Interactions of Ingested Polystyrene Microplastics with Heavy Metals (Cadmium or Silver) as Environmental Pollutants: A Comprehensive In Vivo Study Using Drosophila melanogaster



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. D. melanogaster Strains, Exposure, and Viability (Lethality)
2.3. Trypan Blue Staining
2.4. Interaction between PSMPLs and Intestinal Barrier Components
2.5. Detection of PSMPLs in the Hemolymph
2.6. Intracellular Oxidative Stress (Reactive Oxygen Species, ROS) Detection
2.7. Genotoxicity Induction (Comet Assay)
2.8. Statistical Analysis
3. Results
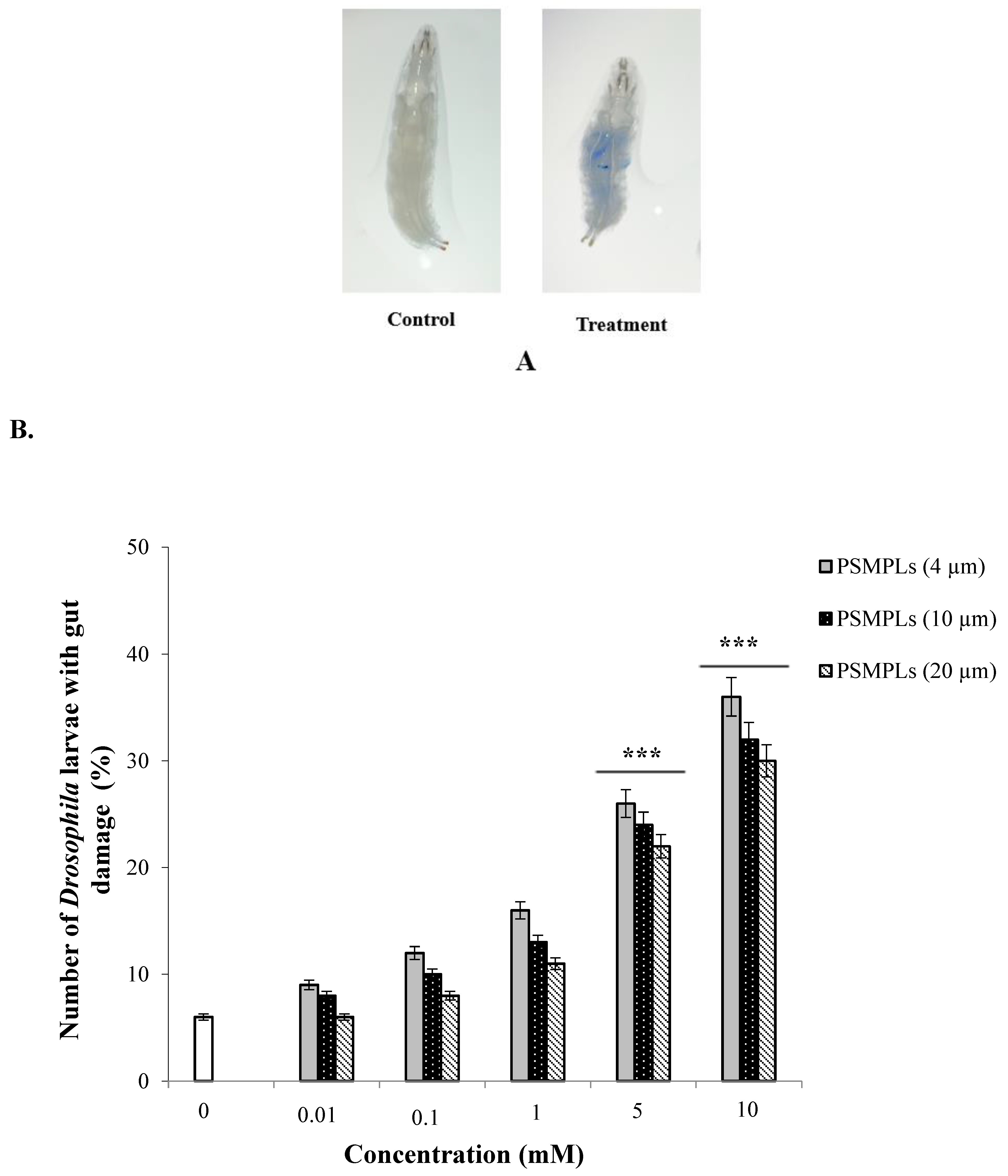
3.1. Gut Damage after Exposure to PSMPLs, CdCl2, AgNO3, and Combined Exposure
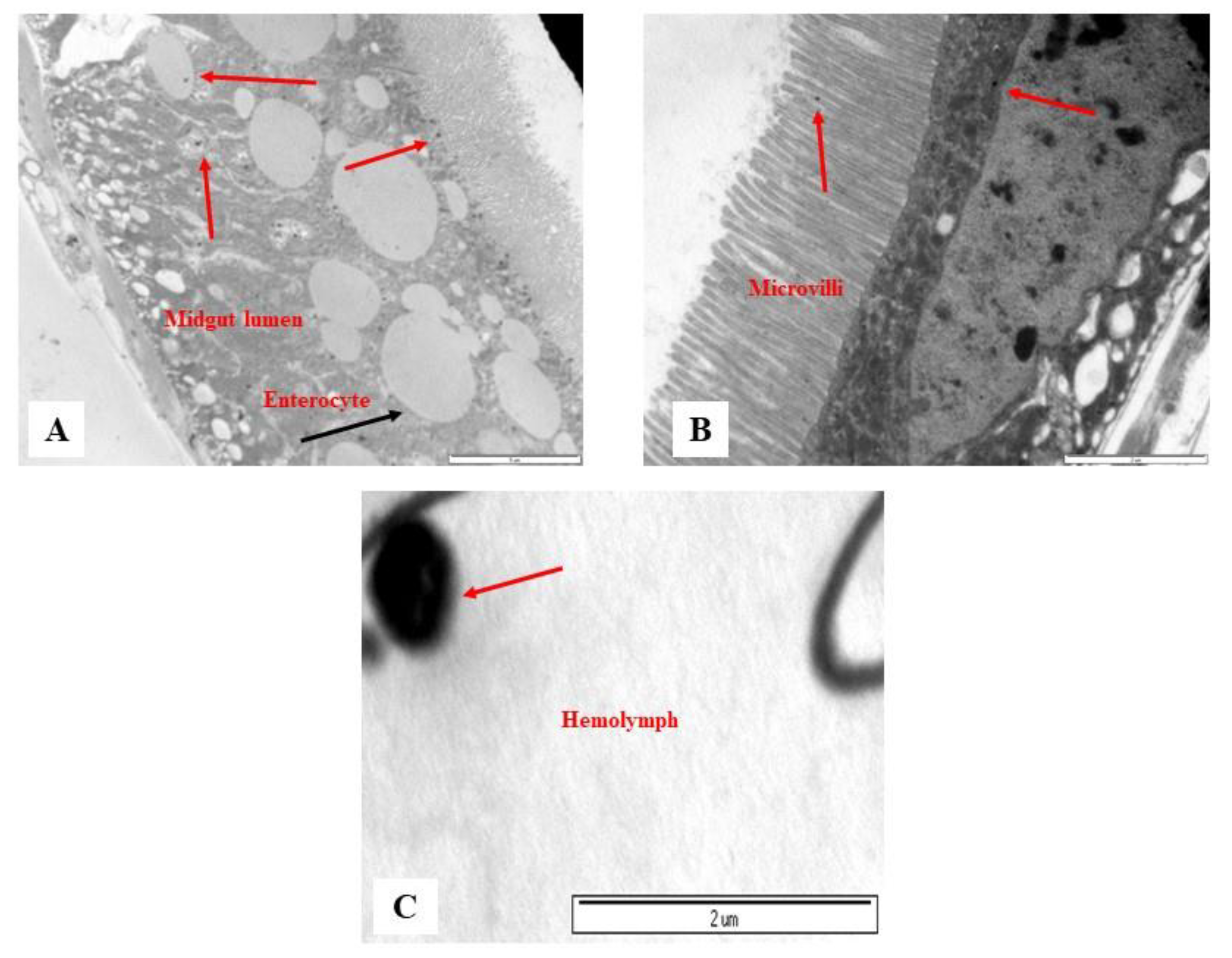
3.2. Monitoring of PSMPLs along the Intestinal Tract
3.3. Oxidative Stress Induction
3.4. Genotoxic Effects Determined by Comet Assay
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hohn, S.; Acevedo-Trejos, E.; Abrams, J.F.; Fulgencio de Moura, J.; Spranz, R.; Merico, A. The long-term legacy of plastic mass production. Sci. Total Environ. 2020, 746, 141115. [Google Scholar] [CrossRef]
- Borrelle, S.B.; Ringma, J.; Law, K.L.; Monnahan, C.C.; Lebreton, L.; McGivern, A.; Murphy, E.; Jambeck, J.; Leonard, G.H.; Hilleary, M.A.; et al. Predicted growth in plastic waste exceeds efforts to mitigate plastic pollution. Science 2020, 369, 1515–1518. [Google Scholar] [CrossRef]
- Rubio, L.; Barguilla, I.; Domenech, J.; Marcos, R.; Hernández, A. Biological effects, including oxidative stress and genotoxic damage, of polystyrene nanoparticles in different human hematopoietic cell lines. J. Hazard. Mater. 2020, 398, 122900. [Google Scholar] [CrossRef] [PubMed]
- Rubio, L.; Marcos, R.; Hernández, A. Potential adverse health effects of ingested micro- and nanoplastics on humans. Lessons learned from in vivo and in vitro mammalian models. J. Toxicol. Environ. Health B Crit. Rev. 2020, 23, 51–68. [Google Scholar] [CrossRef] [PubMed]
- Barnes, D.K.A.; Galgani, F.; Thompson, R.C.; Barlaz, M. Accumulation and fragmentation of plastic debris in global environments. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2009, 364, 1985–1998. [Google Scholar] [CrossRef] [PubMed]
- Bouwmeester, H.; Hollman, P.C.H.; Peters, R.J.B. Potential health impact of environmentally released microand nanoplastics in the human food production chain: Experiences from nanotoxicology. Environ. Sci. Technol. 2015, 49, 8932–8947. [Google Scholar] [CrossRef]
- Demir, E.; Turna Demir, F. Drosophila melanogaster as a dynamic in vivo model organism reveals the hidden effects of interactions between microplastic/nanoplastic and heavy metals. J. Appl. Toxicol. 2022, in press. [Google Scholar] [CrossRef] [PubMed]
- Domenech, J.; Cortés, C.; Vela, L.; Marcos, R.; Hernández, A. Polystyrene nanoplastics as carriers of metals. Interactions of polystyrene nanoparticles with silver nanoparticles and silver nitrate, and its effects on human intestinal Caco-2 cells. Biomolecules 2021, 11, 859. [Google Scholar] [CrossRef] [PubMed]
- Silva, B.; Bastos, A.S.; Justino, C.I.L.; Da Costa, J.P.; Duarte, A.C.; Rocha-Santos, T.A.P. Microplastics in the environment: Challenges in analytical chemistry-a review. Anal. Chim. Acta 2018, 1017, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Chae, Y.; An, Y.J. Effects of micro- and nanoplastics on aquatic ecosystems: Current research trends and perspectives. Mar. Pollut. Bull. 2017, 124, 624–632. [Google Scholar] [CrossRef] [PubMed]
- Schiavo, S.; Oliviero, M.; Chiavarini, S.; Dumontet, S.; Manzo, S. Polyethylene, Polystyrene, and Polypropylene leachate impact upon marine microalgae Dunaliella tertiolecta. J. Toxicol. Environ. Health Part A Curr. Issues 2020, 84, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Liebmann, B.; Köppel, S.; Königshofer, P.; Bucsics, T.; Reiberger, T.; Schwabl, P. Assessment of microplastic concentrations in human stool -Preliminary results of a prospective study. In Proceedings of the UEG Week 2018, Vienna, Austria, 24 October 2018. [Google Scholar]
