
Response of Aquatic Organisms Communities to Global Climate Changes and Anthropogenic Impact: Evidence from Listvennichny Bay of Lake Baikal

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Site Description
2.1. Habitat Characteristics: Natural and Climatic Factors
2.2. Anthropogenic Impact
3. Materials and Methods
3.1. Field Studies
Water Chemistry
3.2. Sampling of Plankton
3.3. Sampling of Benthos
3.4. Laboratory Analyses
3.5. Data Analyses
3.5.1. Communities of Aquatic Organisms
3.5.2. Assessment of Environmental Quality of the Studied Sites According to Biological Indicators
4. Results
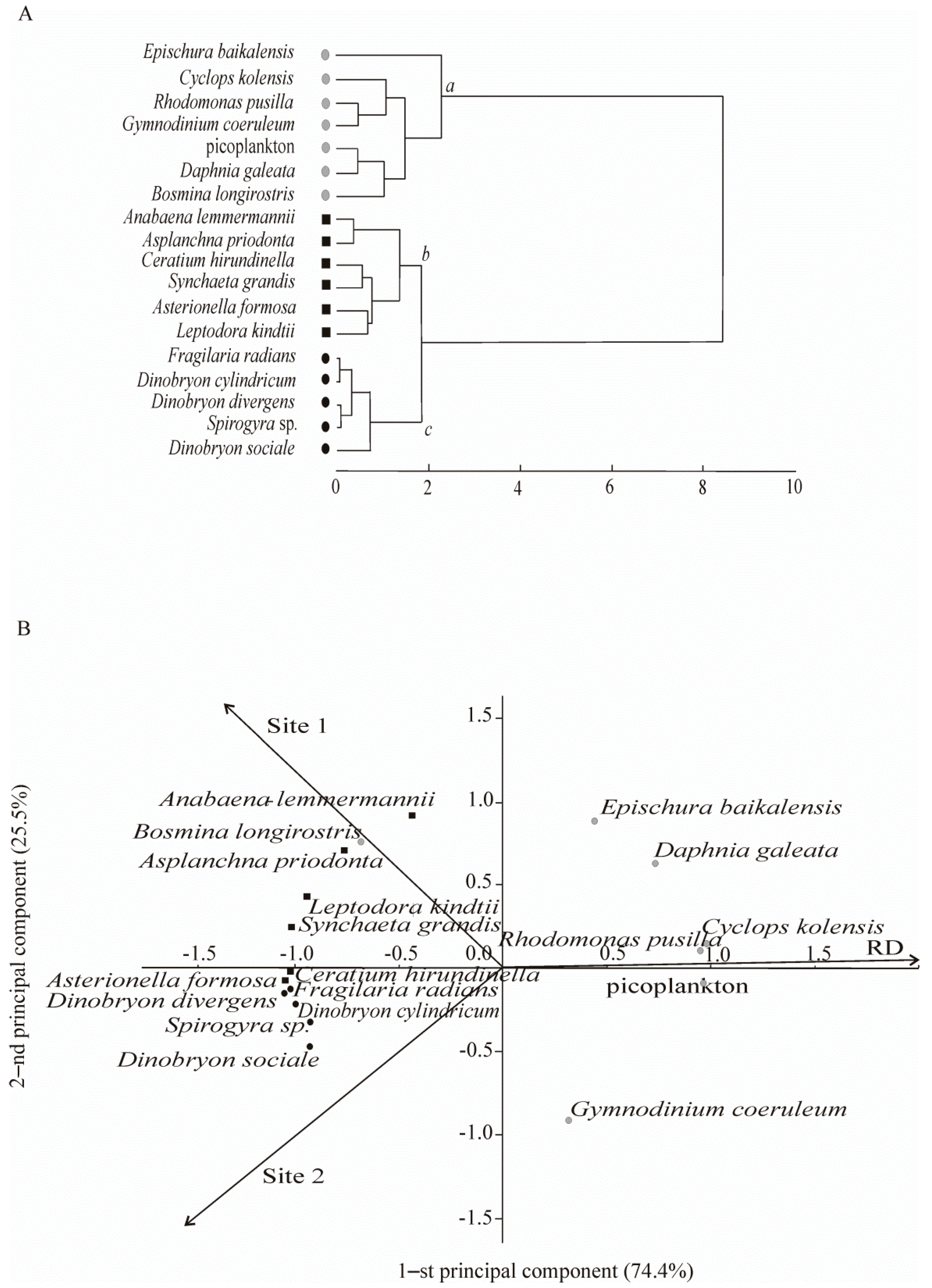
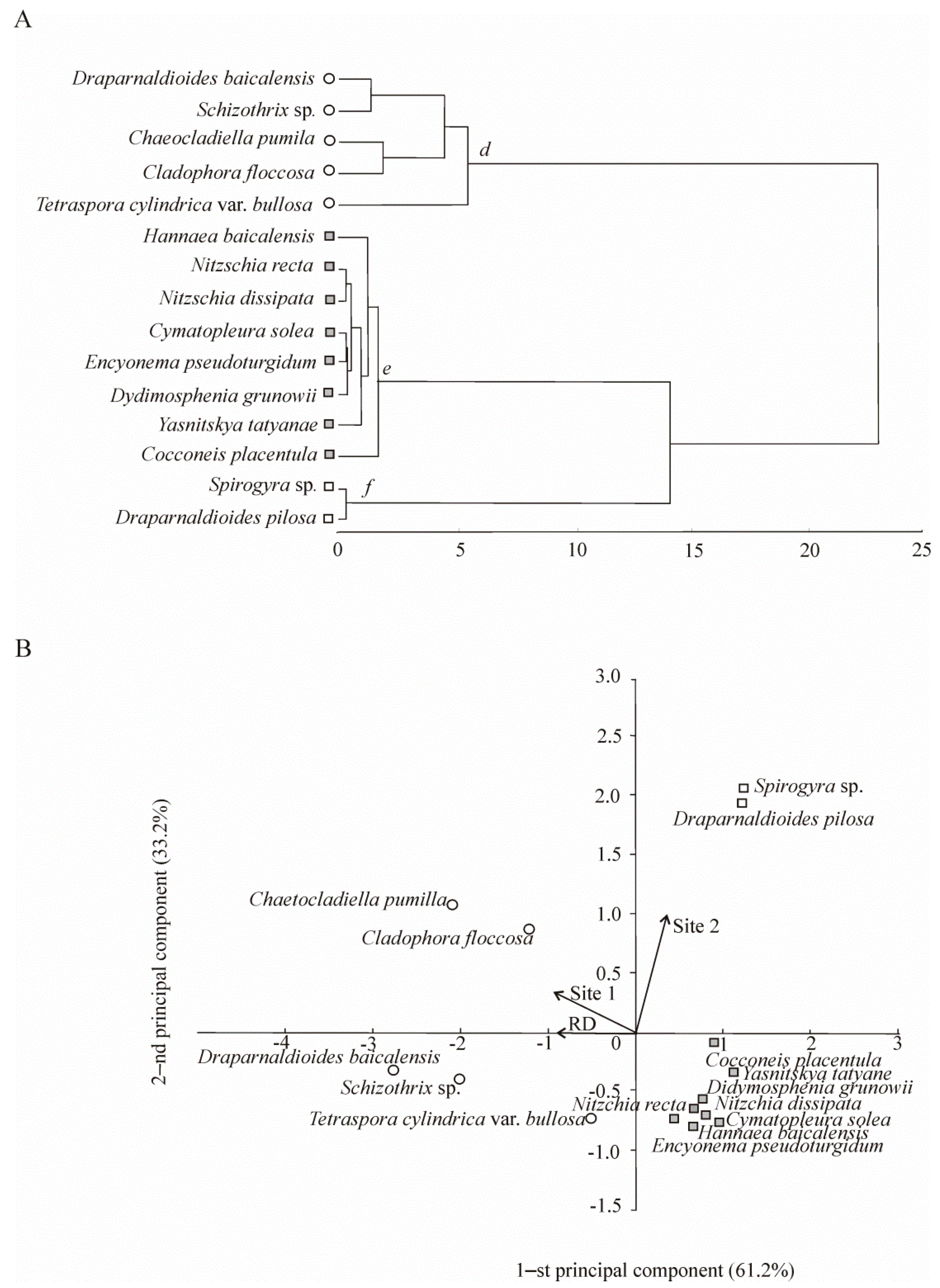
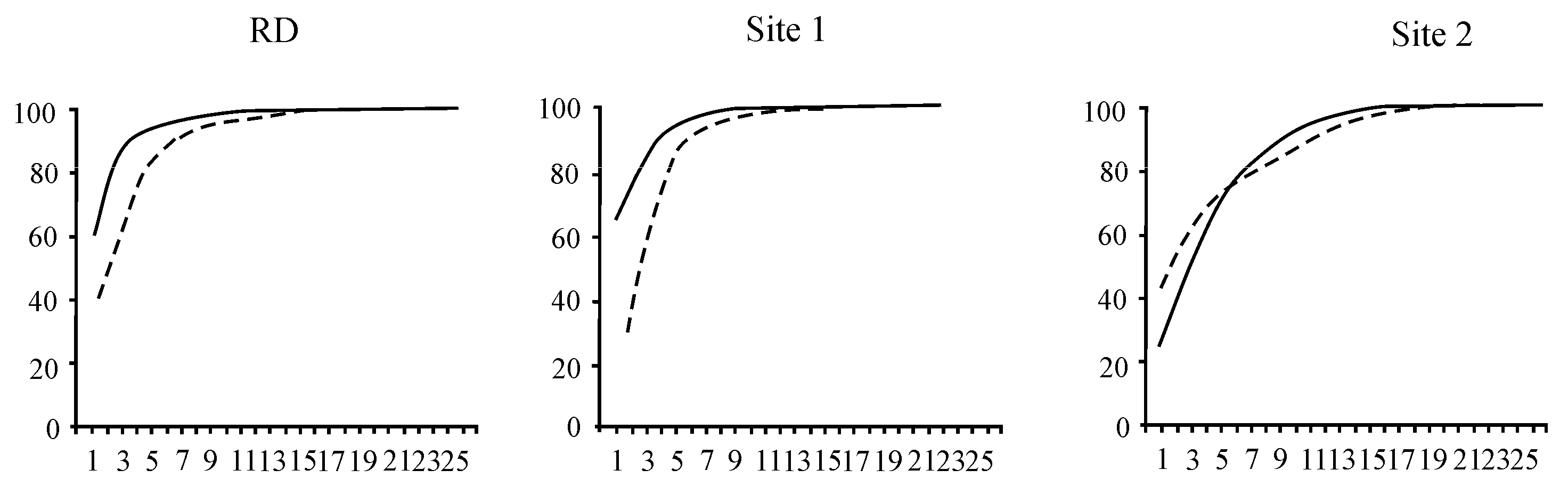
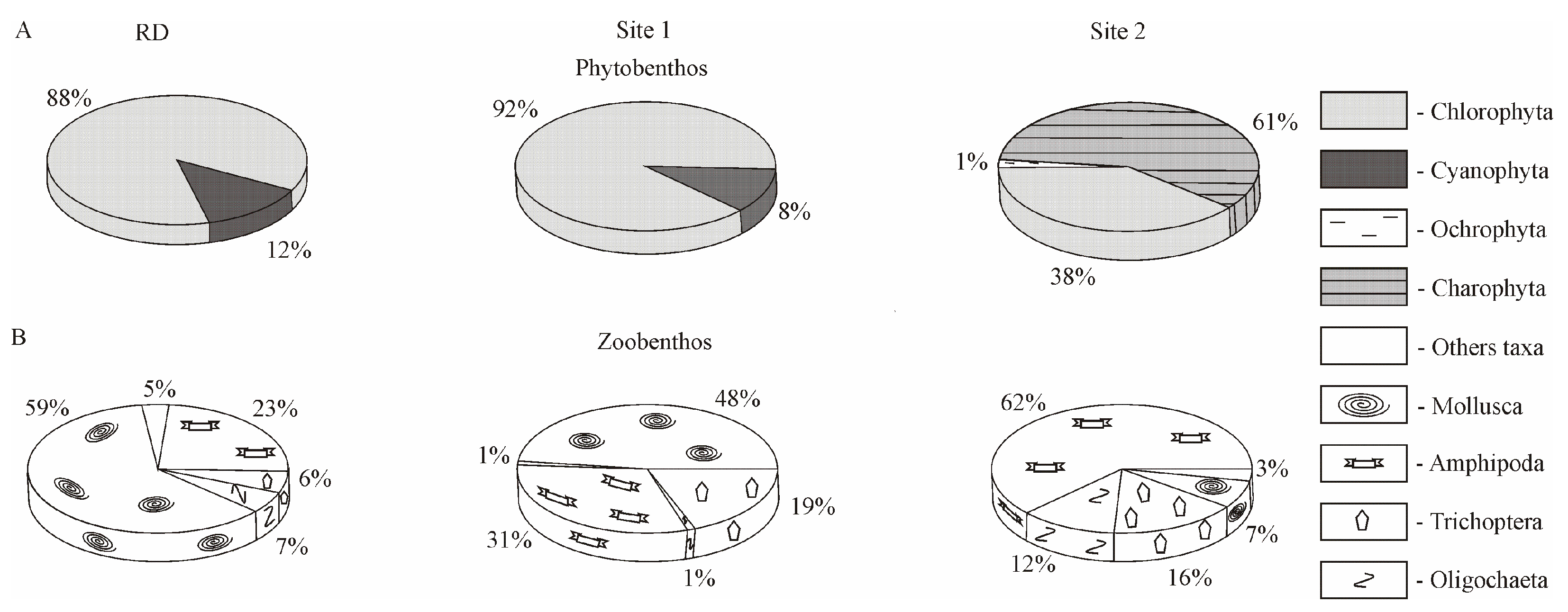
4.1. Biodiversity Pattern: The Species Richness and Structure of Aquatic Organisms Communities
4.2. Species Composition and Diversity of Aquatic Organisms in the Background Area (Site 1)
4.3. Species Composition and Diversity of Aquatic Organisms in the Area with Human Activity in Listvennichny Bay (Site 2)
5. Discussion
5.1. Response of Aquatic Organisms Communities to Climate Change
5.2. Current Structure of Aquatic Organisms Communities in the Area with Human Activity in Listvennichny Bay
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| TP | total phosphorus |
| TDP | total dissolved phosphorus |
| TC | thermotolerant coliforms |
| CFU | colony forming units |
| Tr. | transects |
| s | square |
| RD | reference data |
| PCA | principal component analysis |
| H | the Shannon diversity index |
| E/O | the ratio of eutrophic to oligotrophic species counts |
| M/E | the ratio of mesosaprobiont biomass to the biomass of Baikal endemics |
| S | the saprobity index of a water body or its part |
| Fij | the index of relative biotope confinement |
References
- Valipour, M.; Bateni, S.V.; Jun, C. Global Surface Temperature: A New Insight. Climate 2021, 9, 81. [Google Scholar] [CrossRef]
- Farooq, I.; Shah, A.R.; Salik, K.M.; Ismail, M. Annual, Seasonal and Monthly Trend Analysis of Temperature in Kazakhstan During 1970–2017 Using Non-parametric Statistical Methods and GIS Technologies. Earth Syst. Environ. 2021, 4, 1–21. [Google Scholar]
- Shimaraev, M.N.; Kuimova, L.N.; Sinyukovich, V.N.; Tsekhanovskii, V.V. Manifestation of global climate change in Lake Baikal during the 20th century. Dokl. Earth Sci. 2002, 383A, 288–291. [Google Scholar]
- Todd, M.C.; Mackay, A.W. Large-scale climatic controls on Lake Baikal ice cover. J. Clim. 2003, 16, 3186–3199. [Google Scholar] [CrossRef]
- Pilla, R.M.; Williamson, C.E.; Adamovich, B.V.; Adrian, R.; Anneville, O.; Chandra, S.; Colom-Montero, W.; Devlin, S.P.; Dix, M.A.; Dokulil, M.T.; et al. Deeper waters are changing less consistently than surface waters in a global analysis of 102 lakes. Sci. Rep. 2020, 10, 20514. [Google Scholar] [CrossRef]
