
The Interaction Effect of Laser Irradiation and 6-Benzylaminopurine Improves the Chemical Composition and Biological Activities of Linseed (Linum usitatissimum) Sprouts

 ,
,  , , ,
, , ,  ,
,  and
and 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Set-Up
2.2. Analysis of Leaf Pigments
2.3. Analysis of Mineral Contents
2.4. Analysis of Chemical Properties
2.4.1. Sugar Quantification
2.4.2. The Quantitative Estimation of Alkaloids, Saponin, Steroids, and Tannin
2.4.3. The Quantification of Total Lipid and Protein Contents
2.5. The Determination of Amino Acids
2.6. The Determination of Individual and Total Polyphenol and Flavonoid Contents
2.7. The Determination of Vitamin Contents
2.8. The Determination of Fatty Acid Compounds and Their Total Contents
2.9. Analysis of N, Ammonium, and Nitrate Contents
2.10. The Determination of Total Antioxidant Capacities
2.11. Antibacterial Activity
2.12. Statistical Analyses
3. Results
3.1. Treatment Effect on Biological and Biometric Characteristics
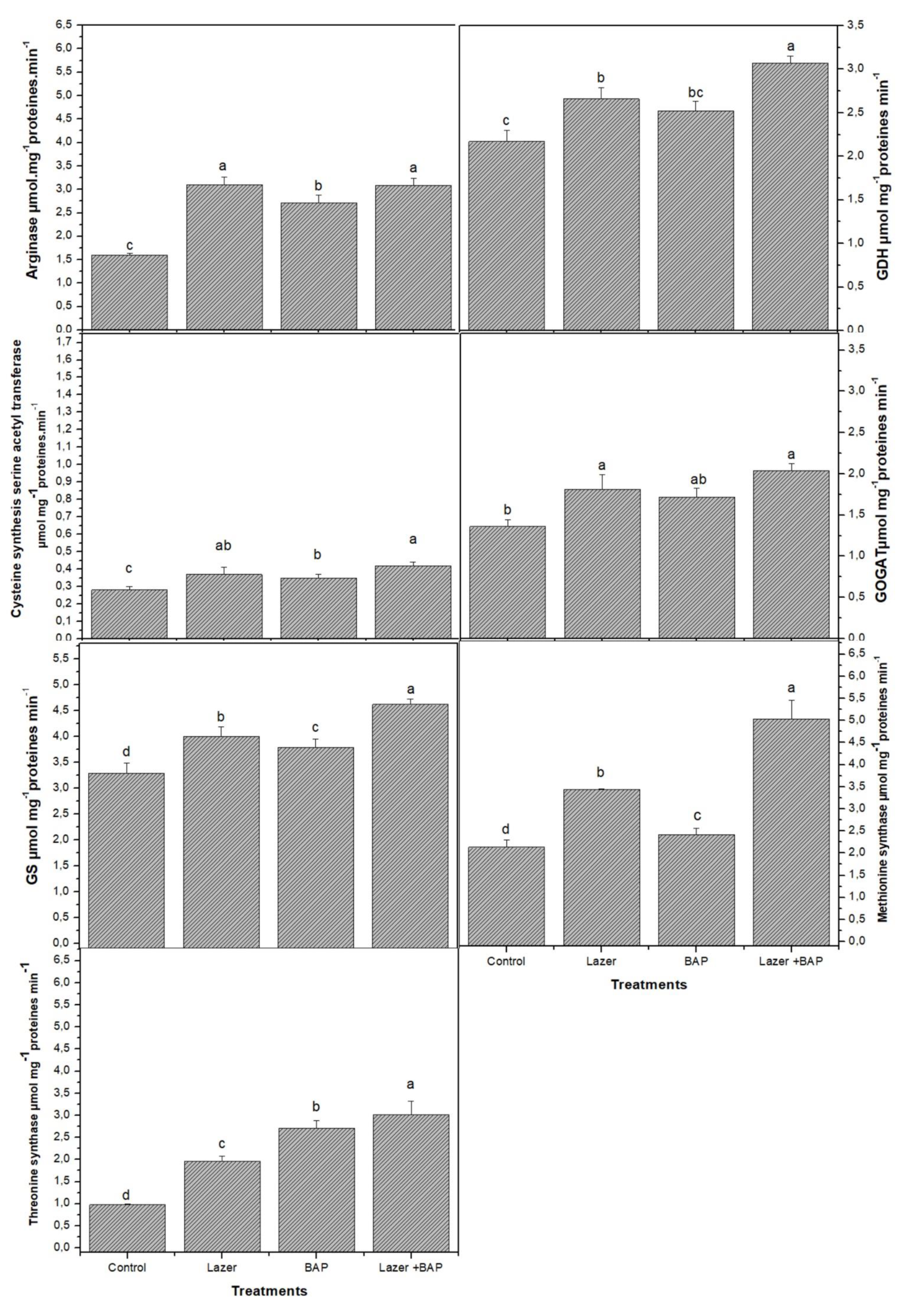
3.2. Primary Metabolites
3.3. Secondary Metabolites
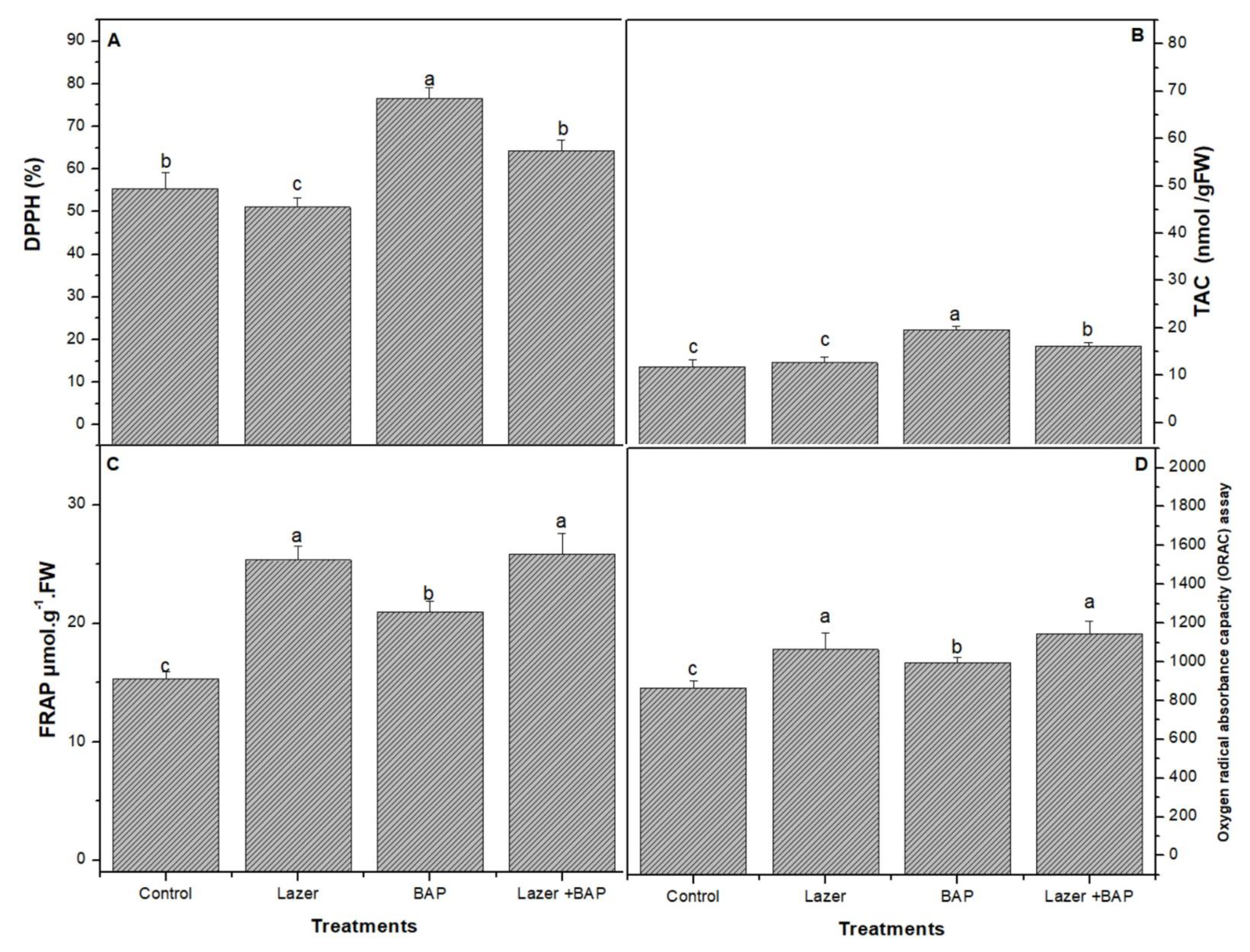
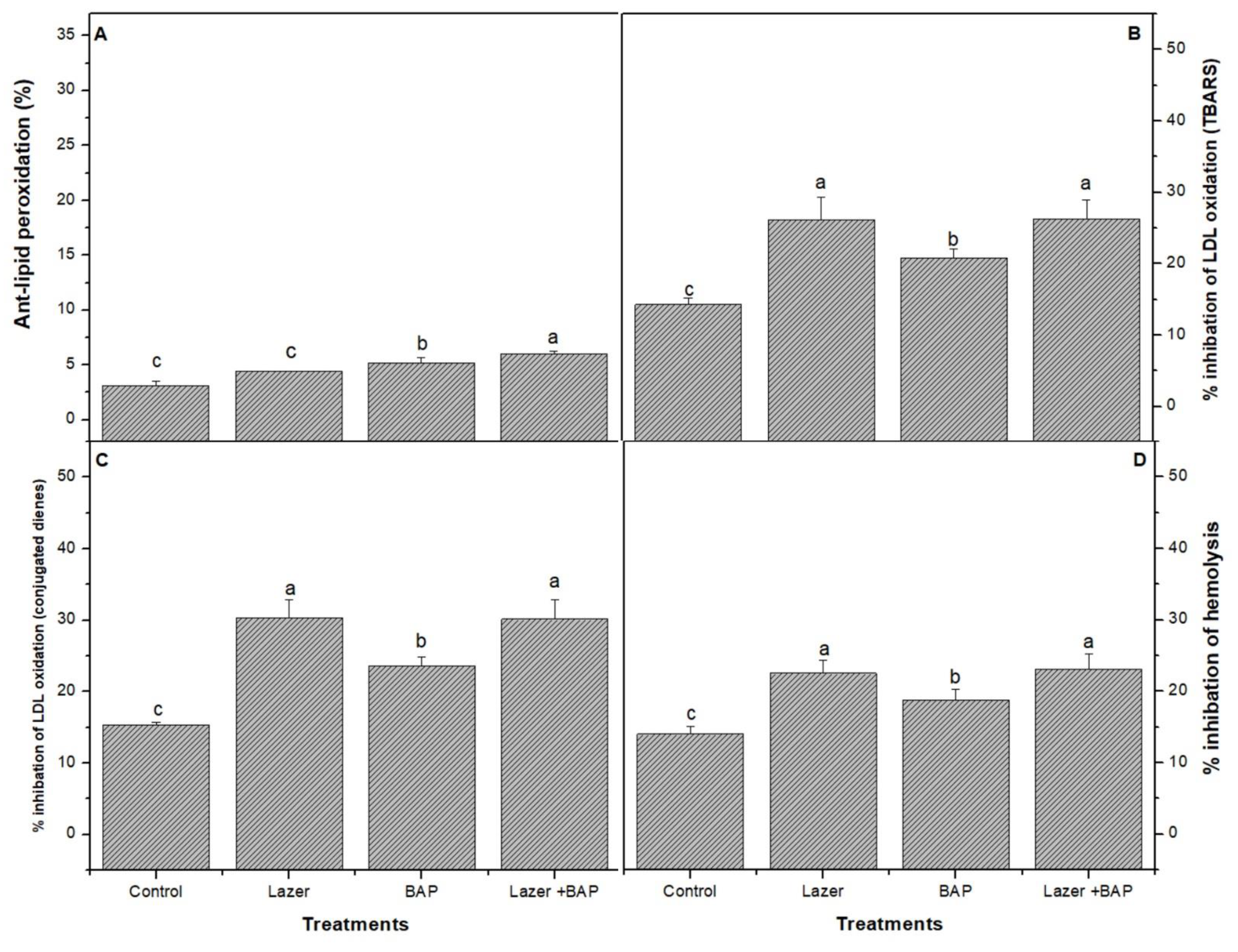
3.4. Biological Activity
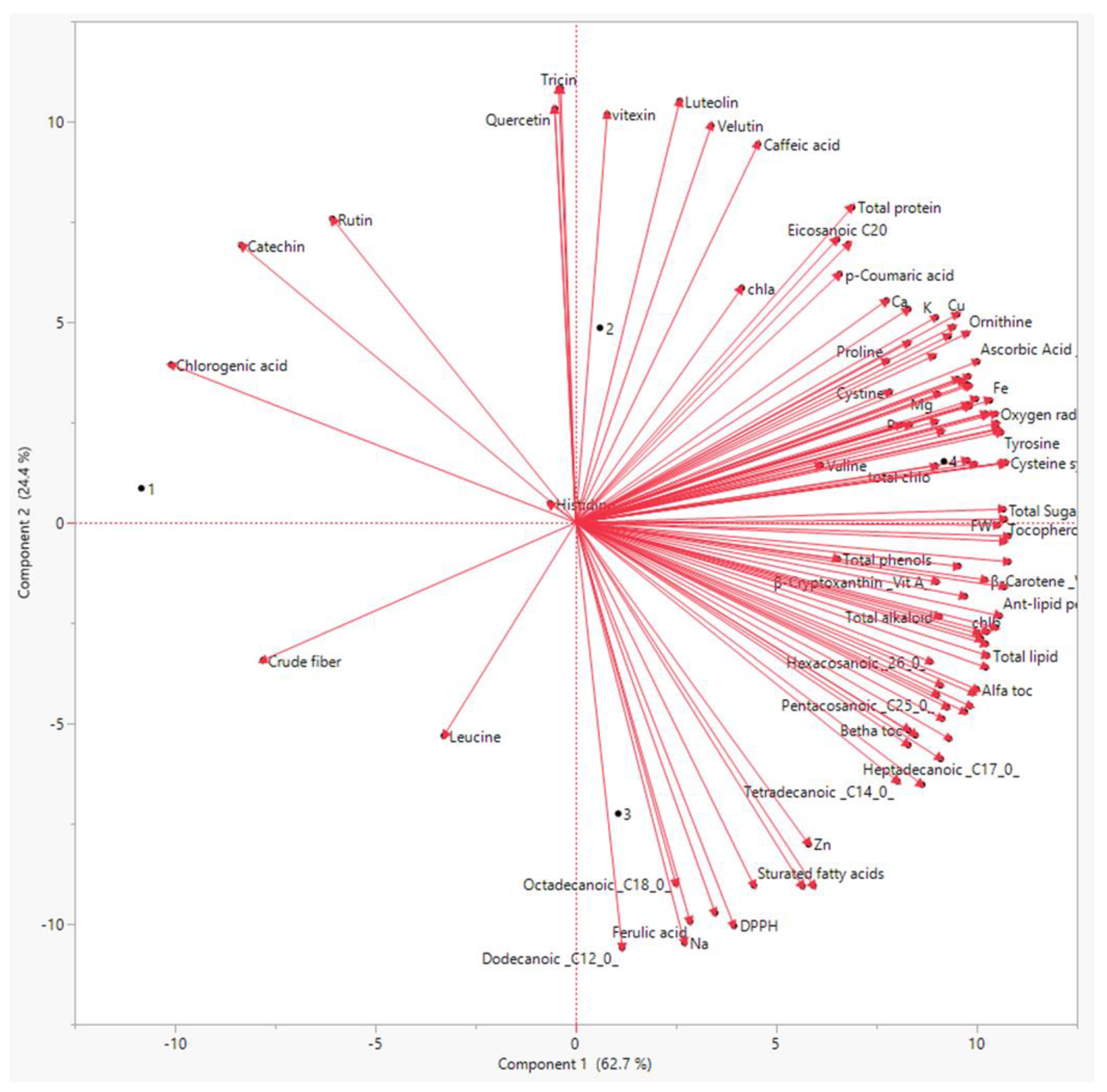
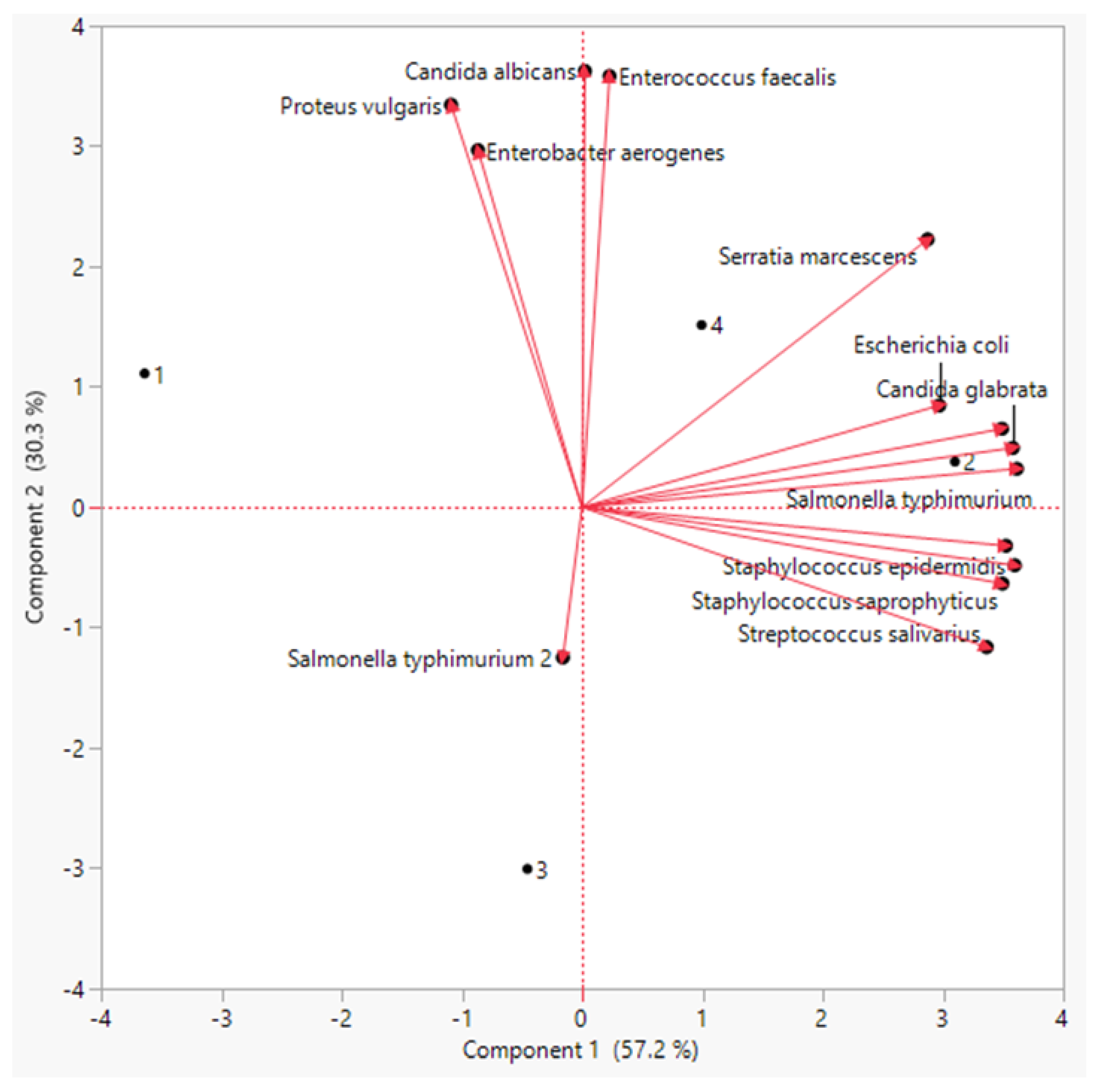
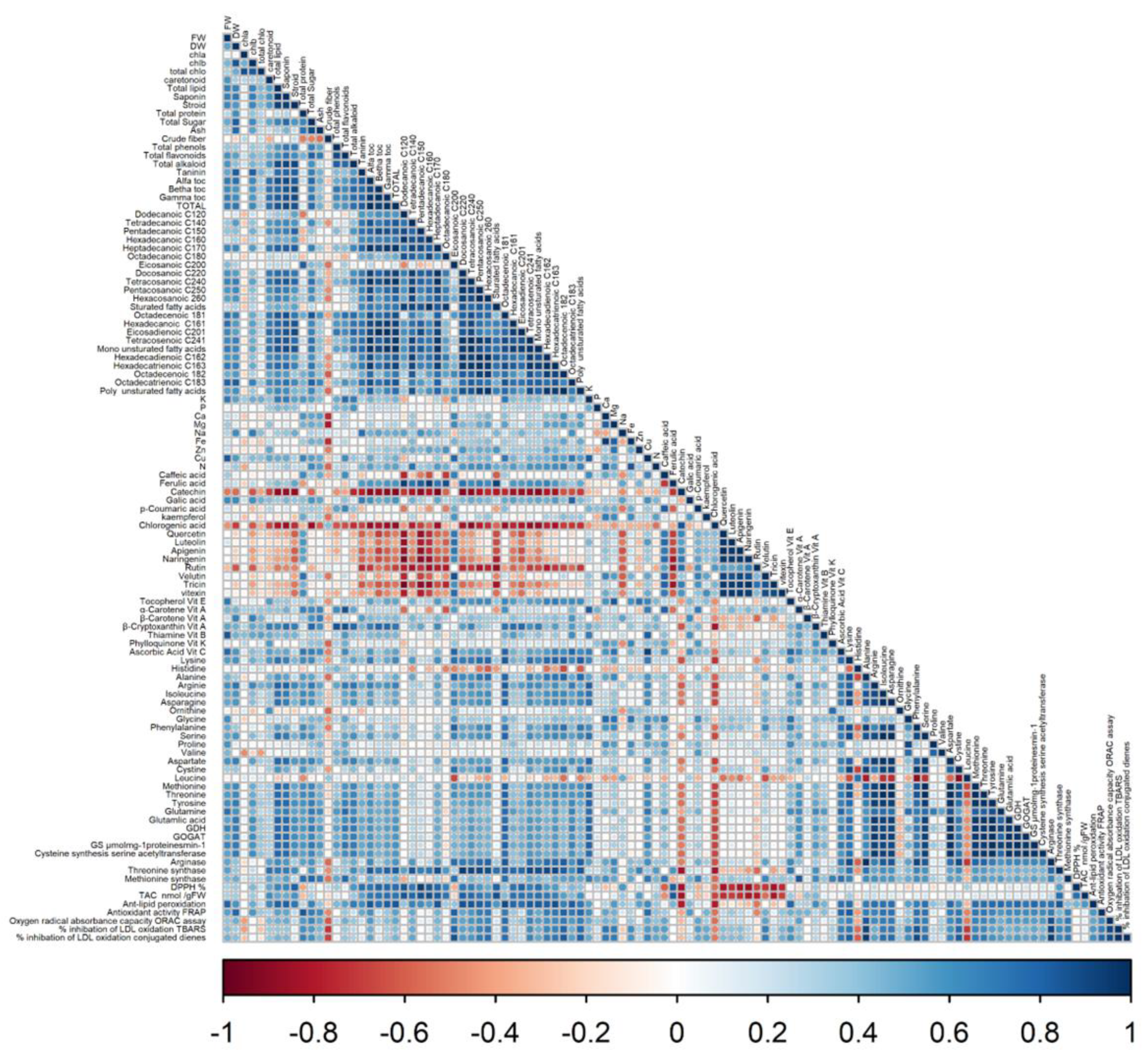
3.5. Principal Component Analysis (PCA) and Correlation Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Raj, S.; Roodbar, S.; Brinkley, C.; Wolfe, D.W. Food Security and Climate Change: Differences in Impacts and Adaptation Strategies for Rural Communities in the Global South and North. Front. Sustain. Food Syst. 2022, 5, 1–18. [Google Scholar] [CrossRef]
- Francis, G.A.; Gallone, A.; Nychas, G.J.; Sofos, J.N.; Colelli, G.; Amodio, M.L.; Spano, G. Factors Affecting Quality and Safety of Fresh-Cut Produce. Crit. Rev. Food Sci. Nutr. 2012, 52, 595–610. [Google Scholar] [CrossRef] [PubMed]
- Al-qabba, M.M.; El-mowafy, M.A.; Althwab, S.A.; Alfheeaid, H.A. Phenolic Profile, Antioxidant Activity, and Ameliorating Efficacy of Chenopodium quinoa Sprouts against CCl4-Induced Oxidative Stress in Rats. Nutrients 2020, 2, 2904. [Google Scholar] [CrossRef] [PubMed]
- Świeca, M. Elicitation with abiotic stresses improves pro-health constituents, antioxidant potential and nutritional quality of lentil sprouts. Saudi J. Biol. Sci. 2015, 22, 409–416. [Google Scholar] [CrossRef]
- Rhaman, M.S.; Imran, S.; Rauf, F.; Khatun, M.; Baskin, C.C.; Murata, Y.; Hasanuzzaman, M. Seed Priming with Phytohormones: An Effective Approach for the Mitigation of Abiotic Stress. Plants 2021, 10, 37. [Google Scholar] [CrossRef]
- Qi, Z.; Yue, M.; Wang, X.L. Laser pretreatment protects cells of broad bean from UV-B radiation damage. J. Photochem. Photobiol. B Biol. 2000, 59, 33–37. [Google Scholar] [CrossRef]
- Okla, M.K.; El-tayeb, M.A.; Qahtan, A.A.; Abdel-maksoud, M.A.; Elbadawi, Y.B.; Alaskary, M.K.; Balkhyour, M.A.; Hassan, A.H.A. Laser Light Treatment of Seeds for Improving the Biomass Photosynthesis, Chemical Composition and Biological Activities of Lemongrass Sprouts. Agronomy 2021, 11, 478. [Google Scholar] [CrossRef]
