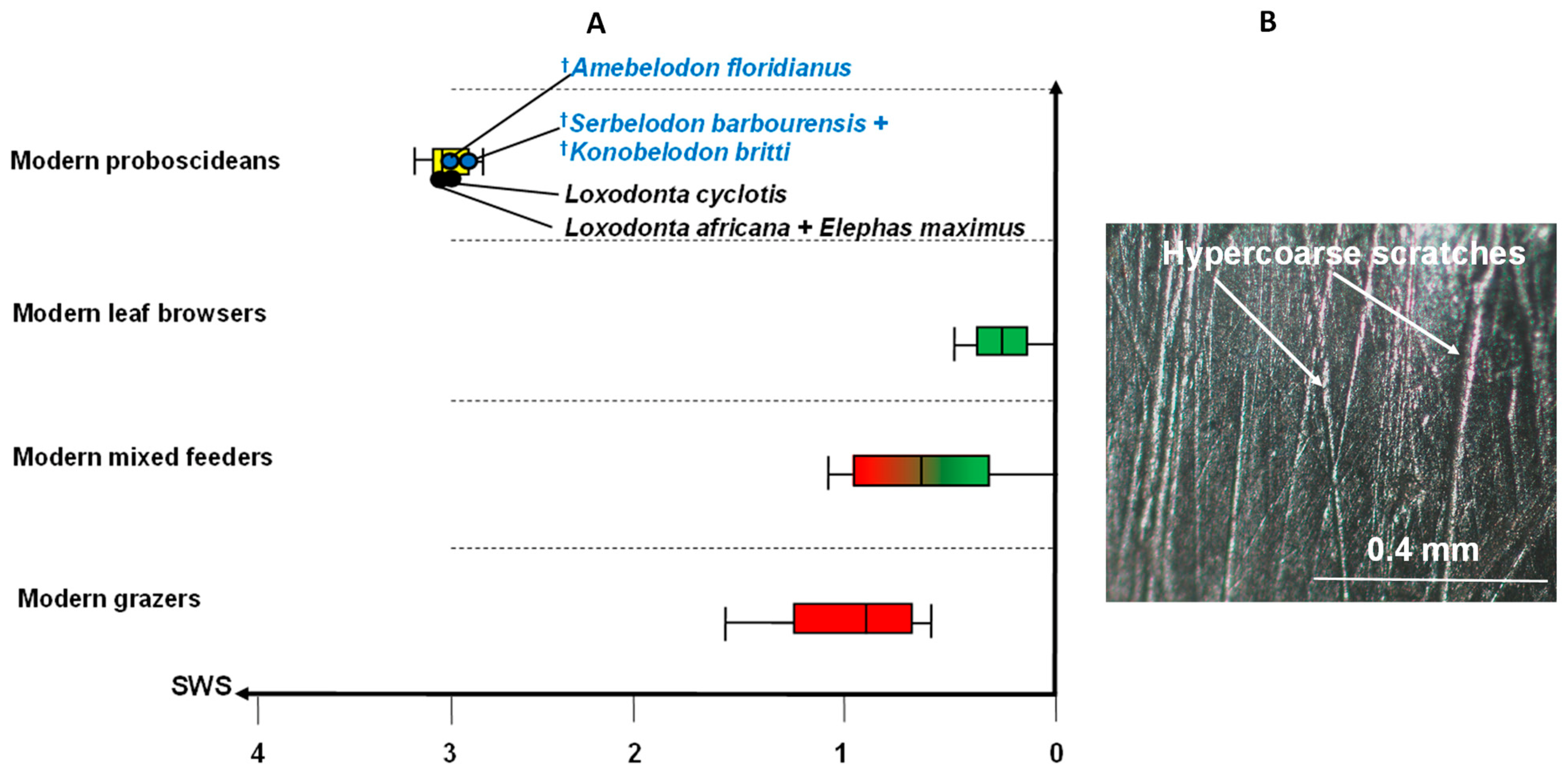
3. Results
4. Discussion
4.1. Value of Microwear
4.2. Fossil Site Paleoecology
4.3. Molar Microwear
4.4. Tusk Microwear
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tassy, P. The Earliest Gomphotheres. In The Proboscidea: Evolution and Palaeoecology of Elephants and Their Relatives; Shoshani, J., Tassy, P., Eds.; Oxford University Press: Oxford, UK, 1996. [Google Scholar]
- Lambert, W.D. The Biogeography of the Gomphotheriid Proboscideans of North America. In The Proboscidea: Evolution and Palaeoecology of Elephants and Their Relatives; Shoshani, J., Tassy, P., Eds.; Oxford University Press: Oxford, UK, 1996; pp. 143–148. [Google Scholar]
- Webb, S.D. Mammalian faunal dynamics of the Great American Interchange. Paleobiology 1976, 2, 220–232. [Google Scholar] [CrossRef]
- Fox, D.L.; Fisher, D.C. Dietary reconstruction of Miocene Gomphotherium (Mammalia, Proboscidea) from the Great Plains region, USA, based on the carbon isotope composition of tusk and molar enamel. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2004, 206, 311–335. [Google Scholar] [CrossRef]
- Asevedo, L.; Winck, G.R.; Mothe, D.; Avilla, L.S. Ancient diet of the Pleistocene gomphothere Notiomastodon platensis (Mammalia, Proboscidea, Gomphotheriidae) from lowland mid-latitudes of South America: Stereomicrowear and tooth calculus analyses combined. Quat. Int. 2012, 255, 42–52. [Google Scholar] [CrossRef]
- Domingo, L.; Koch, P.L.; Grimes, S.T.; Morales, J.; Lόpez-Martínez, N. Isotopic paleoecology of mammals and the middle Miocene cooling event in the Madrid Basin (Spain). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2012, 339–341, 98–113. [Google Scholar] [CrossRef]
- Shoshani, J.; Tassy, P. Advances in proboscidean taxonomy and classification, anatomy and physiology, and ecology and behavior. Quat. Intrl. 2005, 126–128, 5–20. [Google Scholar] [CrossRef]
- Frick, C. New remains of Trilophodont-tetrabelodont mastodons. Bul. AMNH 1933, 59, 1883–1965. [Google Scholar]
- Osborn, H.F.; Granger, W. Platybelodon grangeri, three growth stages, and a new serridentine from Mongolia. Am. Mus. Novit. 1932, 537, 1–13. [Google Scholar]
- Lucas, S.G.; Morgan, G.S. Taxonomy of Rhynchotherium (Mammalia, Proboscidea) from the Miocene-Pliocene of North America. Neogene Mamm. Bull. 2008, 44, 71. [Google Scholar]
- Osborn, H.F.; Granger, W. The shovel-tuskers, Amebelodontinae, of central Asia. Am. Mus. Novit. 1931, 470, 1–12. [Google Scholar]
- Osborn, H.F. Proboscidea; American Museum Press: New York, NY, USA, 1936; Volume 1. [Google Scholar]
- Gheerbrant, E.; Tassy, P. Origin and evolution of proboscideans (Translated from French). C R Palevol. 2009, 8, 281–294. [Google Scholar] [CrossRef]
- Markov, G.N.; Vergiev, S. First report of cf. Protanancus (Mammalia, Proboscidea, Amebelodontidae) from Europe. Geodiversitas 2010, 32, 493–500. [Google Scholar] [CrossRef]