- Izmest’eva, L.R.; Silow, E.A.; Litchman, E. Long-term dynamics of Lake Baikal pelagic phytoplankton under climate change. Inland Water Biol. 2011, 4, 301–307. [Google Scholar] [CrossRef]
- Hsieh, C.H.; Sakai, Y.; Ban, S.; Ishikawa, K.; Ishikawa, T.; Ichise, S.; Yamamura, N.; Kumagai, M. Eutrophication and warming effects on long-term variation of zooplankton in Lake Biwa. Biogeosciences 2011, 8, 1383–1399. [Google Scholar] [CrossRef] [Green Version]
- Cohen, A.S.; Gergurich, E.L.; Kraemer, B.M.; McGlue, M.M.; McIntyre, P.B.; Russell, J.M.; Simmons, J.D.; Swarzenski, P.W. Climate warming reduces fish production and benthic habitat in Lake Tanganyika, one of the most biodiverse freshwater ecosystems. Proc. Natl. Acad. Sci. USA 2016, 113, 9563–9568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hampton, S.E.; McGowan, S.; Ozersky, T.; Virdis, S.G.; Vu, T.T.; Spanbauer, T.L.; Kraemer, B.M.; Swann, G.; Mackay, A.W.; Powers, S.M.; et al. Recent ecological change in ancient lakes. Limnol. Oceanogr. 2018, 63, 2277–2304. [Google Scholar] [CrossRef] [Green Version]
- Albrecht, C.; Föller, K.; Clewing, C.; Hauffe, T.; Wilke, T. Invaders versus endemics: Alien gastropod species in ancient Lake Ohrid. Hydrobiologia 2014, 739, 163–174. [Google Scholar] [CrossRef]
- Lung’ayia, H.B.O.; M’harzi, A.; Tackx, M.; Gichuki, J.; Symoens, J.J. Phytoplankton community structure and environment in the Kenyan waters of Lake Victoria. Freshw. Biol. 2000, 43, 529–543. [Google Scholar] [CrossRef] [Green Version]
- O’Reilly, C.M.; Alin, S.R.; Plisnier, P.D.; Cohen, A.S.; McKee, B.A. Climate change decreases aquatic ecosystem productivity of Lake Tanganyika, Africa. Nature 2003, 424, 766–768. [Google Scholar] [CrossRef] [PubMed]
- Kozhov, M.M. Biology of Lake Baikal; USSR Academy of Sciences Press: Moscow, Russia, 1962; p. 315. [Google Scholar]
- Timoshkin, O.A.; Mazepova, G.F.; Melnik, N.G. Guid and Key to Pelagic Animals of Baikal (with Ecological Notes); Timoshkin, O.A., Mazepova, G.F., Melnik, N.G., Eds.; Science: Novosibirsk, Russia, 1995; p. 694. [Google Scholar]
- Potemkina, T.G.; Potemkin, V.L.; Kotsar, O.V.; Fedotov, A.P. Climate factors as a possible trigger of modern ecological changes in shallow zone of Lake Baikal (Russia). Int. J. Environ. Stud. 2018, 75, 86–98. [Google Scholar] [CrossRef]
- Shimaraev, M.N.; Domysheva, V.M. Trends in Hydrological and Hydrochemical Processes in Lake Baikal under Conditions of Modern Climate Change. In Climatic Change and Global Warming of Inland Waters: Impacts and Mitigation for Ecosystems and Societies; Goldman, C.R., Kumagai, M., Robarts, R.D., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2013; pp. 43–66. [Google Scholar]