- Cortés, C.; Domenech, J.; Salazar, M.; Pastor, S.; Marcos, R.; Hernández, A. Nanoplastics as a potential environmental health factor: Effects of polystyrene nanoparticles on human intestinal epithelial Caco-2 cells. Environ. Sci. Nano 2020, 7, 2722–2785. [Google Scholar] [CrossRef]
- Domenech, J.; Hernández, A.; Rubio, L.; Marcos, R.; Cortés, C. Interactions of polystyrene nanoplastics with in vitro models of the human intestinal barrier. Arch. Toxicol. 2020, 94, 2997–3012. [Google Scholar] [CrossRef]
- Galloway, T.; Cole, M.; Lewis, C. Interactions of microplastic debris throughout the marine ecosystem. Nat. Ecol. Evol. 2017, 1, 116. [Google Scholar] [CrossRef] [PubMed]
- Prata, J.C. Airborne microplastics: Consequences to human health? Environ. Pollut. 2018, 234, 115–126. [Google Scholar] [CrossRef] [PubMed]
- Wright, S.L.; Thompson, R.C.; Galloway, T.S. The physical impacts of microplastics on marine organisms: A review. Environ. Pollut. 2013, 178, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Rist, S.; Carney Almroth, M.; Hartmann, N.B.; Karlsson, T.M. A critical perspective on early communications concerning human health aspects of microplastics. Sci. Total Environ. 2018, 626, 720–726. [Google Scholar] [CrossRef] [PubMed]
- Wright, S.L.; Kelly, F.J. Plastic and human health: A micro issue? Environ. Sci. Technol. 2017, 51, 6634–6647. [Google Scholar] [CrossRef]
- Anbumani, S.; Kakkar, P. Ecotoxicological effects of microplastics on biota: A review. Environ. Sci. Pollut. Res. 2018, 25, 14373–14396. [Google Scholar] [CrossRef]
- Magara, G.; Elia, A.C.; Syberg, K.; Khan, F.R. Single contaminant and combined exposures of polyethylene microplastics and fluoranthene: Accumulation and oxidative stress response in the blue mussel, Mytilus edulis. J. Toxicol. Environ. Health Part A Curr. Issues 2018, 81, 761–773. [Google Scholar] [CrossRef] [PubMed]
- Sussarellu, R.; Suquet, M.; Thomas, Y.; Lambert, C.; Fabioux, C.; Pernet, M.E.; Le Goïc, N.; Quillien, V.; Mingant, C.; Epelboin, Y.; et al. Oyster reproduction is affected by exposure to polystyrene microplastics. Proc. Natl. Acad. Sci. USA 2016, 113, 2430–2435. [Google Scholar] [CrossRef] [PubMed]
- Bakircioglu, D.; Kurtulus, Y.B.; Ucar, G. Determination of some traces metal levels in cheese samples packaged in plastic and tin containers by ICP-OES after dry, wet and microwave digestion. Food Chem. Toxicol. 2011, 49, 202–207. [Google Scholar] [CrossRef] [PubMed]
- Lahimer, M.C.; Ayed, N.; Horriche, J.; Belgaied, S. Characterization of plastic packaging additives: Food contact, stability and toxicity. Arab. J. Chem. 2017, 10, 1938–1954. [Google Scholar] [CrossRef]
- Whitt, M.; Vorst, K.; Brown, W.; Baker, S.; Gorman, L. Survey of heavy metal contamination in recycled polyethylene terephthalate used for food packaging. J. Plast. Film Sheeting 2013, 29, 163–173. [Google Scholar] [CrossRef]
- Sterckeman, T.; Douay, F.; Proix, N.; Fourrier, H.; Perdix, E. Assessment of the contamination of cultivated soils by eighteen trace elements around smelters in the North of France. Water Air Soil Pollut. 2002, 135, 173–194. [Google Scholar] [CrossRef]
- Murray, K.S.; Rogers, D.T.; Kaufman, M.M. Heavy metals in an urban watershed in south eastern Michigan. J. Environ. Qual. 2004, 33, 163–172. [Google Scholar] [CrossRef]
- Alloway, B.J. Heavy Metals in Soils: Trace Metals and Metalloids in Soils and Their Bioavailability; Environmental Pollution; Springer: Dordrecht, The Netherlands, 2013. [Google Scholar]
- Abhay, K.; Prashant, P. Lead and cadmium in soft plastic toys. Curr. Sci. 2007, 93, 818–822. [Google Scholar]
- Chunhabundit, R. Cadmium exposure and potential health risk from foods in contaminated area, Thailand. Toxicol. Res. 2016, 32, 65–72. [Google Scholar] [CrossRef]