- Shoshani, J. Gomphotheres and their position within Proboscidea. In The Proboscidea: Evolution and Palaeoecology of Elephants and Their Relatives; Shoshani, J., Tassy, P., Eds.; Oxford University Press: Oxford, UK, 1996; pp. 149–177. [Google Scholar]
- Sanders, W.J.; Miller, E.R. New proboscideans from the early Miocene of Wadi Moghara, Egypt. J. Vert. Paleontol. 2002, 22, 388–404. [Google Scholar] [CrossRef]
- Leidy, J. Vertebrate fossils from Florida. Proc. Acad. Nat. Sci. Phil. 1885, 36, 118–119. [Google Scholar]
- Leidy, J. Fossil bones from Florida. Proc. Acad. Nat. Sci. Phil. 1888, 1884, 309–310. [Google Scholar]
- Hulbert, R.C., Jr.; Poyer, A.R.; Webb, S.D. Tyner Farm, a new early Hemphillian local fauna from north-central Florida. J. Vert. Paleontol. 2002, 22, 68A. [Google Scholar]
- Webb, S.D.; MacFadden, B.J.; Baskin, J.A. Geology and paleontology of the Love Bone Bed from the late Miocene of Florida. Am. J. Sci. 1981, 281, 513–544. [Google Scholar] [CrossRef]
- Hulbert, R.C., Jr. Late Miocene Tapirus (Mammalia, Perissodactyla) from Florida, with description of a new species, Tapirus webbi. Bull. Fla. Mus. Nat. Hist. 2005, 45, 465–494. [Google Scholar]
- Rensberger, J.M. Scanning electron microscopy of wear and occlusal events in some small herbivores. In Development, Function and Evolution of Teeth; Butler, P.M., Joysey, K.A., Eds.; Academic Press: London, UK, 1978; pp. 415–438. [Google Scholar]
- Walker, A.; Hoeck, H.N.; Perez, L. Microwear of Mammalian Teeth as an Indicator of Diet. Science 1978, 201, 908–910. [Google Scholar] [CrossRef]
- Solounias, N.; Semprebon, G. Advances in the Reconstruction of Ungulate Ecomorphology with Application to Early Fossil Equids. Am. Mus. Novit. 2002, 3366, 1–49. [Google Scholar] [CrossRef]
- Semprebon, G.M.; Godfrey, L.R.; Solounias, N.; Sutherland, M.R.; Jungers, W.L. Can low-magnification stereomicroscopy reveal diet? J. Hum. Evol. 2004, 47, 115–144. [Google Scholar] [CrossRef]
- Merceron, G.; Blondel, C.; Brunet, M.; Sen, S.; Solounias, N.; Viriot, L.; Heintz, E. The Late Miocene paleoenvironment of Afghanistan as inferred from dental microwear in artiodactyls. Palaeogeogr. Palaeoclim. Palaeoecol. 2004, 207, 143–163. [Google Scholar] [CrossRef]
- Merceron, G. A New Method of Dental Microwear Analysis: Application to Extant Primates and Ouranopithecus macedoniensis (Late Miocene of Greece). PALAIOS 2005, 20, 551–561. [Google Scholar] [CrossRef]
- Scott, R.S.; Ungar, P.S.; Bergstrom, T.S.; Brown, C.A.; Grine, F.E.; Teaford, M.F.; Walker, A. Dental microwear texture analysis shows within-species diet variability in fossil hominins. Nature 2005, 436, 693–695. [Google Scholar] [CrossRef] [PubMed]
- Scott, R.S.; Ungar, P.S.; Bergstrom, T.S.; Brown, C.A.; Childs, B.E.; Teaford, M.F.; Walker, A. Dental microwear texture analysis: Technical considerations. J. Hum. Evol. 2006, 51, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Ungar, P.S.; Scott, R.S.; Scott, J.R.; Teaford, M. Dental microwear analysis: Historical perspectives and new approaches. In Technique and Application in Dental Anthropology; Irish, J.D., Nelson, G.C., Eds.; Cambridge University Press: Cambridge, UK, 2008; pp. 389–425. [Google Scholar] [CrossRef]