- Kravtsova, L.S.; Izhboldina, L.A.; Khanaev, I.V.; Pomazkina, G.V.; Rodionova, E.V.; Domysheva, V.M.; Sakirko, M.V.; Tomberg, I.V.; Kostornova, T.Y.; Kravchenko, O.S.; et al. Nearshore benthic blooms of filamentous green algae in Lake Baikal. J. Great Lakes Res. 2014, 40, 441–448. [Google Scholar] [CrossRef]
- Timoshkin, O.A.; Moore, M.V.; Kulikova, N.N.; Tomberg, I.V.; Malnik, V.V.; Shimaraev, M.N.; Troitskaya, E.S.; Shirokaya, A.A.; Sinyukovich, V.N.; Zaitseva, E.P.; et al. Groundwater contamination by sewage causes benthic algal outbreaks in the littoral zone of Lake Baikal (East Siberia). J. Great Lakes Res. 2018, 44, 230–244. [Google Scholar] [CrossRef]
- Khanaev, I.V.; Kravtsova, L.S.; Maikova, O.O.; Bukshuk, N.A.; Sakirko, M.V.; Kulakova, N.V.; Butina, T.V.; Nebesnykh, I.A.; Belikov, S.I. Current state of the sponge fauna (Porifera: Lubomirskiidae) of Lake Baikal: Sponge disease and the problem of conservation of diversity. J. Great Lakes Res. 2018, 44, 77–85. [Google Scholar] [CrossRef]
- Bukin, Y.S.; Bondarenko, N.A.; Rusanov, I.I.; Pimenov, N.V.; Bukin, S.V.; Pogodaeva, T.V.; Chernitsyna, S.M.; Shubenkova, O.V.; Ivanov, V.G.; Zakharenko, A.S.; et al. Interconnection of bacterial and phytoplanctonic communities with hydrochemical parameters from ice and under-ice water in coastal zone of Lake Baikal. Sci. Rep. 2020, 10, 11087. [Google Scholar] [CrossRef]
- Khodzher, T.V.; Domysheva, V.M.; Sorokovikova, L.M.; Sakirko, M.V. Current chemical composition of Lake Baikal water. Inland Waters 2017, 7, 250–258. [Google Scholar] [CrossRef]
- Onischuk, N.A.; Netsvetaeva, O.G.; Tomberg, I.V.; Sakirko, M.V.; Domysheva, V.M.; Golobokova, L.P.; Khodzher, T.V. Seasonal dynamics of mineral forms of nitrogen in the rivers, snow cover and precipitation at the southwest coast of the Southern Baikal. Limnol. Freshw. Biol. 2019, 3, 245–252. [Google Scholar] [CrossRef]
- Malnik, V.V.; Timoshkin, O.A.; Suturin, A.N.; Onishchuk, N.A.; Sakirko, M.V.; Tomberg, I.V.; Gorshkova, A.S.; Zabanova, N.S. Anthropogenic Changes in the Hydrochemical and Sanitary–Microbiological Characteristics of Water Quality in Southern Baikal Tributaries: Listvennichnyi Bay. Water Resour. 2019, 46, 748–758. [Google Scholar] [CrossRef]
- Boeva, L.V. Guidelines on Chemical Analysis of Inland Surface Waters, Part 1; NOK: Rostov-on-Don, Russia, 2009; p. 1044. [Google Scholar]
- Wetzel, R.G. Limnological Analyses; Springer: New York, NY, USA, 1991; p. 391. [Google Scholar]
- Timoshkin, O.A.; Proviz, V.I.; Sitnikova, T.Y. (Eds.) Index of Animal Species Inhabiting Lake Baikal and Its Catchment Area; Vol. 2: Basins and Channels in the South of East Siberia and north Mongolia, Book 1; Science: Novosibirsk, Russia, 2009; p. 900. [Google Scholar]
- Tarasova, E.N.; Mescheryakova, A.I. Current State of the Hydrochemical Regime of Lake Baikal; Science: Novosibirsk, Russia, 1992; p. 143. [Google Scholar]