- Turner, A. Cadmium pigments in consumer products and their health risks. Sci. Total Environ. 2019, 657, 1409–1418. [Google Scholar] [CrossRef]
- Liu, H.; Liu, K.; Fu, H.; Ji, R.; Qu, X. Sunlight mediated cadmium release from colored microplastics containing cadmium pigment in aqueous phase. Environ. Pollut. 2020, 263, 114484. [Google Scholar] [CrossRef]
- Demir, E.; Vales, G.; Kaya, B.; Creus, A.; Marcos, R. Genotoxic analysis of silver nanoparticles in Drosophila. Nanotoxicology 2011, 5, 417–424. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Geller, B.; Moniz, K.; Das, P.; Chippindale, A.K.; Walker, V.K. Monitoring the developmental impact of copper and silver nanoparticle exposure in Drosophila and their microbiomes. Sci. Total Environ. 2014, 487, 822–829. [Google Scholar] [CrossRef] [PubMed]
- Holmes, L.A.; Turner, A.; Thompson, R.C. Adsorption of trace metals to plastic resin pellets in the marine environment. Environ. Pollut. 2012, 160, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Munier, B.; Bendell, L.I. Macro and micro plastics sorb and desorb metals and act as a point source of trace metals to coastal ecosystems. PLoS ONE 2018, 13, e0191759. [Google Scholar]
- Khan, F.R.; Syberg, K.; Shashoua, Y.; Bury, N.R. Influence of polyethylene microplastic beads on the uptake and localization of silver in zebrafish (Danio rerio). Environ. Pollut. 2015, 206, 73–79. [Google Scholar] [CrossRef]
- Qin, L.; Duan, Z.; Cheng, H.; Wang, Y.; Zhang, H.; Zhu, Z.; Wang, L. Size-dependent impact of polystyrene microplastics on the toxicity of cadmium through altering neutrophil expression and metabolic regulation in zebrafish larvae. Environ. Pollut. 2021, 291, 118169. [Google Scholar] [CrossRef]
- Lu, K.; Qiao, R.; An, H.; Zhang, Y. Influence of microplastics on the accumulation and chronic toxic effects of cadmium in zebrafish (Danio rerio). Chemosphere 2018, 202, 514–520. [Google Scholar] [CrossRef]
- Qiao, R.X.; Lu, K.; Deng, Y.F.; Ren, H.Q.; Zhang, Y. Combined effects of polystyrene microplastics and natural organic matter on the accumulation and toxicity of copper in zebrafish. Sci. Total Environ. 2019, 682, 128–137. [Google Scholar] [CrossRef]
- Chua, E.M.; Shimeta, J.; Nugegoda, D.; Morrison, P.D.; Clarke, B.O. Assimilation of polybrominated diphenyl ethers from microplastics by the marine amphipod, Allorchestes compressa. Environ. Sci. Technol. 2014, 48, 8127–8134. [Google Scholar] [CrossRef]
- Cho, Y.M.; Choi, K.H. The current status of studies of human exposure assessment of microplastics and their health effects: A rapid systematic review. Environ. Anal. Health Toxicol. 2021, 36, e2021004. [Google Scholar] [CrossRef]
- Yong, C.Q.Y.; Valiyaveetill, S.; Tang, B.L. Toxicity of microplastics and nanoplastics in mammalian systems. Int. J. Environ. Res. Public Health 2020, 17, 1509. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wolosker, M.B.; Zhao, Y.; Ren, H.; Lemos, B. Exposure to microplastics cause gut damage, locomotor dysfunction, epigenetic silencing, and aggravate cadmium (Cd) toxicity in Drosophila. Sci. Total Environ. 2020, 744, 140979. [Google Scholar] [CrossRef] [PubMed]
- Demir, E. Adverse biological effects of ingested polystyrene microplastics using Drosophila melanogaster as a model in vivo organism. J. Toxicol. Environ. Health Part A Curr. Issues 2021, 84, 649–660. [Google Scholar] [CrossRef]
- Jimenez-Guri, E.; Roberts, K.E.; García, F.C.; Tourmente, M.; Longdon, B.; Godley, B.J. Transgenerational effects on development following microplastic exposure in Drosophila melanogaster. Peer J. Zool. Sci. 2021, 9, e11369. [Google Scholar] [CrossRef]
- Matthews, S.; Xu, E.G.; Dumont, E.R.; Meola, V.; Pikuda, O.; Cheong, R.S.; Guo, M.; Tahara, R.; Larsson, H.C.E.; Tufenkji, N. Polystyrene micro-and nanoplastics affect locomotion and daily activity of Drosophila melanogaster. Environ. Sci. Nano 2021, 8, 110–121. [Google Scholar] [CrossRef]