- Ungar, P.S.; Scott, J.R.; Schubert, B.W.; Stynder, D.D. Carnivoran dental microwear textures: Comparability of carnassial facets and functional differentiation of postcanine teeth. Mammalia 2010, 74, 219–224. [Google Scholar] [CrossRef]
- Rivals, F.; Semprebon, G.; Lister, A. An examination of dietary diversity patterns in Pleistocene proboscideans (Mammuthus, Palaeoloxodon, and Mammut) from Europe and North America as revealed by dental microwear. Quat. Int. 2012, 255, 188–195. [Google Scholar] [CrossRef]
- Rivals, F.; Mol, D.; Lacombat, F.; Lister, A.M.; Semprebon, G.M. Resource partitioning and niche separation between mammoths (Mammuthus rumanus and Mammuthus meridionalis) and gomphotheres (Anancus arvernensis) in the Early Pleistocene of Europe. Quat. Int. 2015, 379, 164–170. [Google Scholar] [CrossRef]
- Semprebon, G.M.; Rivals, F.; Fahlke, J.M.; Sanders, W.J.; Lister, A.M.; Göhlich, U.B. Dietary reconstruction of pygmy mammoths from Santa Rosa Island of California. Quat. Int. 2016, 406, 123–136. [Google Scholar] [CrossRef]
- Grine, F.E. Dental evidence for dietary differences in Australopithecus and Paranthropus: A quantitative analysis of permanent molar microwear. J. Hum. Evol. 1986, 15, 783–822. [Google Scholar] [CrossRef]
- Rivals, F.; Semprebon, G.M. Dietary plasticity in ungulates: Insight from tooth microwear analysis. Quat. Int. 2011, 245, 279–284. [Google Scholar] [CrossRef]
- Semprebon, G.M.; Rivals, F. Was grass more prevalent in the pronghorn past? An assessment of the dietary adaptations of Miocene to Recent Antilocapridae (Mammalia: Artiodactyla). Palaeogeogr. Palaeoclim. Palaeoecol. 2007, 253, 332–347. [Google Scholar] [CrossRef]
- Semprebon, G.M.; Rivals, F. Trends in the paleodietary habits of fossil camels from the Tertiary and Quaternary of North America. Palaeogeogr. Palaeoclim. Palaeoecol. 2010, 295, 131–145. [Google Scholar] [CrossRef]
- King, T.; Andrews, P.; Boz, B. Effect of taphonomic processes on dental microwear. Am. J. Phys. Anthropol. 1999, 108, 359–373. [Google Scholar] [CrossRef]
- Lambert, W.D. Rediagnosis of the genus Amebelodon (Mammalia, Proboscidea, Gomphotheriidae) with a new subgenus and species. Amebelodon Britti. J. Paleont. 1990, 64, 1032–1041. [Google Scholar] [CrossRef]
- Konidaris, G.E.; Roussiakis, S.J.; Theodorou, G.E.; Koufos, G.D. The Eurasian occurrence of the shovel-tusker Konobelodon (Mammalia, Proboscidea) as illuminated by its presence in the late Miocene of Pikermi (Greece). J. Vert. Paleontol. 2014, 34, 1437–1453. [Google Scholar] [CrossRef]
- Lambert, W.D. The Fauna and Paleoecology of the Late Miocene Moss Acres Racetrack Site. Ph.D. Thesis, University of Florida, Marion, County, FL, USA, 1994. [Google Scholar]