- Makarova, I.V.; Pichkily, L.O. Some problem in the procedure of calculating phytoplankton biomass. Botanic. Zhurn. 1970, 55, 1488–1494. [Google Scholar]
- Pesenko, Y.A. Principles and Methods of Quantitative Analysis in Faunistic Studies; Science: Moscow, Russia, 1982; p. 287. [Google Scholar]
- Odum, E.P. Fundamentals of Ecology, 3rd ed.; W.B. Saunders: Philadelphia, PA, USA, 1971; p. 226. [Google Scholar]
- Warwick, R.M.; Clarke, K.R. Relearning the ABC: Taxonomic changes and abundance/biomass relationships in disturbed benthic communities. Mar. Biol. 1994, 118, 739–744. [Google Scholar] [CrossRef]
- Pantle, R.; Buck, H. The biological monitoring of waters and the representation of results. Gas-u. Wasser. 1955, 96, 604. [Google Scholar]
- Sládecek, V. Zur Ermittlung des Indikations-Gewichtes in der biologischen Gewässeruntersuchung. Arch. Hydrobiol. 1964, 60, 241–243. [Google Scholar]
- Unified Methods for the Study of Water Quality; Part III. Methods of Biological Analysis of Waters. Indicators of Saprobidity; Secretariat of the CMEA: Moscow, Russia, 1977; p. 91.
- Barinova, S.S.; Medvedeva, L.A. Atlas of Algae-Indicators of Saprobicity (Russian Far East); Dalnauka: Vladivostok, Russia, 1996; p. 334. [Google Scholar]
- Hajnal, E.; Padisak, J. Analysis of Long-Term Ecological Status of Lake Balaton Based on the ALMOBAL Phytoplankton Database. Hydrobiologia 2008, 599, 227–237. [Google Scholar] [CrossRef]
- Kamenir, Y.; Winder, M.; Dubinsky, Z.; Zohary, T.; Schladow, G. Lake Tahoe vs. Lake Kinneret phytoplankton: Comparison of long-term taxonomic size structure consistency. Aquat. Sci. 2008, 70, 195–203. [Google Scholar] [CrossRef]
- Winder, M.; Reuter, J.E.; Schladow, S.G. Lake warming favours small-sized planktonic diatom species. Proc. R. Soc. B Biol. Sci. 2009, 276, 427–435. [Google Scholar] [CrossRef]
- Bondarenko, N.A.; Ozersky, T.; Obolkina, L.A.; Tikhonova, I.V.; Sorokovikova, E.G.; Sakirko, M.V.; Potapov, S.A.; Blinov, V.V.; Zhdanov, A.A.; Belykh, O.I. Recent changes in the spring microplankton of Lake Baikal. Limnologica 2019, 75, 19–29. [Google Scholar] [CrossRef]
- Izmest’eva, L.R.; Moore, M.V.; Hampton, S.E.; Silow, E.A. Seasonal dynamics of common phytoplankton in Lake Baikal. Proc. Samara Russ. Acad. Sci. Cent. 2006, 8, 191–196. [Google Scholar]
- Popovskaya, G.I.; Usoltseva, M.V.; Domysheva, V.M.; Sakirko, M.V.; Blinov, V.V.; Khodzher, T.V. The spring phytoplankton in the pelagic zone of Lake Baikal during 2007–2011. Geogr. Nat. Resour. 2015, 36, 253–262. [Google Scholar] [CrossRef]
- Votintsev, K.K.; Popovskaya, G.I.; Mazepova, G.F. Physical and Chemical Regime and Life of Plankton in the Selenga Region of Lake Baikal; Academy of Sciences of the USSR Press: Moscow, Russia, 1963; p. 322. [Google Scholar]