- Almuhayawi, M.S.; Hassan, A.H.A.; Al Jaouni, S.K.; Alkhalifah, D.H.M.; Hozzein, W.N.; Selim, S.; AbdElgawad, H.; Khamis, G. Influence of elevated CO2 on nutritive value and health-promoting prospective of three genotypes of Alfalfa sprouts (Medicago Sativa). Food Chem. 2021, 340, 128147. [Google Scholar] [CrossRef]
- Sarkar, D.; Kar, S.K.; Chattopadhyay, A.; Rakshit, A.; Tripathi, V.K.; Dubey, P.K.; Abhilash, P.C. Low input sustainable agriculture: A viable climate-smart option for boosting food production in a warming world. Ecol. Indic. 2020, 115, 106412. [Google Scholar] [CrossRef]
- Zrig, A. Impact of sprouting under potassium nitrate priming on nitrogen assimilation and bioactivity of three Medicago species. Plants 2022, 11, 71. [Google Scholar] [CrossRef]
- Sytar, O.; Kumari, P.; Yadav, S.; Brestic, M.; Rastogi, A. Phytohormone Priming: Regulator for Heavy Metal Stress in Plants. J. Plant Growth Regul. 2019, 38, 739–752. [Google Scholar] [CrossRef]
- Sathasivam, R.; Kim, M.C.; Chung, Y.S. Effect of cytokinins on growth and phenylpropanoid accumulation in Tartary buckwheat sprouts (Fagopyrum esculentum Moench). J. Arid. Agric. 2021, 7, 89–94. [Google Scholar] [CrossRef]
- Sun, S.; Hou, S.; Sun, Z.; Linghu, B.; Wang, Y.; Huang, K. Regeneration of buckwheat plantlets from hypocotyl and the influence of exogenous hormones on rutin content and rutin biosynthetic gene expression In Vitro. J. Arid. Agri. 2014, 3, 1159–1167. [Google Scholar] [CrossRef]
- Baenas, N.; Villaño, D.; García-Viguera, C.; Moreno, D.A. Optimizing elicitation and seed priming to enrich broccoli and radish sprouts in glucosinolates. Food Chem. 2016, 204, 314–319. [Google Scholar] [CrossRef] [PubMed]
- Okla, M.K.; Rubnawaz, S.; Dawoud, T.M.; Al-Amri, S.; El-Tayeb, M.A.; Abdel-Maksoud, M.A.; Akhtar, N.; Zrig, A.; AbdElgayed, G.; AbdElgawad, H. Laser Light Treatment Improves the Mineral Composition, Essential Oil Production and Antimicrobial Activity of Mycorrhizal Treated Pelargonium graveolens. Molecules 2022, 27, 1752. [Google Scholar] [CrossRef] [PubMed]
- Tavarini, S.; Castagna, A.; Conte, G.; Foschi, L.; Sanmartin, C.; Incrocci, L.; Ranieri, A.; Serra, A.; Angelini, L.G. Evaluation of Chemical Composition of Two Linseed Varieties as Sources of Health-Beneficial Substances. Molecules 2019, 24, 3729. [Google Scholar] [CrossRef]
- Goyal, A.; Sharma, V.; Upadhyay, N. Flax and flaxseed oil: An ancient medicine & modern functional food. J. Food Sci. Technol. 2014, 51, 1633–1653. [Google Scholar] [CrossRef]
- Han, H. Antioxidant Activity of Flaxseed (Linum usitatissimum L.) shell and Analysis of Its Polyphenol Contents by LC-MS/MS, Hasibe Yılma z and İlhami Gülçi n. Rec. Nat. Prod. 2018, 4, 397–402. [Google Scholar] [CrossRef]
- Thayer, S.S.; Bjrkman, O. Leaf Xanthophyll content and composition in sun and shade determined by HPLC. Photosynth. Res. 1990, 23, 331–343. [Google Scholar] [CrossRef]