- Shoshani, J.; Tassy, P. (Eds.) Summary, Conclusions, and a Glimpse into the Future. In The Proboscidea: Evolution and Palaeoecology of Elephants and Their Relatives; Oxford University Press: Oxford, UK, 1996; pp. 335–348. [Google Scholar]
- Lambert, W.D.; Shoshani, J. Proboscidea. In Evolution of Tertiary Mammals of North America: Terrestrial Carnivores, Ungulates, and Ungulatelike Mammals; Janis, C.M., Scott, K.M., Jacobs, L.L., Eds.; Cambridge University Press: Cambridge, UK, 1998; Volume 1, pp. 606–621. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeo. Elec. 2001, 4, 9. [Google Scholar]
- Calandra, I.; Göhlich, U.B.; Merceron, G. How could sympatric megaherbivores coexist? Example of niche partitioning within a proboscidean community from the Miocene of Europe. Die Nat. 2008, 95, 831–838. [Google Scholar] [CrossRef]
- Graham, A. Late Cenozoic evolution of tropical lowland vegetation in Veracruz, Mexico. Evolution 1975, 29, 723–735. [Google Scholar] [CrossRef]
- Leidy, J. Fossil Vertebrates from the Alachua Clays of Florida. Wagner Free Institute of Science: Philadelphia, PA, USA, 1896; Volume 4. [Google Scholar]
- Hulbert, R.C. Mixson’s Bone Bed. 2013. Available online: https://www.floridamuseum.ufl.edu/florida-vertebrate-fossils/sites/mixsons-bone-bed/ (accessed on 17 October 2022).
- Hulbert, R.C.; Whitmore, F.C. Late Miocene Mammals from the Mauvilla Local Fauna, Alabama; University of Florida: Florida, CA, USA, 2006; pp. 1–28. [Google Scholar]
- Hulbert, R.C., Jr. Calippus and Protohippus (Mammalia, Perissodactyla, Equidae) from the Miocene (Barstovian-early Hemphillian) of the Gulf Coastal Plain. Bul. Fl. Nat. Hist. Mus. 1988, 32, 221–340. [Google Scholar]
- Lambert, W.D. The osteology and paleoecology of the giant otter Enhydritherium terraenovae. J. Vertebr. Paléontol. 1997, 17, 738–749. [Google Scholar] [CrossRef]
- Webb, S.D. Rise and fall of the late Miocene ungulate fauna in North America. In Coevolution; Nitecki, M.H., Ed.; University of Chicago Press: Chicago, IL, USA, 1983; pp. 267–306. [Google Scholar]
- Zhang, H.; Wang, Y.; Janis, C.M.; Goodall, R.H.; Purnell, M.A. An examination of feeding ecology in Pleistocene proboscideans from southern China (Sinomastodon, Stegodon, Elephas), by means of dental microwear texture analysis. Quat. Int. 2017, 445, 60–70. [Google Scholar] [CrossRef]
- Lambert, W.D. The feeding habits of the shovel-tusked gomphotheres: Evidence from tusk wear patterns. Paleobiology 1992, 18, 132–147. [Google Scholar] [CrossRef]
- Pérez-Crespo, V.A.; Prado, J.L.; Alberdi, M.T.; Arroyo-Cabrales, J.; Johnson, E. Feeding ecology of the gomphotheres (Proboscidea, Gomphotheriidae) of America. Quat. Sci. Rev. 2019, 229, 106126. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locality | Museum | Number | |
|---|---|---|---|
| A. floridanus | Mixson’s Bone Bed; Alachua Formation; Todd Co. | AMNH | 37 |
| A. floridanus | Mixon’s Bone Bed; Alachua FM; Levy Co. | YPM | 3 |
| A. floridanus | Tyner Farm | UF | 4 |
| A. floridanus | McGehee Farm | UF | 1 |