- Ryves, D.B.; Jewson, D.H.; Sturm, M.; Battarbee, R.W.; Flower, R.J.; Mackay, A.W.; Granin, N.G. Quantitative and qualitative relationships between planktonic diatom communities and diatom assemblages in sedimenting material and surface sediments in Lake Baikal, Siberia. Limnol. Oceanogr. 2003, 48, 1643–1661. [Google Scholar] [CrossRef] [Green Version]
- Coumou, D.; Lehmann, J.; Beckmann, J. The weakening summer circulation in the Northern Hemisphere mid-latitudes. Science 2015, 348, 324–327. [Google Scholar] [CrossRef] [PubMed]
- Panizzo, V.N.; Roberts, S.; Swann, G.E.A.; McGowan, S.; Mackay, A.W.; Vologina, E.; Pashley, V.; Horstwood, M.S.A. Spatial differences in dissolved silicon utilization in Lake Baikal, Siberia: Examining the impact of high diatom biomass events and eutrophication. Limnol. Oceanogr. 2018, 63, 1562–1578. [Google Scholar] [CrossRef] [Green Version]
- Swann, G.E.; Panizzo, V.N.; Piccolroaz, S.; Pashley, V.; Horstwood, M.S.; Roberts, S.; Vologina, E.; Piotrowska, N.; Sturm, M.; Zhdanov, A.; et al. Changing nutrient cycling in Lake Baikal, the world’s oldest lake. Proc. Natl. Acad. Sci. USA 2020, 117, 27211–27217. [Google Scholar] [CrossRef]
- O’Donnell, D.R.; Wilburn, P.; Silow, E.A.; Yampolsky, L.Y.; Litchman, E. Nitrogen and phosphorus colimitation of phytoplankton in Lake Baikal: Insights from a spatial survey and nutrient enrichment experiments. Limnol. Oceanogr. 2017, 62, 1383–1392. [Google Scholar] [CrossRef]
- Vollenweider, R.A. Concept of nutrient load as a basis for the external control of the eutrophication process in lakes and reservoirs. Zeitsch. Wass. Adwass. Forsch. 1979, 12, 46–56. [Google Scholar]
- Izhboldina, L.A. Meio- and Macrophytobenthos of Lake Baikal (Algae); Irk. State University Press: Irkutsk, Russia, 1990; p. 176. [Google Scholar]
- Pomazkina, G.V.; Shcherbakova, T.A. Characteristics of mass species of Bacillariophyta of littoral zone of Lake Baikal (Russia). Algology 2011, 21, 52–67. [Google Scholar]
- Kravtsova, L.S.; Mizandrontsev, I.B.; Vorobyova, S.S.; Izhboldina, L.A.; Mincheva, E.V.; Potyomkina, T.G.; Golobokova, L.P.; Sakirko, M.V.; Triboy, T.I.; Khanaev, I.V.; et al. Influence of water motion on the spatial distribution of Spirogyra in Lake Baikal. J. Great Lakes Res. 2020, 46, 29–40. [Google Scholar] [CrossRef]
- Pourriot, R.; Clement, P. Action de facteurs externes sur la reproduction et le cycle reproducteur des rotifères. Acta Oecol. Gen. 1981, 2, 135–151. [Google Scholar]
- Hampton, S.E.; Izmest’eva, L.R.; Moore, M.V.; Katz, S.L.; Dennis, B.; Silow, E.A. Sixty years of environmental change in the world’s largest freshwater lake—Lake Baikal, Siberia. Glob. Chang. Biol. 2008, 14, 1947–1958. [Google Scholar] [CrossRef] [Green Version]
- Kappes, H.; Mechenich, C.; Sinsch, U. Long-term dynamics of Asplanchna priodonta in LakeWindsborn with comments on the diet. Hydrobiologia 2000, 432, 91–100. [Google Scholar] [CrossRef]