- Abdelgawad, H.; De Vos, D.; Zinta, G.; Domagalska, M.A.; Beemster, G.T.S.; Asard, H. Grassland species differentially regulate proline concentrations under future climate conditions: An integrated biochemical and modelling approach. New Phytol. 2015, 208, 354–369. [Google Scholar] [CrossRef]
- Alasalvar, C.; Shahidi, F.; Liyanapathirana, C.M.; Ohshima, T. Turkish Tombul Hazelnut (Corylus avellana L.) 1 Compositional Characteristics. J. Agric. Food Chem. 2003, 51, 3790–3796. [Google Scholar] [CrossRef] [PubMed]
- Gomaa, N.H.; Abdelgawad, H.R. Phytotoxic effects of Echinochloa colona (L.) Link (Poaceae) extracts on the germination and seedling growth of weeds. Span. J. Agric. Res. 2012, 10, 492–501. [Google Scholar] [CrossRef]
- Torras-Claveria, L.; Berkov, S.; Codina, C.; Viladomat, F.; Bastida, J. Metabolomic analysis of bioactive Amaryllidaceae alkaloids of ornamental varieties of Narcissus by GC-MS combined with k-means cluster analysis. Ind. Crops Prod. 2014, 56, 211–222. [Google Scholar] [CrossRef]
- Kamal, J. Quantification of alkaloids, phenols and flavonoids in sunflower (Helianthus annuus L.). African J. Biotechnol. 2011, 10, 3149–3151. [Google Scholar] [CrossRef]
- Hassanpour, H.; Khavari-Nejad, R.A.; Niknam, V.; Razavi, K.; Najafi, F. Effect of penconazole and drought stress on the essential oil composition and gene expression of Mentha pulegium L. (Lamiaceae) at flowering stage. Acta Physiol. Plant. 2014, 36, 1167–1175. [Google Scholar] [CrossRef]
- Cataldo, D.A.; Haroon, M.H.; Schrader, L.E.; Youngs, V.L. Rapid colorimetric determination of nitrate in plant tissue by nitration of salicylic acid. Commun. Soil Sci. Plant Anal. 1975, 6, 71–80. [Google Scholar] [CrossRef]
- Tanha, A.; Golzardi, F.; Mostafavi, K. Seed Priming to Overcome Autotoxicity of Alfalfa (Medicago sativa). World J. Environ. Biosci. 2017, 6, 1–5. [Google Scholar]
- Alukedi, A.O.; Almarie, A.A.; Alalousi, M.A.; Farhan, S.S.; Almehemdi, A.F. Effect of laser exposure as pre sowing seed paiming in three flax cultivars (Linum usitatissimum L.). Plant Cell Biotechnol. Mol. Biol. 2021, 22, 43–52. [Google Scholar]
- Podleśny, J.; Stochmal, A.; Podleśna, A.; Misiak, L.E. Effect of laser light treatment on some biochemical and physiological processes in seeds and seedlings of white lupine and faba bean. Plant Growth Regul. 2012, 67, 227–233. [Google Scholar] [CrossRef]
- Hasan, M.; Hanafiah, M.M.; Alhilfy, I.H.H.; Taha, Z.A. Comparison of the effects of two laser photobiomodulation techniques on bio-physical properties of Zea mays L. seeds. PeerJ 2021, 9, e10614. [Google Scholar] [CrossRef]
- Sevostyanova, N.N.; Pchelina, E.A.; Gordievskaia, V.O.; Danilovskikh, M.G.; Trezorova, O.Y. Effect of laser irradiation on the processes involved in growth of mustard and radish seeds. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2020; Volume 613. [Google Scholar] [CrossRef]
- Mangena, P. Role of Benzyladenine Seed Priming on Growth and Physiological and Biochemical Response of Soybean Plants Grown under High. Int.J. Agron. 2020, 2020, 8847098. [Google Scholar] [CrossRef]