| S. barbourensis | Love Bone Bed | UF | 10 |
| K. britti | Moss Acres Racetrack | UF | 3 |
| Tusks Analyzed | |||
|---|---|---|---|
| Species | Locality | Number of Upper Tusks | Number of Lower Tusks |
| A. floridanus | Tyner Farm | 5 | 3 |
| K. britti | Moss Acres | 4 | 1 |
| S. barbourensis | Love Bone Bed | 0 | 1 |
| Taxon | Locality | N | P | LP | S | % F | %C | %M | %CH | %H | %G | %PP | SWS | 0–17 | Diet |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Amebelodon floridanus | Mixson’s bone bed, Levy Co. (N = 43), Tyner Farm (N = 3), and McGehee Farm (N = 1) | 47 | 36.4 | 2.3 | 10.1 | 0 | 4.3 | 2.1 | 93.6 | 0 | 6.4 | 8.5 | 2.9 | 97.9 | B |
| Serbelodonbarbourensis | Love Bone Bed, Florida | 11 | 27.6 | 6 | 12.6 | 0 | 0 | 0 | 100 | 0 | 18.2 | 9.1 | 2.9 | 100 | B |
| Konobelodonbritti | Moss Acres Racetrack, Florida | 3 | 21.2 | 1.3 | 10.3 | 0 | 0 | 0 | 100 | 0 | 33.3 | 0 | 2.7 | 100 | B |
| Loxodonta cyclotis | Africa | 6 | 29.8 | 17 | 12.9 | 0 | 0 | 16.7 | 66.7 | 16.7 | 33.3 | 33.3 | 2.8 | 83.3 | B |
| Loxodontaafricana | Africa | 33 | 22.9 | 3.5 | 17.4 | 0 | 0 | 0 | 87.9 | 12.1 | 36.4 | 3.03 | 3.1 | 39.4 | MF |
| Elephas maximus | Southeast Asia | 10 | 20.9 | 3.7 | 18.3 | 0 | 0 | 0 | 90 | 10 | 50 | 0 | 3.1 | 40 | MF |
| Species | Locality | Number of Upper Tusks | Scratches Mostly Perpendicular to Enamel Band | Scratches Mostly Parallel to the Enamel Band | Tip Wear |
|---|---|---|---|---|---|
| A. floridanus | Tyner Farm | 5 | Yes (1 out of 5) | Yes (4 out of 5) | Yes (2 out of 5) |
| K. britti | Moss Acres | 4 | Yes (3 out of 4) | Yes (1 out of 4) | No |
| Species | Locality | Number of Lower Tusks | Dorsal and Ventral Wear Facets | Dorsal and Ventral Polishing | Tip Wear |
|---|---|---|---|---|---|
| A. floridanus | Tyner Farm | 3 | No | No | Yes (1 out of 3) |
| K. britti | Moss Acres | 1 | No | Yes | No |
| S. barbourensis | Love Bone Bed | 1 | Yes | No | Yes |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Semprebon, G.M.; Pirlo, J.; Dudek, J. Dietary Habits and Tusk Usage of Shovel-Tusked Gomphotheres from Florida: Evidence from Stereoscopic Wear of Molars and Upper and Lower Tusks. Biology 2022, 11, 1748. https://doi.org/10.3390/biology11121748
Semprebon GM, Pirlo J, Dudek J. Dietary Habits and Tusk Usage of Shovel-Tusked Gomphotheres from Florida: Evidence from Stereoscopic Wear of Molars and Upper and Lower Tusks. Biology. 2022; 11(12):1748. https://doi.org/10.3390/biology11121748
Chicago/Turabian StyleSemprebon, Gina M., Jeanette Pirlo, and Julia Dudek. 2022. "Dietary Habits and Tusk Usage of Shovel-Tusked Gomphotheres from Florida: Evidence from Stereoscopic Wear of Molars and Upper and Lower Tusks" Biology 11, no. 12: 1748. https://doi.org/10.3390/biology11121748
APA StyleSemprebon, G. M., Pirlo, J., & Dudek, J. (2022). Dietary Habits and Tusk Usage of Shovel-Tusked Gomphotheres from Florida: Evidence from Stereoscopic Wear of Molars and Upper and Lower Tusks. Biology, 11(12), 1748. https://doi.org/10.3390/biology11121748