- Shen, J.; Liang, B.; Zhang, D.; Li, Y.; Tang, H.; Zhong, L.; Xu, Y. Effects of PET microplastics on the physiology of Drosophila. Chemosphere 2021, 283, 131289. [Google Scholar] [CrossRef]
- Alaraby, M.; Abass, D.; Domenech, J.; Hernández, A.; Marcos, R. Hazard assessment of ingested polystyrene nanoplastics in Drosophila larvae. Environ. Sci. Nano 2022, 9, 1845–1857. [Google Scholar] [CrossRef]
- Alaraby, M.; Abass, D.; Villacorta, A.; Hernández, A.; Marcos, R. Antagonistic in vivo interaction of polystyrene nanoplastics and silver compounds. A study using Drosophila. Sci. Total Environ. 2022, 842, 156923. [Google Scholar] [CrossRef]
- Liang, B.; Zhang, D.; Liu, X.; Xu, Y.; Tang, H.; Li, Y.; Shen, J. Sex-specific effects of PET-MPs on Drosophila lifespan. Arch. Insect Biochem. Physiol. 2022, 110, e21909. [Google Scholar] [CrossRef]
- Flecknell, P. Replacement, reduction and refinement. ALTEX 2002, 19, 73–78. [Google Scholar]
- Alaraby, M.; Annangi, B.; Marcos, R.; Hernández, A. Drosophila melanogaster as a suitable in vivo model to determine potential side effects of nanomaterials: A review. J. Toxicol. Environ. Health Part B Crit. Rev. 2016, 19, 65–104. [Google Scholar] [CrossRef] [PubMed]
- Demir, E.; Turna Demir, F.; Marcos, R. Drosophila as a suitable in vivo model in the safety assessment of nanomaterials. Adv. Exp. Med. Biol. 2022, 1357, 275–301. [Google Scholar] [PubMed]
- Hartung, T.; Sabbioni, E. Alternative In Vitro Assays in Nanomaterial Toxicology. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2011, 3, 545–573. [Google Scholar] [CrossRef]
- Barguilla, I.; Domenech, J.; Rubio, L.; Marcos, R.; Hernández, A. Nanoplastics and arsenic co-exposures exacerbate oncogenic biomarkers under an in vitro long-term exposure scenario. Int. J. Mol. Sci. 2022, 23, 2958. [Google Scholar] [CrossRef] [PubMed]
- Demir, E. Mechanisms and biological impacts of graphene and multi-walled carbon nanotubes on Drosophila melanogaster: Oxidative stress, genotoxic damage, phenotypic variations, locomotor behavior, parasitoid resistance, and cellular immune response. J. Appl. Toxicol. 2022, 42, 450–474. [Google Scholar] [CrossRef]
- Turna Demir, F.; Demir, E. Exposure to boron trioxide nanoparticles and ions cause oxidative stress, DNA damage, and phenotypic alterations in Drosophila melanogaster as an in vivo model. J. Appl. Toxicol. 2022, in press. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, A.F.; Meyer, D.N.; Petriv, A.M.V.; Soto, A.L.; Shields, J.N.; Akemann, C.; Baker, B.B.; Tsou, W.L.; Zhang, Y.; Baker, T.R. Nanoplastics impact the zebrafish (Danio rerio) transcriptome: Associated developmental and neurobehavioral consequences. Environ. Pollut. 2020, 266, 115090. [Google Scholar] [CrossRef]
- Teng, M.; Zhao, X.; Wu, F.; Wang, C.; Wang, C.; White, J.C.; Zhao, W.; Zhou, L.; Yan, S.; Tian, S. Charge-specific adverse effects of polystyrene nanoplastics on zebrafish (Danio rerio) development and behavior. Environ. Int. 2022, 163, 107154. [Google Scholar] [CrossRef]
- Jewett, E.; Arnott, G.; Connolly, L.; Vasudevan, N.; Kevei, E. Microplastics and their impact on reproduction-can we learn from the C. elegans model? Front. Toxicol. 2022, 4, 748912. [Google Scholar] [CrossRef]
- Rajak, P.; Dutta, M.; Roy, S. Altered differential hemocyte count in 3rd instar larvae of Drosophila melanogaster as a response to chronic exposure of Acephate. Interdiscip. Toxicol. 2015, 8, 84–88. [Google Scholar] [CrossRef]
- Demir, E.; Kansız, S.; Doğan, M.; Topel, Ö.; Akkoyunlu, G.; Kandur, M.Y.; Turna Demir, F. Hazard Assessment of the Effects of Acute and Chronic Exposure to Permethrin, Copper Hydroxide, Acephate, and Validamycin Nanopesticides on the Physiology of Drosophila: Novel Insights into the Cellular Internalization and Biological Effects. Int. J. Mol. Sci. 2022, 23, 9121. [Google Scholar] [CrossRef] [PubMed]
- Rand, M.D. Drosophotoxicology: The growing potential for Drosophila in neurotoxicology. Neurotoxicol. Teratol. 2010, 32, 74–83. [Google Scholar] [CrossRef]