- Chaudhuri, G.; Selvaraj, U.; Babu, V.; Thilagaraj, R.W. Recent Trends in Phosphatase-Mediated Bioremediation; IntechOpen: London, UK, 2017. [Google Scholar] [CrossRef]
- Vasilevski, G. Perspective of the application of biophysical methods in sustanable agriculure. Bulg. J. Plant Physiol. 2003, 29, 179–186. [Google Scholar]
- Hernández, J.A.; Almansa, M.S. Short-term effects of salt stress on antioxidant systems and leaf water relations of pea leaves. Physiol. Plant. 2002, 115, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Pari, A.; Karalija, E.; Jasmina, Č. Growth, secondary metabolites production, antioxidative and antimicrobial activity of mint under the influence of plant growth regulators. Act. Biolog. Szeged. 2017, 61, 189–195. [Google Scholar]
- Kajla, P.; Sharma, A.; Sood, D.R. Flaxseed—A potential functional food source. J. Food Sci. Technol. 2015, 52, 1857–1871. [Google Scholar] [CrossRef]
- Marsh, Z.; Yang, T.; Nopo-olazabal, L.; Wu, S.; Ingle, T.; Joshee, N.; Medina-bolivar, F. Phytochemistry Effect of light, methyl jasmonate and cyclodextrin on production of phenolic compounds in hairy root cultures of Scutellaria lateriflora. Phytochemistry 2014, 107, 50–60. [Google Scholar] [CrossRef]
- Abbasi, B.H.; Tian, C.L.; Murch, S.J.; Saxena, P.K.; Liu, C.Z. Light-enhanced caffeic acid derivatives biosynthesis in hairy root cultures of Echinacea purpurea. Plant Cell Rep. 2007, 26, 1367–1372. [Google Scholar] [CrossRef]
- Ufaz, S.; Galili, G. Improving the Content of Essential Amino Acids in Crop Plants: Goals and Opportunities 1. Plant Physiol. 2008, 147, 954–961. [Google Scholar] [CrossRef]
- Khan, H.A.; Ayub, C.M.; Balal, R.M. Effect of seed priming with NaCl on salinity tolerance of hot pepper (Capsicum annuum L.) at seedling stage. Soil Environ. 2009, 28, 81–87. [Google Scholar]
- Teixeira, W.F.; Fagan, E.B.; Soares, L.H.; Soares, J.N.; Reichardt, K.; Neto, D.D. Seed and Foliar Application of Amino Acids Improve Variables of Nitrogen Metabolism and Productivity in Soybean Crop. Front. Plant Sci. 2018, 9, 396. [Google Scholar] [CrossRef]
- Janeczko, A.; Dziurka, M.; Ostrowska, A.; Biesaga-Koscielniak, J.; Koscielniak, J. Improving vitamin content and nutritional value of legume yield through. Legume Res. Int. J. 2015, 38, 185–193. [Google Scholar] [CrossRef]
- Perveen, R.; Jamil, Y.; Ali, Q.; Baloch, Y. He-Ne Laser-Induced Improvement in Biochemical, Physiological, Growth and Yield Characteristics in Sunflower. Photochem. Photobiol. 2011, 87, 1453–1463. [Google Scholar] [CrossRef] [PubMed]
- Asghar, T.; Jamil, Y.; Iqbal, M.; Abbas, M. Laser light and magnetic field stimulation effect on some biochemical, enzymes activity and chlorophyll contents in soybean seeds and seedlings during early growth stages. JPB 2016, 165, 283–290. [Google Scholar] [CrossRef]
- Sathasivam, R.; Ki, J.S. A review of the biological activities of microalgal carotenoids and their potential use in healthcare and cosmetic industries. Mar. Drugs 2018, 16, 26. [Google Scholar] [CrossRef] [PubMed]
- Aladjadjiyan, A. The use of physical methods for plant growing stimulation in Bulgaria. J. Cent. Eur. Agric. 2007, 8, 369–380. [Google Scholar]