- Genkai-Kato, M.; Sekino, T.; Yoshida, T.; Miyasaka, H.; Khodzher, T.V.; Belykh, O.A.; Melnik, N.G.; Kawabata, Z.; Higashi, M.; Nakanishi, M. Nutritional diagnosis of phytoplankton in Lake Baikal. Ecol. Res. 2002, 17, 135–142. [Google Scholar] [CrossRef]
- Guildford, S.J.; Hecky, R.E. Total nitrogen, total phosphorus, and nutrient limitation in lakes and oceans: Is there a common relationship? Limnol. Oceanogr. 2000, 45, 1213–1223. [Google Scholar] [CrossRef] [Green Version]
- Tezuka, Y. C:N:P ratios of seston in Lake Biwa as indicators of nutrient deficiency in phytoplankton and decomposition process of hypolimnetic particulate matter. Jpn. J. Limnol. 1985, 46, 239–246. [Google Scholar] [CrossRef]
- Brown, K.R.; Gerber, A.; Bedulina, D.; Timofeev, M.A. Human impact and ecosystemic helth at Lake Baikal. Water 2021, 8, 1528. [Google Scholar]
- Pokrovskaya, T.N.; Mironova, N.Y.; Shilkrot, G.S. Macrophyte Lakes and Their Eutrophication; Science: Moscow, Russia, 1983; p. 153. [Google Scholar]
- Nozaki, K.; Darijav, K.; Akatsuka, T.; Goto, N.; Mitamura, O. Development of green algae in the benthic algal community in a littoral sand-beach zone of Lake Biwa. Limnology 2003, 4, 161–165. [Google Scholar] [CrossRef]
- Rosenberger, E.E.; Hampton, S.E.; Fradkin, S.C.; Kennedy, B.P. Effects of shoreline development on the nearshore environment in large deep oligotrophic lakes. Freshw. Biol. 2008, 53, 1673–1691. [Google Scholar] [CrossRef]
- Hainz, R.; Wöber, C.; Schagerl, M. The relationship between Spirogyra (Zygnematophyceae, Streptophyta) filament type groups and environmental conditions in Central Europe. Aquat. Bot. 2009, 91, 173–180. [Google Scholar] [CrossRef]
- Kravtsova, L.S.; Mekhanikova, I.V.; Izhboldina, L.A. Role of Algae Cenoses in the Spatial Distribution of Macrozoobenthos in the Littoral Zone of Lake Baikal. Hydrobiol. J. 2008, 44, 16–25. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Area | Water | Si | NO2− | NH4+ | NO3− | P-PO43− | TDP | TP |
|---|---|---|---|---|---|---|---|---|
| Site 1 | surface water of Baikal coastal zone | 0.23 ± 0.03 | 0.003 ± 0.002 | 0.009 ± 0.000 | 0.10 ± 0.00 | 0.004 ± 0.001 | 0.006 ± 0.001 | 0.018 ± 0.009 |
| near-bottom water of Baikal coastal zone | 0.18 ± 0.03 | 0.002 ± 0.001 | 0.009 ± 0.003 | 0.14 ± 0.02 | 0.004 ± 0.000 | 0.006 ± 0.002 | 0.010 ± 0.001 | |
| groundwater of Baikal beach | 2.71 ± 2.50 | 0.006 ± 0.004 | 0.015 ± 0.005 | 0.23 ± 0.04 | 0.005 ± 0.001 | 0.013 ± 0.007 | 0.025 ± 0.014 | |
| Site 2 | surface water of Baikal coastal zone | 0.19 ± 0.01 | 0.002 ± 0.001 | 0.008 ± 0.001 | 0.11 ± 0.00 | 0.003 ± 0.001 | 0.005 ± 0.000 | 0.010 ± 0.001 |
| near-bottom water of Baikal coastal zone | 0.17 ± 0.02 | <0.002 | 0.007 ± 0.001 | 0.14 ± 0.02 | 0.003 ± 0.001 | 0.005 ± 0.000 | 0.011 ± 0.001 | |