- Chifiriuc, M.C.; Ratiu, A.C.; Popa, M.; Ecovoiu, A.A. Drosophotoxicology: An emerging research area for assessing nanoparticles interaction with living organisms. Int. J. Mol. Sci. 2016, 17, 36. [Google Scholar] [CrossRef]
- Wolf, M.J.; Amrein, H.; Izatt, J.A.; Choma, M.A.; Reedy, M.C.; Rockman, H.A. From The Cover: Drosophila as a model for the identification of genes causing adult human heart disease. Proc. Natl. Acad. Sci. USA 2006, 103, 1394–1399. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, C. Drosophila melanogaster: A model and a tool to investigate malignancy and identify new therapeutics. Nat. Rev. Cancer 2013, 13, 172–183. [Google Scholar] [CrossRef] [PubMed]
- Latouche, M.; Lasbleiz, C.; Martin, E.; Monnier, V.; Debeir, T.; Mouatt-Prigent, A.; Muriel, M.P.; Morel, L.; Ruberg, M.; Brice, A.; et al. A conditional pan neuronal Drosophila model of spinocerebellar ataxia 7 with a reversible adult phenotype suitable for identifying modifier Genes. J. Neurosci. 2007, 27, 2483–2492. [Google Scholar]
- Bilen, J.; Bonini, N.M. Drosophila as a model for human neurodegenerative disease. Annu. Rev. Genet. 2005, 39, 153–171. [Google Scholar] [CrossRef]
- Moloney, A.; Sattelle, D.B.; Lomas, D.A.; Crowther, D.C. Alzheimer’s disease: Insights from Drosophila melanogaster models. Trends Biochem. Sci. 2010, 35, 228–235. [Google Scholar] [CrossRef]
- Ambegaokar, S.S.; Roy, B.; Jackson, G.R. Neurodegenerative models in Drosophila: Polyglutamine disorders, Parkinson disease, and amyotrophic lateral sclerosis. Neurobiol. Dis. 2010, 40, 29–39. [Google Scholar] [CrossRef]
- Gratz, S.J.; Ukken, F.P.; Rubinstein, C.D.; Thiede, G.; Donohue, L.K.; Cummings, A.M.; O’Connor-Giles, K.M. Highly specific and efficient CRISPR/Cas9-catalyzed homology-directed repair in Drosophila. Genetics 2014, 196, 961–971. [Google Scholar] [CrossRef] [PubMed]
- Gratz, S.J.; Rubinstein, C.D.; Harrison, M.M.; Wildonger, J.; O’Connor-Giles, K.M. CRISPR Cas9 genome editing in Drosophila. Curr. Protoc. Mol. Biol. 2015, 111, 31.2.1–31.2.20. [Google Scholar] [CrossRef]
- Meltzer, H.; Marom, E.; Alyagor, I.; Mayseless, O.; Berkun, V.; Segal-Gilboa, N.; Unger, T.; Luginbuhl, D.; Schuldiner, O. Tissue-specific (ts)CRISPR as an efficient strategy for in vivo screening in Drosophila. Nat. Commun. 2019, 10, 2113. [Google Scholar] [CrossRef] [PubMed]
- Ng, C.T.; Ong, C.N.; Yu, L.E.; Bay, B.H.; Baeg, G.H. Toxicity study of zinc oxide nanoparticles in cell culture and in Drosophila melanogaster. J. Vis. Exp. 2019, 151, e59510. [Google Scholar] [CrossRef] [PubMed]
- Sabat, D.; Patnaik, A.; Ekka, B.; Dash, P.; Mishra, M. Investigation of titania nanoparticles on behaviour and mechanosensory organ of Drosophila melanogaster. Physiol. Behav. 2016, 167, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Alaraby, M.; Demir, E.; Hernandez, A.; Marcos, R. Assessing potential harmful effects of CdSe quantum dots by using Drosophila melanogaster as in vivo model. Sci. Total Environ. 2015, 530, 66–75. [Google Scholar] [CrossRef]
- Alaraby, M.; Hernandez, A.; Annangi, B.; Demir, E.; Bach, J.; Rubio, L.; Creus, A.; Marcos, R. Antioxidant and antigenotoxic properties of CeO2 NPs and cerium sulphate: Studies with Drosophila melanogaster as a promising in vivo model. Nanotoxicology 2015, 9, 749–759. [Google Scholar] [CrossRef] [PubMed]
- Alaraby, M.; Demir, E.; Domenech, J.; Velazquez, A.; Hernandez, A.; Marcos, R. In vivo evaluation of the toxic and genotoxic effects of exposure to cobalt nanoparticles in Drosophila melanogaster. Environ. Sci. Nano 2020, 7, 610–622. [Google Scholar] [CrossRef]
- Demir, E. An in vivo study of nanorod, nanosphere, and nanowire forms of titanium dioxide using Drosophila melanogaster: Toxicity, cellular uptake, oxidative stress, and DNA damage. J. Toxicol. Environ. Health Part A Curr. Issues 2020, 83, 456–469. [Google Scholar] [CrossRef] [PubMed]