- Almuhayawi, S.M.; Almuhayawi, M.S.; Al Jaouni, S.K.; Selim, S.; Hassan, A.H. Effect of Laser Light on Growth, Physiology, Accumulation of Phytochemicals, and Biological Activities of Sprouts of Three Brassica Cultivars. J. Agric. Food Chem. 2021, 69, 6240–6250. [Google Scholar] [CrossRef]
- Bongoni, R.N. Antibacterial and Antifungal activities of Linum Usitatissimum (Flax seeds). Int. J. Pharm. Educ. Res. 2016, 3, 4–8. [Google Scholar]
- Zheng, C.J.; Yoo, J.S.; Lee, T.G.; Cho, H.Y.; Kim, Y.H.; Kim, W.G. Fatty acid synthesis is a target for antibacterial activity of unsaturated fatty acids. FEBS Lett. 2005, 579, 5157–5162. [Google Scholar] [CrossRef]
- Mumtaz, S.; Mumtaz, S.; Ali, S.; Tahir, H.M.; Kazmi, S.A.R.; Mughal, T.A.; Younas, M. Evaluation of antibacterial activity of vitamin C against human bacterial pathogens. Braz. J. Biol. 2021, 83, 1–8. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | Laser | BAP | Laser + BAP | |
|---|---|---|---|---|
| FW g/plants | 1.16 ± 0.19 b | 1.51 ± 0.19 ab | 1.60 ± 0.19 ab | 2.09 ± 0.10 a |
| DW g/plants | 0.34 ± 0.03 b | 0.43 ± 0.05 a | 0.44 ± 0.03 a | 0.49 ± 0.02 a |
| Chlorophyll a (mg·g−1 FW) | 3.46 ± 0.28 ab | 3.37 ± 0.42 b | 3.21 ± 0.11 b | 3.72 ± 0.14 a |
| Chlorophyll b (mg·g−1 FW) | 1.87 ± 0.05 b | 2.06 ± 0.34 ab | 2.27 ± 0.11 a | 2.47 ± 0.15 a |
| Total Chlorophyll (mg·g−1 FW) | 5.34 ± 0.23 b | 5.43 ± 0.72 b | 5.47 ± 0.14 ab | 6.20 ± 0.26 a |
| Catenoids (mg·g−1 FW) | 0.41 ± 0.04 b | 0.44 ± 0.01 ab | 0.49 ± 0.02 ab | 0.55 ± 0.01 a |
| Control | Laser | BAP | Laser + BAP | ||
|---|---|---|---|---|---|
| Amino Acids (mg·g−1 FW) | Lysine | 1.4 ± 0.08 b | 2.63 ± 0.17 a | 2.33 ± 0.20 a | 2.67 ± 0.05 a |
| Histidine | 1.7 ± 0.08 a | 1.13 ± 0.01 b | 1.29 ± 0.10 ab | 1.71 ± 0.08 a | |
| Alanine | 0.5 ± 0.02 b | 1.22 ± 0.11 a | 0.97 ± 0.17 b | 1.12 ± 0.11 a | |
| Arginine | 1.0 ± 0.02 b | 1.17 ± 0.13 b | 1.14 ± 0.08 b | 1.37 ± 0.03 a | |
| Isoleucine | 0.1 ± 0.01 b | 0.12 ± 0.01 a | 0.10 ± 0.02 b | 0.13 ± 0.01 a | |
| Asparagine | 0.5 ± 0.06 b | 0.90 ± 0.10 a | 0.75 ± 0.12 ab | 0.90 ± 0.05 a | |
| Ornithine | 0.1 ± 0.03 b | 0.12 ± 0.02 a | 0.10 ± 0.00 b | 0.13 ± 0.02 a | |
| Glycine | 0.6 ± 0.06 b | 0.70 ± 0.09 a | 0.62 ± 0.02 b | 0.80 ± 0.02 a | |
| Phenylalanine | 0.2 ± 0.02 c | 0.32 ± 0.00 a | 0.24 ± 0.04 b | 0.30 ± 0.02 a | |
| Serine | 0.2 ± 0.02 c | 0.29 ± 0.03 a | 0.24 ± 0.04 b | 0.30 ± 0.02 a | |
| Proline | 0.6 ± 0.06 b | 0.59 ± 0.07 b | 0.56 ± 0.03 b | 0.72 ± 0.01 a | |
| Valine | 0.2 ± 0.03 b | 0.22 ± 0.04 b | 0.23 ± 0.01 b | 0.29 ± 0.01 a | |
| Aspartate | 0.01 ± 0.00 b | 0.02 ± 0.00 a | 0.02 ± 0.00 a | 0.02 ± 0.00 a | |
| Cystine | 0.01 ± 0.00 b | 0.09 ± 0.01 a | 0.05 ± 0.03 ab | 0.07 ± 0.01 a | |
| Leucine | 0.1 ± 0.02 b | 0.01 ± 0.00 c | 0.10 ± 0.05 b | 0.11 ± 0.02 a | |
| Methionine | 0.008 ± 0.00 b | 0.01 ± 0.00 a | 0.01 ± 0.00 a | 0.01 ± 0.00 a | |
| Threonine | 0.05 ± 0.01 b | 0.07 ± 0.01 ab | 0.07 ± 0.01 ab | 0.08 ± 0.00 a | |
| Tyrosine | 0.5 ± 0.05 c | 0.65 ± 0.07 a | 0.60 ± 0.06 ab | 0.73 ± 0.04 a | |