| groundwater of Baikal beach | 1.37 ± 0.64 | 0.034 ± 0.026 | 0.009 ± 0.001 | 0.23 ± 0.04 | 0.030 ± 0.006 | 0.035 ± 0.005 | 0.063 ± 0.011 | |
| Rivers | mouth of the Krestovka and Cheremshanka rivers | 7.07 ± 0.22 | 0.038 ± 0.033 | 0.009 ± 0.003 | 3.94 ± 3.09 | 0.027 ± 0.018 | 0.029 ± 0.018 | 0.034 ± 0.019 |
| RD | surface water of Baikal coastal zone near Berezovy Cape in 2000 according to [26] | 0.46 | 0.002 | 0.010 | 0.08 | 0.002 | ||
| near-bottom water of Baikal coastal zone near Berezovy Cape in 2000 according to [26] | 0.47 | 0.007 | 0.035 | 0.23 | 0.004 | |||
| RD | surface water of Baikal southern basin in the 1987 according to [27] | 1.14 | 0.23 | 0.004 |
| Taxa | Fij | ||
|---|---|---|---|
| RD | Site 1 | Site 2 | |
| Phytoplankton | |||
| Anabaena lemmermannii P. Richt. | −0.316 | +0.651 | −0.589 |
| Asterionella formosa Hass. | −0.996 | +0.284 | +0.331 |
| Ceratium hirundinella (O. Müll.) Schrank. | −0.633 | +0.267 | +0.182 |
| Dinobryon cylindricum Imhof. | −0.999 | +0.179 | +0.427 |
| Dinobryon divergens Imhof. | −0.988 | +0.185 | +0.419 |
| Dinobryon sociale Ehr. | −0.985 | −0.236 | +0.701 |
| Gymnodinium coeruleum Ant. | +0.120 | −0.135 | 0.000 |
| Rhodomonas pusilla (Bachm.) Javorn. | +0.529 | −0.222 | −0.451 |
| Spirogyra sp. | −1.000 | +0.010 | +0.557 |
| Fragilaria radians Kütz. | −0.988 | +0.202 | +0.403 |
| picoplankton | +0.735 | −0.539 | −0.591 |
| Zooplankton | |||
| Asplanchna priodonta Gosse. | −0.809 | +0.551 | +0.441 |
| Bosmina longirostris (O.F.Müller). | −0.693 | +0.417 | +0.497 |
| Cyclops kolensis Lilljeborg. | +0.880 | −0.909 | −0.403 |
| Daphnia galeata Sars. | +0.184 | −0.324 | +0.368 |
| Epischura baikalensis Sars. | +0.047 | +0.057 | −0.644 |
| Leptodora kindtii (Focke). | −1.000 | +0.714 | +0.423 |
| Synchaeta grandis Zacharias. | −0.928 | +0.370 | +0.712 |
| Benthic macroalgae | |||
| Spirogyra sp. | −1.000 | −1.000 | +0.999 |
| Chaetocladiella pumila (Meyer) C.Meyer et Skabitsch. | +0.401 | +0.306 | −0.974 |
| Cladophora floccosa C.Meyer. | +0.880 | −0.779 | −0.850 |
| Draparnaldioides baicalensis C.Meyer et Skabitsch. | +0.301 | +0.418 | −1.000 |
| Draparnaldioides pilosa C.Meyer et Skbitsch. | −1.000 | −1.000 | +1.000 |
| Tetraspora cylindrica var. bullosa C. Meyer. | +1.000 | −1.000 | −1.000 |
| Schizothrix sp. | +0.601 | +0.060 | −1.000 |
| Benthic microalgae | |||
| Cocconeis placentula Ehrenb. | −0.853 | +0.453 | +0.649 |
| Cymatopleura solea (Bréb) W.Sm. | +0.206 | +0.343 | −1.000 |
| Didymosphenia grunowii Lange-Betalot & Metzel. | −0.131 | −0.311 | +0.320 |
| Encyonema pseudoturgidum Pomazk. & Rodion. | +1.000 | −1.000 | −1.000 |
| Hannaea baicalensis Genk., Popovsk. and Kulik. | +0.987 | −0.984 | −0.983 |
| Nitzschia dissipata (Kütz.) Grun. | +0.491 | −0.366 | −0.480 |