- Demir, E.; Marcos, R. Assessing the genotoxic effects of two lipid peroxidation products (4-oxo-2-nonenal and 4-hydroxy-hexenal) in haemocytes and midgut cells of Drosophila melanogaster larvae. Food Chem. Toxicol. 2017, 105, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Demir, E.; Marcos, R. Antigenotoxic potential of boron nitride nanotubes. Nanotoxicology 2018, 12, 868–884. [Google Scholar] [CrossRef]
- Irving, P.; Ubeda, J.M.; Doucet, D.; Troxler, L.; Lagueux, M.; Zachary, D.; Hoffmann, J.A.; Hetru, C.; Meister, M. New insights into Drosophila larval haemocyte functions through genome-wide analysis. Cell. Microbiol. 2005, 7, 335–3350. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.P.; McCoy, M.T.; Tice, R.R.; Schneider, E.L. A simple technique for quantitation of low levels of DNA damage in individual cells. Exp. Cell Res. 1988, 175, 184–191. [Google Scholar] [CrossRef]
- Ghosh, M.; Manivannan, J.; Sinha, S.; Chakraborty, A.; Mallick, S.K.; Bandyopadhyay, M.; Mukherjee, A. In vitro and in vivo genotoxicity of silver nanoparticles. Mutat. Res. 2012, 749, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, I.; Chowdhuri, D.K.; Bajpayee, M.; Dhawan, A. Evaluation of in vivo genotoxicity of cypermethrin in Drosophila melanogaster using the alkaline comet assay. Mutagenesis 2004, 19, 85–90. [Google Scholar] [CrossRef]
- Końca, K.; Lankoff, A.; Banasik, A.; Lisowska, H.; Kuszewski, T.; Góźdź, S.; Koza, Z.; Wojcik, A. A cross-platform public domain PC image-analysis program for the comet assay. Mutat. Res. 2003, 534, 15–20. [Google Scholar] [CrossRef]
- Turna Demir, F.; Yavuz, M. Heavy metal accumulation and genotoxic effects in levant vole (Microtus guentheri) collected from contaminated areas due to mining activities. Environ. Pollut. 2020, 256, 113378. [Google Scholar] [CrossRef]
- Turna Demir, F. In vivo effects of 1, 4-dioxane on genotoxic parameters and behavioral alterations in Drosophila melanogaster. J. Toxicol. Environ. Health Part A Curr. Issues 2022, 85, 414–430. [Google Scholar] [CrossRef] [PubMed]
- Dikilitas, M.; Kocyigit, A. Assessment of computerized and manual analysis of slides processed in single cell gel electrophoresis assay. Fresenius Environ. Bull. 2012, 21, 2981–2987. [Google Scholar]
- Von Moos, N.; Burkhardt-Holm, P.; Kohler, A. Uptake and effects of microplastics on cells and tissue of the blue mussel Mytilus edulis L. after an experimental exposure. Environ. Sci. Technol. 2012, 46, 11327–11335. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Takada, H. Microplastic fragments and microbeads in digestive tracts of planktivorous fish from urban coastal waters. Sci. Rep. 2016, 6, 8. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Zhang, Y.; Lemos, B.; Ren, H. Tissue accumulation of microplastics in mice and biomarker responses suggest widespread health risks of exposure. Sci. Rep. 2017, 7, 46687. [Google Scholar] [CrossRef] [PubMed]
- Al-Jaibachi, R.; Cuthbert, R.N.; Callaghan, A. Up and away: Ontogenic transference as a pathway for aerial dispersal of microplastics. Biol. Lett. 2018, 14, 20180479. [Google Scholar] [CrossRef] [PubMed]
- Strober, W. Trypan blue exclusion test of cell viability. Curr. Protoc. Immunol. 2001, 111, A3–B3. [Google Scholar] [CrossRef]
- Choi, J.S.; Jung, Y.J.; Hong, N.H.; Hong, S.H.; Park, J.W. Toxicological effects of irregularly shaped and spherical microplastics in a marine teleost, the sheepshead minnow (Cyprinodon variegatus). Mar. Pollut. Bull. 2018, 129, 231–240. [Google Scholar] [CrossRef]
- Avio, C.G.; Gorbi, S.; Milan, M.; Benedetti, M.; Fattorini, D.; d’Errico, G.; Pauletto, M.; Bargelloni, L.; Regoli, F. Pollutants bioavailability and toxicological risk from microplastics to marine mussels. Environ. Pollut. 2015, 198, 211–222. [Google Scholar] [CrossRef]
- Domenech, J.; Marcos, R. Pathways of human exposure to microplastics, and estimation of the total burden. Curr. Opin. Food Sci. 2021, 39, 144–151. [Google Scholar] [CrossRef]