| Glutamine | 46.5 ± 4.9 c | 53.32 ± 0.49 b | 50.3 ± 2.3 b | 62.53 ± 1.36 a | |
| Glutamic acid | 34.0 ± 3.63 c | 45.19 ± 7.7 ab | 43.00 ± 4.84 b | 50.90 ± 3.56 a | |
| Vitamins (mg·g−1 FW) | α-Carotene (Vit A) | 0.4 ± 0.05 c | 0.52 ± 0.08 b | 0.23 ± 0.01 c | 0.82 ± 0.12 a |
| β-Carotene (Vit A) | 0.1 ± 0.04 b | 0.16 ± 0.04 b | 0.20 ± 0.06 ab | 0.26 ± 0.06 a | |
| β-Cryptoxanthin (Vit A) | 0.06 ± 0.01 c | 0.06 ± 0.03 c | 0.11 ± 0.01 b | 0.19 ± 0.03 a | |
| Thiamine (Vit B) | 0.05 ± 0.01 b | 0.07 ± 0.02 b | 0.06 ± 0.02 b | 0.16 ± 0.08 a | |
| Ascorbic acid (Vit C) | 1.30 ± 0.03 b | 1.96 ± 0.06 a | 1.60 ± 0.18 b | 2.40 ± 0.15 a | |
| Phylloquinone (Vit K) | 0.12 ± 0.03 b | 0.14 ± 0.04 ab | 0.11 ± 0.01 b | 0.23 ± 0.05 a | |
| α-Tocopherol (Vit E) | 187 ± 84 b | 241.8 ± 10.5 ab | 302.56 ± 6.03 a | 323.16 ± 7 a | |
| β-Tocopherol (Vit E) | 74.8 ± 0.23 b | 73.0 ± 1.8 b | 96.66 ± 0.48 a | 105.81 ± 4.4 a | |
| γ- Tocopherol (Vit E) | 15.00 ± 0.17 b | 15.96 ± 0.02 b | 20.5 ± 0.15 ab | 22.2 ± 0.84 a | |
| Tocopherol (Vit E) | 0.5 ± 0.08 c | 0.91 ± 0.18 a | 0.89 ± 0.19 b | 1.07 ± 0.04 a | |
| Fatty Acids (mg·g−1 FW) | Dodecanoic acid (C12:0) | 0.18 ± 0.01 b | 0.18 ± 0.00 b | 0.23 ± 0.00 a | 0.19 ± 0.00 b |
| Tetradecanoic acid (C14:0) | 0.16 ± 0.00 b | 0.2 ± 0.00 b | 0.24 ± 0.02 a | 0.21 ± 0.01 a | |
| Pentadecanoic acid (C15:0) | 0.02 ± 0.00 ab | 0.02 ± 0.00 ab | 0.03 ± 0.00 a | 0.03 ± 0.00 a | |
| Hexadecanoic acid (C16:0) | 10.02 ± 0.12 b | 10.54 ± 0.01 b | 13.7 ± 0.10 a | 10.98 ± 0.05 b | |
| Heptadecanoic acid (C17:0) | 0.03 ± 0.00 b | 0.03 ± 0.00 b | 0.04 ± 0.00 a | 0.04 ± 0.00 a | |
| Octadecanoic acid (C18:0) | 9.84 ± 0.23 c | 10.29 ± 0.31 b | 11.45 ± 0.32 a | 10.07 ± 0.29 c | |
| Eicosanoic acid(C20:0) | 1.11 ± 0.04 b | 1.40 ± 0.04 a | 1.16 ± 0.01 b | 1.29 ± 0.04 a | |
| Docosanoic acid (C22:0) | 1.16 ± 0.01 b | 1.23 ± 0.01 b | 1.31 ± 0.02 a | 1.30 ± 0.03 a | |
| Tetracosanoic acid (C24:0) | 1.48 ± 0.03 c | 1.67 ± 0.01 b | 1.87 ± 0.03 a | 1.90 ± 0.04 a | |
| Pentacosanoic acid (C25:0) | 0.09 ± 0.00 c | 0.12 ± 0.00 b | 0.14 ± 0.00 a | 0.13 ± 0.00 ab | |
| Hexacosanoic acid (26:0) | 0.02 ± 0.00 b | 0.02 ± 0.00 b | 0.03 ± 0.00 a | 0.02 ± 0.00 b | |
| Octadecenoic acid (18:1) | 30.86 ± 0.60 c | 41.51 ± 2.31 b | 43.17 ± 2.39 b | 52.61 ± 2.29 a | |
| Hexadecanoic acid (C16:1) | 0.42 ± 0.01 a | 0.43 ± 0.02 a | 0.5 ± 0.01 a | 0.50 ± 0.01 a | |
| Eicosadienoic acid (C20:1) | 0.22 ± 0.00 b | 0.22 ± 0.01 b | 0.29 ± 0.01 a | 0.32 ± 0.01 a | |
| Tetracosenoic acid (C24:1) | 0.29 ± 0.01 b | 0.42 ± 0.03 ab | 0.51 ± 0.03 a | 0.52 ± 0.01 a | |
| Hexadecatrienoic acid (C16:2) | 0.90 ± 0.01 b | 1.22 ± 0.03 b | 1.35 ± 0.01 a | 1.28 ± 0.04 ab | |
| Hexadecatrienoic acid (C16:3) | 0.11 ± 0.00 b | 0.14 ± 0.00 a | 0.16 ± 0.00 a | 0.16 ± 0.01 a | |
| Octadecenoic acid (18:2) | 18.93 ± 0.56 b | 24.45 ± 0.77 a | 24.37 ± 0.93 a | 24.44 ± 1.12 a | |
| Octadecatrienoic acid (C18:3) | 44.37 ± 1.34 c | 56.66 ± 4.07 c | 61.43 ± 8.82 b | 71.18 ± 3.61 a | |
| Total fatty acids | 277.6 ± 5.14 c | 330.8 ± 12.0 b | 419.7 ± 5.4 a | 451.2 ± 12.2 a | |