| Nitzschia recta Hantzsch. | +1.000 | −1.000 | −1.000 |
| Yasnitskya tatyanae Pomazk. & Rodion. | −1.000 | −1.000 | +1.000 |
| Value | Phytoplankton | Zooplankton | Macroalgae (Meyo-, Macrophytes) | Microalgae | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| RD | Site 1 | Site 2 | RD | Site 1 | Site 2 | RD | Site 1 | Site 2 | RD | Site 1 | Site 2 | |
| Number of taxa | 42 | 45 | 46 | 26 | 19 | 24 | 15 | 23 | 28 | 35 | 33 | 33 |
| β-diversity by Shannon, H | 1.5 | 2.8 | 2.7 | 1.2 | 1.4 | 2.0 | 1.8 | 1.0 | 2.1 | 2.0 | 2.1 | 1.7 |
| Number of indicator species | 36 | 37 | 39 | 18 | 13 | 17 | 7 | 10 | 11 | 27 | 20 | 18 |
| Saprobity index, S | 1.47 | 1.60 | 1.70 | 1.45 | 1.39 | 1.53 | 1.48 | 1.57 | 2.17 | 1.0 | 1.28 | 1.45 |
| Ratio of eutrophic species number to oligotrophic species number, E/O | 2:1 | 2:1 | 2:1 | 1:2 | 1:2 | 1:2 | 1:1 | 1:2 | 3:1 | 1:3 | 1:2 | 1:2 |
| Ratio of mesosaprobiont species biomass to endemic species biomass, M/E | 1:6 * | 3:1 * | 6:1 * | 1:4 | 1:5 | 2:1 | 1:18 | 1:7 | 3:1 | 1:47 | 1:2 | 7:1 |
| Total biomass, B ± m | 190.7 ± 41.5 | 186.9 ± 22.5 | 246.0 ± 16.3 | 672.7 ± 158.6 | 723.7 ± 166.1 | 100.7 ± 14.3 | 245,500 ± 65,500 | 185,287 ± 74,870 | 367,564 ± 119,414 | 39.3 ± 10.5 | 10.0 ± 3.2 | 12.7 ± 5.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kravtsova, L.; Vorobyeva, S.; Naumova, E.; Izhboldina, L.; Mincheva, E.; Potemkina, T.; Pomazkina, G.; Rodionova, E.; Onishchuk, N.; Sakirko, M.; et al. Response of Aquatic Organisms Communities to Global Climate Changes and Anthropogenic Impact: Evidence from Listvennichny Bay of Lake Baikal. Biology 2021, 10, 904. https://doi.org/10.3390/biology10090904
Kravtsova L, Vorobyeva S, Naumova E, Izhboldina L, Mincheva E, Potemkina T, Pomazkina G, Rodionova E, Onishchuk N, Sakirko M, et al. Response of Aquatic Organisms Communities to Global Climate Changes and Anthropogenic Impact: Evidence from Listvennichny Bay of Lake Baikal. Biology. 2021; 10(9):904. https://doi.org/10.3390/biology10090904
Chicago/Turabian StyleKravtsova, Lyubov, Svetlana Vorobyeva, Elena Naumova, Lyudmila Izhboldina, Elena Mincheva, Tatyana Potemkina, Galina Pomazkina, Elena Rodionova, Natalya Onishchuk, Mariya Sakirko, and et al. 2021. "Response of Aquatic Organisms Communities to Global Climate Changes and Anthropogenic Impact: Evidence from Listvennichny Bay of Lake Baikal" Biology 10, no. 9: 904. https://doi.org/10.3390/biology10090904
APA StyleKravtsova, L., Vorobyeva, S., Naumova, E., Izhboldina, L., Mincheva, E., Potemkina, T., Pomazkina, G., Rodionova, E., Onishchuk, N., Sakirko, M., Nebesnykh, I., & Khanaev, I. (2021). Response of Aquatic Organisms Communities to Global Climate Changes and Anthropogenic Impact: Evidence from Listvennichny Bay of Lake Baikal. Biology, 10(9), 904. https://doi.org/10.3390/biology10090904