- Poma, A.; Vecchiotti, G.; Colafarina, S.; Zarivi, O.; Aloisi, M.; Arrizza, L.; Chichiriccò, G.; Di Carlo, P. In vitro genotoxicity of polystyrene nanoparticles on the human fibroblast Hs27 cell line. Nanomaterials 2019, 9, 1299. [Google Scholar] [CrossRef]
- Capo, F.; Wilson, A.; Di Cara, F. The intestine of Drosophila melanogaster: An emerging versatile model system to study intestinal epithelial homeostasis and host-microbial interactions in humans. Microorganisms 2019, 7, 336. [Google Scholar] [CrossRef]
- Qiao, J.; Chen, R.; Wang, M.; Bai, R.; Cui, X.; Liu, Y.; Wu, C.; Chen, C. Perturbation of gut microbiota plays an important role in micro/nanoplastics-induced gut barrier dysfunction. Nanoscale 2021, 13, 8806–8816. [Google Scholar] [CrossRef]
- Douglas, A.E. Simple animal models for microbiome research. Nat. Rev. Microbiol. 2019, 17, 764–775. [Google Scholar] [CrossRef]
- Lesperance, D.N.; Broderick, N.A. Microbiomes as modulators of Drosophila melanogaster homeostasis and disease. Curr. Opin. Food Sci. 2020, 39, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Yang, J.; Wu, W.M.; Zhao, J.; Song, Y.; Gao, L.; Yang, R.; Jiang, L. Biodegradation and mineralization of polystyrene by plastic-eating mealworms: Part 2. Role of gut microorganisms. Environ. Sci. Technol. 2015, 49, 12087–12093. [Google Scholar] [CrossRef]
- Huerta Lwanga, E.; Thapa, B.; Yang, X.; Gertsen, H.; Salánki, T.; Geissen, V.; Garbeva, P. Decay of low-density polyethylene by bacteria extracted from earthworm’s guts: A potential for soil restoration. Sci. Total Environ. 2018, 624, 753–757. [Google Scholar] [CrossRef] [PubMed]
- Fournier, E.; Etienne-Mesmin, L.; Grootaert, C.; Jelsbak, L.; Syberg, K.; Blanquet-Diot, S.; Mercier-Bonin, M. Microplastics in the human digestive environment: A focus on the potential and challenges facing in vitro gut model development. J. Hazard. Mater. 2021, 415, 125632. [Google Scholar] [CrossRef]
- Lei, L.; Wu, S.; Lu, S.; Liu, M.; Song, Y.; Fu, Z.; Shi, H.; Raley-Susman, K.M.; He, D. Microplastic particles cause intestinal damage and other adverse effects in zebrafish Danio rerio and nematode Caenorhabditis elegans. Sci. Total Environ. 2018, 619–620, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Shi, Q.; Tang, J.; Liu, X.; Liu, R. Ultraviolet-induced photodegradation elevated the toxicity of polystyrene nanoplastics on human lung epithelial A549 cells. Environ. Sci. Nano 2021, 8, 2660–2675. [Google Scholar] [CrossRef]
- Sendra, M.; Saco, A.; Yeste, M.P.; Romero, A.; Novoa, B.; Figueras, A. Nanoplastics: From tissue accumulation to cell translocation into Mytilus galloprovincialis hemocytes. Resilience of immune cells exposed to nanoplastics and nanoplastics plus Vibrio splendidus combination. J. Hazard. Mat. 2020, 388, 121788. [Google Scholar] [CrossRef]
- Parenti, C.C.; Binelli, A.; Caccia, S.; Della Torre, C.; Magni, S.; Pirovano, G.; Casartelli, M. Ingestion and effects of polystyrene nanoparticles in the silkworm Bombyx mori. Chemosphere 2020, 257, 127203. [Google Scholar] [CrossRef]
- Magni, S.; Gagné, F.; André, C.; Della Torre, C.; Auclair, J.; Hanana, H.; Parenti, C.C.; Bonasoro, F.; Binelli, A. Evaluation of uptake and chronic toxicity of virgin polystyrene microbeads in freshwater zebra mussel Dreissena polymorpha (Mollusca: Bivalvia). Sci. Total Environ. 2018, 631, 778–788. [Google Scholar] [CrossRef]
- Adolfsson, K.; Abariute, L.; Dabkowska, A.P.; Schneider, M.; Häcker, U.; Prinz, C.N. Direct comparison between in vivo and in vitro microsized particle phagocytosis assays in Drosophila melanogaster. Toxicol. In Vitro 2018, 46, 213–218. [Google Scholar] [CrossRef]
- Liu, Z.; Huang, Y.; Jiao, Y.; Chen, Q.; Wu, D.; Yu, P.; Li, Y.; Cai, M.; Zhao, Y. Polystyrene nanoplastic induces ROS production and affects the MAPK-HIF-1/NFkB-mediated antioxidant system in Daphnia pulex. Aquat. Toxicol. 2020, 220, 105420. [Google Scholar] [CrossRef]