| Saturated fatty acids | 24.1 ± 0.21 b | 25.67 ± 0.35 ab | 30.19 ± 0.21 a | 26.1 ± 0.37 ab | |
| Monounsaturated fatty acids | 0.92 ± 0.01 b | 1.06 ± 0.02 b | 1.28 ± 0.04 ab | 1.34 ± 0.03 a | |
| Polyunsaturated fatty acids | 35.99 ± 0.97 b | 46.07 ± 1.38 a | 47.00 ± 1.46 a | 46.97 ± 1.86 a | |
| Total lipide | 139.0 ± 12.8 b | 175.5 ± 16.9 b | 210.7 ± 20.5 a | 234.4 ± 7.0 a | |
| Total protein | 131.1 ± 12.9 b | 261.1 ± 25.1 a | 137.0 ± 22.7 b | 236.7 ± 9.2 a | |
| Total Sugar | 199.7 ± 27.8 b | 317.0 ± 30.5 a | 328.2 ± 39.8 a | 470.1 ± 43.1 a | |
| Crude fiber | 3.7 ± 0.44 a | 2.5 ± 0.24 b | 3.1 ± 0.27 a | 2.9 ± 0.53 b |
| Mg·g−1 FW | Control | Laser | BAP | Laser + BAP |
|---|---|---|---|---|
| Steroid | 117.0 ± 11.07 b | 114.6 ± 11.04 b | 172.8 ± 12.83 a | 190.2 ± 7.53 a |
| Tannin | 52.7 ± 3.63 b | 58.4 ± 5.63 b | 73.8 ± 5.40 a | 71.9 ± 3.44 a |
| Saponin | 11.3 ± 0.96 c | 13.7 ± 1.31 c | 16.8 ± 1.11 b | 19.8 ± 0.40 a |
| Total alkaloid | 25.8 ± 3.80 b | 26.3 ± 2.53 b | 34.6 ± 5.28 a | 45.8 ± 1.84 a |
| Caffeic acid | 4.18 ± 0.34 c | 5.37 ± 0.55 b | 2.55 ± 0.21 d | 6.01 ± 0.44 a |
| Ferulic acid | 0.05 ± 0.00 c | 0.08 ± 0.01 b | 0.32 ± 0.01 a | 0.10 ± 0.04 b |
| Galic acid | 5.18 ± 0.55 b | 6.57 ± 0.14 b | 6.93 ± 1.52 b | 8.48 ± 0.66 a |
| p-Coumaric acid | 1.59 ± 0.05 b | 1.70 ± 0.17 ab | 1.30 ± 0.12 b | 2.31 ± 0.34 a |
| Chlorogenic acid | 0.17 ± 0.01 a | 0.13 ± 0.01 a | 0.10 ± 0.01 b | 0.09 ± 0.02 b |
| Total phenols (TPC) | 12.6 ± 0.88 b | 9.4 ± 0.90 c | 12.9 ± 0.64 b | 20.2 ± 2.00 a |
| Catechin | 1.81 ± 0.06 a | 1.52 ± 0.15 a | 0.78 ± 0.13 b | 0.95 ± 0.02 b |
| kaempferol | 0.94 ± 0.09 b | 1.37 ± 0.14 a | 1.10 ± 0.24 b | 1.30 ± 0.21 a |
| Quercetin | 2.33 ± 0.11 b | 3.57 ± 0.36 a | 1.11 ± 0.10 c | 2.25 ± 0.06 b |
| Luteolin | 0.08 ± 0.00 b | 0.12 ± 0.01 a | 0.04 ± 0.00 c | 0.10 ± 0.02 ab |
| Apigenin | 0.24 ± 0.01 ab | 0.37 ± 0.04 a | 0.12 ± 0.01 b | 0.23 ± 0.01 a |
| Naringenin | 1.72 ± 0.06 a | 2.23 ± 0.23 a | 0.69 ± 0.03 b | 1.75 ± 0.07 a |
| Rutin | 1.63 ± 0.07 a | 1.26 ± 0.13 ab | 0.78 ± 0.07 b | 1.23 ± 0.14 ab |
| Velutin | 0.01 ± 0.00 ab | 0.02 ± 0.00 a | 0.00 ± 0.00 b | 0.02 ± 0.00 a |
| Tricin | 1.39 ± 0.06 a | 1.72 ± 0.18 a | 0.52 ± 0.07 b | 1.42 ± 0.05 a |
| vitexin | 0.68 ± 0.03 b | 1.01 ± 0.10 a | 0.46 ± 0.04 b | 0.73 ± 0.04 ab |
| Total flavonoids (TFs) | 0.3 ± 0.02 b | 0.3 ± 0.03 b | 0.3 ± 0.01 b | 0.8 ± 0.05 a |
| mg/g−1 DW | Control | Laser | BAP | Laser + BAP |
|---|---|---|---|---|
| Ash | 1.6 ± 0.47 b | 2.3 ± 0.22 a | 1.9 ± 0.49 b | 3.2 ± 0.25 a |
| N | 28.95 ± 4.09 b | 40.90 ± 6.62 b | 46.02 ± 8.45 ab | 49.79 ± 6.55 a |
| P | 5.75 ± 0.85 b | 5.80 ± 0.03 b | 5.74 ± 1.58 b | 7.48 ± 0.26 a |
| K | 11.98 ± 0.86 c | 16.31 ± 1.47 b | 12.39 ± 1.82 c | 21.73 ± 1.80 a |
| Ca | 1.78 ± 0.50 d | 4.00 ± 0.93 a | 2.55 ± 0.54 c | 3.41 ± 0.71 b |
| Mg | 0.70 ± 0.27 b | 2.99 ± 0.76 a | 2.01 ± 0.88 a | 2.67 ± 0.85 a |
| Na | 0.37 ± 0.07 b | 0.25 ± 0.27 b | 0.79 ± 0.26 a | 0.49 ± 0.21 ab |
| Fe | 0.16 ± 0.01 c | 0.23 ± 0.05 b | 0.20 ± 0.02 b | 0.28 ± 0.06 a |
| Zn | 0.04 ± 0.00 b | 0.04 ± 0.01 b | 0.06 ± 0.01 a | 0.05 ± 0.01 ab |
| Cu | 0.08 ± 0.01 d | 0.19 ± 0.01 b | 0.12 ± 0.02 c | 0.23 ± 0.04 a |
| Control | Laser | BAP | Laser + BAP | ||
|---|---|---|---|---|---|
| Diameter of Inhibition Zones (mm) | |||||
| Gram-positive | Staphylococcus saprophyticus | 13.1 ± 0.7 c | 21.8 ± 2.53 a | 19.4 ± 2.8 b | 20.6 ± 1.8 a |
| Staphylococcus epidermidis | 8.9 ± 0.54 c | 19.22 ± 1.96 a | 15.1 ± 4.4 b | 16.4 ± 1.20 b | |
| Enterococcus faecalis | 16.6 ± 1.6 a | 16.26 ± 0.79 a | 11.0 ± 3.1 b | 16.6 ± 1.51 a | |
| Streptococcus salivarius | 6.9 ± 0.39 b | 14.26 ± 2.89 a | 13.0 ± 5.0 a | 12.9 ± 2.7 ab | |
| Gram-negative | Escherichia coli | 6.1 ± 0.39 b | 10.31 ± 11.03 a | 6.17 ± 0.9 b | 8.5 ± 3.81 a |
| SalmonellaentericaTyphimurium | 9.3 ± 0.9 c | 19.95 ± 4.42 a | 13.2 ± 3.7 b | 16.1 ± 2.9 ab | |
| Pseudomonas aeruginosa | 19.9 ± 1.1 c | 26.6 ± 2.04 a | 22.8 ± 3.3 b | 26.2 ± 1.5 a | |
| Proteus vulgaris | 21.3 ± 4.0 a | 18.8 ± 2.5 b | 17.0 ± 0.3 b | 21.6 ± 0.6 a | |
| Enterobacter aerogenes | 15.9 ± 1.0 a | 14.53 ± 0.93 b | 14.07 ± 1.0 b | 16.8 ± 0.4 a | |
| Serratia marcescens | 6.4 ± 0.78 b | 8.21 ± 1.39 a | 6.04 ± 0.03 b | 7.84 ± 0. ab | |
| Salmonella typhimurium | 14.7 ± 1.4 b | 14.00 ± 2.34 b | 17.16 ± 0.1 a | 17.3 ± 1.3 a | |
| Fungus | Candida albicans | 12.1 ± 2.9 a | 11.65 ± 1.82 ab | 9.58 ± 1.4 b | 12.65 ± 0.9 a |
| Candida glabrata | 3.6 ± 0.4 b | 8.19 ± 2.03 a | 5.10 ± 1.5 b | 6.43 ± 1.2 ab | |
| Aspergillus flavus | 11.2 ± 1.7 b | 17.0 ± 1.96 a | 15.0 ± 2.2 ab | 16.4 ± 1.27 a | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zrig, A.; Najar, B.; Magdy Korany, S.; Hassan, A.H.A.; Alsherif, E.A.; Ali Shah, A.; Fahad, S.; Selim, S.; AbdElgawad, H. The Interaction Effect of Laser Irradiation and 6-Benzylaminopurine Improves the Chemical Composition and Biological Activities of Linseed (Linum usitatissimum) Sprouts. Biology 2022, 11, 1398. https://doi.org/10.3390/biology11101398
Zrig A, Najar B, Magdy Korany S, Hassan AHA, Alsherif EA, Ali Shah A, Fahad S, Selim S, AbdElgawad H. The Interaction Effect of Laser Irradiation and 6-Benzylaminopurine Improves the Chemical Composition and Biological Activities of Linseed (Linum usitatissimum) Sprouts. Biology. 2022; 11(10):1398. https://doi.org/10.3390/biology11101398
Chicago/Turabian StyleZrig, Ahlem, Basma Najar, Shereen Magdy Korany, Abdelrahim H. A. Hassan, Emad A. Alsherif, Anis Ali Shah, Shah Fahad, Samy Selim, and Hamada AbdElgawad. 2022. "The Interaction Effect of Laser Irradiation and 6-Benzylaminopurine Improves the Chemical Composition and Biological Activities of Linseed (Linum usitatissimum) Sprouts" Biology 11, no. 10: 1398. https://doi.org/10.3390/biology11101398
APA StyleZrig, A., Najar, B., Magdy Korany, S., Hassan, A. H. A., Alsherif, E. A., Ali Shah, A., Fahad, S., Selim, S., & AbdElgawad, H. (2022). The Interaction Effect of Laser Irradiation and 6-Benzylaminopurine Improves the Chemical Composition and Biological Activities of Linseed (Linum usitatissimum) Sprouts. Biology, 11(10), 1398. https://doi.org/10.3390/biology11101398