- Lin, W.; Jiang, R.; Hu, S.; Xiao, X.; Wu, J.; Wei, S.; Xiong, Y.; Ouyang, G. Investigating the toxicities of different functionalized polystyrene nanoplastics on Daphnia magna. Ecotoxicol. Environ. Saf. 2019, 80, 509–516. [Google Scholar] [CrossRef]
- Chen, H.; Hua, X.; Li, H.; Wang, C.; Dang, Y.; Ding, P.; Yu, Y. Transgenerational neurotoxicity of polystyrene microplastics induced by oxidative stress in Caenorhabditis elegans. Chemosphere 2021, 272, 129642. [Google Scholar] [CrossRef]
- Babaei, A.A.; Rafiee, M.; Khodagholi, F.; Ahmadpour, E.; Amereh, F. Nanoplastics-induced oxidative stress, antioxidant defense, and physiological response in exposed Wistar albino rats. Environ. Sci. Pollut. Res. 2022, 29, 11332–11344. [Google Scholar] [CrossRef]
- Carbone, M.; Arron, S.T.; Beutler, B.; Bononi, A.; Cavenee, W.; Cleaver, J.E.; Croce, C.M.; D’Andrea, A.; Foulkes, W.D.; Gaudino, G.; et al. Tumour predisposition and cancer syndromes as models to study gene-environment interactions. Nat. Rev. Cancer 2020, 20, 533–549. [Google Scholar] [CrossRef]
- Barzilai, A.; Yamamoto, K.I. DNA damage responses to oxidative stress. DNA Repair 2004, 3, 1109–1115. [Google Scholar] [CrossRef]
- Baiken, Y.; Kanayeva, D.; Taipakova, S.; Groisman, R.; Ishchenko, A.A.; Begimbetova, D.; Matkarimov, B.; Saparbaev, M. Role of base excision repair pathway in the processing of complex DNA damage generated by oxidative stress and anticancer drugs. Front. Cell Dev. Biol. 2021, 8, 617884. [Google Scholar] [CrossRef]
- Godoy, V.; Blázquez, G.; Calero, M.; Quesada, L.; Martín-Lara, M.A. The potential of microplastics as carriers of metals. Environ. Pollut. 2019, 255, 113363. [Google Scholar] [CrossRef]
- Hüffer, T.; Hofmann, T. Sorption of non-polar organic compounds by micro-sized plastic particles in aqueous solution. Environ. Pollut. 2016, 214, 194–201. [Google Scholar] [CrossRef]
- Massos, A.; Turner, A. Cadmium, lead, and bromine in beached microplastics. Environ. Pollut. 2017, 227, 139–145. [Google Scholar] [CrossRef]
- Prunier, J.; Maurice, L.; Perez, E.; Gigault, J.; Wickmann, P.A.C.; Davranche, M.; ter Halle, A. Trace metals in polyethylene debris from the North Atlantic subtropical gyre. Environ. Pollut. 2019, 245, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Wong, C.S.; Chen, D.; Lu, X.; Wang, F.; Zeng, E.Y. Interaction of toxic chemicals with microplastics: A critical review. Water Res. 2018, 139, 208–219. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.M.; Karim, M.R.; Zheng, X.; Li, X. Heavy metal and metalloid pollution of soil, water and foods in Bangladesh: A critical review. Int. J. Environ. Res. Public Health. 2018, 15, 2825. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Turna Demir, F.; Akkoyunlu, G.; Demir, E. Interactions of Ingested Polystyrene Microplastics with Heavy Metals (Cadmium or Silver) as Environmental Pollutants: A Comprehensive In Vivo Study Using Drosophila melanogaster. Biology 2022, 11, 1470. https://doi.org/10.3390/biology11101470
Turna Demir F, Akkoyunlu G, Demir E. Interactions of Ingested Polystyrene Microplastics with Heavy Metals (Cadmium or Silver) as Environmental Pollutants: A Comprehensive In Vivo Study Using Drosophila melanogaster. Biology. 2022; 11(10):1470. https://doi.org/10.3390/biology11101470
Chicago/Turabian StyleTurna Demir, Fatma, Gökhan Akkoyunlu, and Eşref Demir. 2022. "Interactions of Ingested Polystyrene Microplastics with Heavy Metals (Cadmium or Silver) as Environmental Pollutants: A Comprehensive In Vivo Study Using Drosophila melanogaster" Biology 11, no. 10: 1470. https://doi.org/10.3390/biology11101470
APA StyleTurna Demir, F., Akkoyunlu, G., & Demir, E. (2022). Interactions of Ingested Polystyrene Microplastics with Heavy Metals (Cadmium or Silver) as Environmental Pollutants: A Comprehensive In Vivo Study Using Drosophila melanogaster. Biology, 11(10), 1470. https://doi.org/10.3390/biology11101470